Ce este de obicei clasificat Alzheimer? Alzheimer nu este o boala specifica, ci mai degrabă un set de simptome. Exista multe „cauze radacina/de baza” diferite ale setului de simptome cunoscut sub numele de „Alzheimer.”
Unele dintre elementele care sunt cunoscute de a provoca Alzheimer includ:
1) cantități masive de microbi din organism.
Microbii elimina toxine și aceste toxine (produse reziduale ale microbilor) sunt extrem de acide și interfereaza cu semnalizarea (interneuronala) de pe creier, provocând „ceață pe creier / brain fog”
2) Metale grele (de exemplu, mercur si altele),
3) Alte metale (de exemplu aluminiu).
4) atacuri cerebrale ( stiute sau nestiute),
5) radicalii liberi ( stres oxidativ),
6) Boala Lyme (frecvent diagnostica gresit ca scleroza in placi/multipla )
7) hipertensiunea
8) Microbii și paraziți din organe, care slăbesc sistemul imunitar,
9) droguri ilegale,
10) Unele medicamente prescrise ( farma),
11) Lipsa de oxigen. Odata ce imbatranim, mai puțin oxigen este absorbit, deși, în multe cazuri, problema nu este o lipsa de oxigen, ci toxinele secretate de microbi (vezi # 1),
12) Dieta, care este bogata in grasimi rele (de exemplu, acizi grași trans) si saraca in grasimi bune (de exemplu, omega-3).Aceasta include ingerarea unor tipuri de grăsimi (de exemplu, grăsimi saturate, hidrogenate sau partial hidrogenate) frecvent (consumul frecvent de grasimi animale, margarina vegetala si uleiuri vegetale hidrogenate(chiar si partial), unt de arahide,etc)
13) Aspartam, glutamatul monosodic, HVP, cisteină și alte „excitotoxine” (care distrug celulele creierului),
14) Lipsa de magneziu și alte minerale / vitamine / enzime/ nutrienti esentiale(i),
Din păcate, medicina alopata, in incercarea lor continua să vândă droguri și de a obține averi, foarte rar face orice efort pentru a găsi „cauza rădăcină” a bolii Alzheimer. Medicina alopata ”trateaza” tipic doar simptomele vizuale ale bolii, și, rareori, „sapă mai adânc de atât”.
Metale grele, cum ar fi mercurul (surse sunt amalgamul dentar, consumul de cantități mari de pește oceanic, vaccinări, etc), plumbul (găsit în ciocolată, printre altele) și alte metale grele care pot veni de la apa de izvor/nefiltrata, de exemplu, poate fi o cauza de boli ale creierului precum Alzheimer, dementa si poate si Parkinson .
Aluminiu (care poate fi introdus si de vaccinări sau din alte surse precum vase gatit/folie aluminu ), nu este departe de metale grele.
Din toate aceste lucruri important este că nu este suficient oxigen si nici suficiente grasimi bune ce intra in creier ci prea multe lucruri nocive ajung in creier.
Sau altceva interferează cu semnalizarea (interneuronala) a creierului.
Atunci când creierul beneficiaza de o multime de oxigen, și nu blochează nimic semnalizarea creierului, mintea este limpede și ascuțita.
Este atunci când lucruri cum ar fi enumerate mai sus, sau droguri ilegale, . blocheaza oricare dintre aceste două lucruri apar probleme majore.
Optiuni/ Suplimentele care ar putea fi /ar trebui luate zilnic
Aceasta este o listă de suplimente zilnice pentru atunci când memoria este o problemă.
„Secret” este un anti-oxidant superb care activeaza enzimele SOD in interiorul celulelor. Nu pot menționa numele acestuia, astfel că va trebui să meargeti la un alt site, care este protejată prin parolă, pentru a afla mai multe despre aceasta: Utilizator = no Parola = link http://www.natural-product.info/Product01/Product01.html
(exact 2 pilule pe zi conform cancertutor.com -uitati-va la ce contine si inlocuiti cu ingrediente naturale din alte surse in dozele adecvate daca nu se paote obtine acest produs ) 9) Suplimente de iod (de exemplu potasiu iod, alge marin Kelp-atentie sa fie sursa bio, farmaemtale grele ) 10) Supliment de OMEGA 3 ( nucifere ,samburi si seminte, ulei de in , ulei de ficat COD si de KRILL (dovedite în studii – atentie sa nu contina metale grele aceste suplimente oceanice) ) 11) Argintul Coloidal sau alte lucruri care ucid microbii din sânge 12) supliment de magneziu (de exemplu, supliment calciu / magneziu / zinc) 13) picolinat de crom 14) ulei de cocos (de calitate sau nuca de cocos intreaga-ieftina si aduce multe alte beneficii) și suplimente de zinc 15) Vitamina B1 (tiamina) 16) Vitamina B2 (riboflavina) 17) Vitamina B3 (niacina / niacinamide) 18) Vitamina B12 (vitamina cheie) 19) Bacopa (planta de probleme de memorie) 21) Ginkgo Biloba (planta de probleme de memorie) 22) transfer point (pentru sistemul imunitar sau alt supliment imunitar precum ciuperci imuno,terapii ozon/apa ionizata alcalina(bune si pentru oxigenare), aparate electromedicale precum Bob Beck, regulatori imunitari gen 714X, etc –
Un alt element cheie este chelarea de mercur si aluminiu din creier !!
In multe cazuri, acestea sunt o cauză majoră de dementa si Alzheimer:
Aici este o listă a acestor produse:
1) Zeoliti 2) Chlorella (atât un chelator și furnizează minerale esențiale) 3) Cilantro (chelant de mercur și aluminiu) 4) litiu orotat (a se vedea, de asemenea, # 27, în Manualul de referință) 5) EDTA Chelation (tratament la clinica, probabil cea mai bună metodă) 6) Găsiți un medic specializat în chelare
Mai multe comentarii
Acest articol vorbește despre multe alte lucruri relevante.
Cauza numarul 1 boli creier – toxinele(secrette de microorgansime nocive, toxine care blocheaza/interfereaza cu semnalizarea interneuronala radio a creierului ) Probabil, niciodată nu a trecut prin mintea cititorului ca niste microbi ar putea fi cauza majora de Alzheimer.
Sunt !!Pentru ca acesti microbi secreta toxine si aceste toxine sunt foarte acide și blocheaza semnalizarea radio din creier.
Cum pot bloca toxinele semnalizarea creier?
Ceea ce oamenii nu realizeaza este faptul că o mare parte din semnalizarea creierului este similar cu un turn de unde radio. Creierul nu are semnale exclusiv prin reacții chimice. S-a demonstrat că creierul comunica și prin intermediul undelor electrice.
Nu microbii înșiși sunt care blocheaza aceste unde electrice, deși acestia ar putea. Problema sunt „toxinele”, sau „deseurile” sau „excrementele” secretate de acesti microbi care produc daune. Acestea se amestecă cu sângele și sunt foarte eficiente in blocarea semnalizarii creierului.
Toxinele sunt foarte acide. Acești acizi pot dizolva literalmente nervii !! În timp ce corpul poate, în condiții adecvate, reconstrui aceste nervi, regenerarea nu va fi capabila de a restabili memoria pierdută, dar va restabili creierul.
Pneumoniae Chlamydia și Helicobacter pylori sunt 2 microbi care pot provoca Alzheimer. Herpes simplex virus 1 este asociat cu boala Alzheimer la persoanele care posedă forma APOE4 a genei APOE (APOE4 permite virusului herpes să intre creier).
Protozoarul Toxoplasma gondii a fost asociată cu boala Alzheimer.
Dar amintiți-vă că sunt toxinele/deseurile produse de ele care blochează semnalizarea creierlui mai mult decat microbi în sine.
Mai mult , uciderea in acelasi timp a unui număr masiv de microbi genereaza o ceata pe creier si mai masiva. Când acesti microbi mor inacelasi timp in numar mare creierul este inundat toxine.
Unde trăiesc aceste microbi?
Ei trăiesc în principal în ficat. Ficatul este un locde inmultire pentru microbi. Produsele reziduale sunt răspândite prin sange.
Unii dintre paraziți / flukes/tenii din ficat pot fi de 20 de picioare lungime !!
Și exista microbi în creier în sine, deoarece există microbi in sangele fiecarei persoane.
Ei excreta cantitati masive de toxine foarte acide. Creierul pacientilor/bolnavilor de Alzheimer innoata literalmente într-o mare de toxine. Cu toate toxinele este un miracol ca, creierul poate funcționa și, în multe cazuri, nu poate.
Deci, cum puteti scapa de acesti microbi si toxine lor? Sunt menționate(de catre cancertutor.com) două moduri majore.
Primul mod este Protocolul Photon.
Acesta este modul „tintit”, care se concentrează pe microbii specifici care provoaca boala.
A doua modalitate este Protocolul Bob Beck.
Aceasta curăță sângele de microbi si ajuta astfel si sistemul imunitar
Exista si alte modalitati enuntate in finalul articolului.Consider ca fiecare trebuie sa faca ce ii este la indemana, functie de posibilitati, sa se orienteze si sa se descurce cum poate ,nu sa se simta complexat de limitari materiale (pe care cu totii le avem).In plus, spre deosebire de limitarile materiale, accesul la Dumnezeu este gratuit si ajutorul DOMNULUI intrece orice lucru material:
„cautati mai intai imparatia lui Dumnezeu si toate celelalte se vor adauga voua.”
Personal, mi -as folosi de toate metodele accesbile mie si as observa ce functioneaza in cazul meu.
1) Protocolul Photon
O modalitate (recomandata de cei de la cancertutor ) de scapa de microbi este de a lucra cu Institutul Ed Skilling. Folosind o consultare pot identifica microbul specific care cauzează Alzheimer, sau orice altceva este cauza Alzheimer.
Dacă problemele sunt toxine, ei vor recomanda probabil un protocol Photon (care este utilizat în mod normal pentru cancer), însemnând unul sau ambele dispozitive Photon.
Există două dispozitive Ed Skilling, Photon Genie (aproximativ 3000 $) și Photon Genius(aproximativ 20000 $).(sume uriase pentru cei mai multi din noi; cu toate acestea, am vazut un Genie fol0sit de mai multi bolnavi cu diverse afectiuni grave si toti au fost multumiti;insa consider ca fiecare dintre noi are liber arbitru si un punct de plecare inainte de a face astfel de investii este de a vorbi cu fosti utilizatori )
Ambele aparate, precum si Quantum Pulse au la baza o tehnologie Nicolas Tesla.
Photon Genie poate fi plasat sub saltea pe timp de noapte sau se legat de corp, chiar în timp ce conduceți. Poate fi folosit 24 de ore pe zi, 7 zile pe săptămână (de exemplu, 24/7) pentru a energiza celulele , a intari imunitatea și pt a face alte lucruri.
Photon Genius este de sase picioare inaltime si 5 picioare lățime. Acesta este în esență o versiune „supraalimentata” Photon Genie (Institutul Skilling il numește „Genie pe steroizi”), dar de fapt include si alte lucruri, cum ar fi infra-roșu.
Datorită puterii și dimensiunilor sale Photon Genius este staționar și are nevoie de un circuit dedicat de 40-amp.
Deoarece Photon Genie este atât un dispozitiv 24/7 și poate ajuta la regenerarea nervilor, chiar dacă o persoană opteaza pentru un Genius(cel mai scump), acestea ar trebui să cumpere, de asemenea, un Genie.
Pentru claritate, un Genie(cel mai ieftin) este necesar în fiecare caz și un Genius este opțională pentru cei care își pot permite (presupunând consultarea identifică toxine sunt problema).
Dar, din nou, mai întâi o consultare cu Institutul Ed Skilling pentru a afla „cauza rădăcină” a Alzheimer.
Consultarea va analiza exact care specii de microbi cauzează Alzheimer și / sau a oricărui alt factor care contribuie.Tratamentul poate direcționa apoi functie cauzele specifice ale Alzheimer.
NOTA: si aceasta consultare este destul de scumpa(desi e foarte posibil sa fie inclusa in pretul unui astfel de aparat ).Daca nu as fi vazut pe viu lucrand astfel de aparate (GENIE) pe bolnavi de cancer si faptul ca intr-adevar au efecte benefice, nu stiu daca as fi mentionat despre ele(si aceasta pentru ca au pretul pe care il au si cand vine vorba de bani omul se transforma…)
Consultarea va duce la un protocol nutritional care este sinergică cu dispozitivul Photon. Protocolul nutrițional va include o curețare ficat pentru a scapa de trematode hepatice și alți paraziți de ficat.
Este o abordare orientată, dar o consultare este disponibila pentru fiecare pacient
Din ce mi-au spus alti pacienti cancer consultarea in sine este 1500 dolari (ceea ce pentru bugetul nostru este destul de/foarte scump).Intrebati totusi daca nu cumva este inclusa
Pentru mai multe informații, faceți clic pe aceste link-uri:
„Cercetatorii au analizat 1658 adulti care au fost de 65 de ani sau mai în vârstă. Toti acestia puteau merge fără ajutor și nu aveau semne de dementa, boli cardiovasculare, accident vascular cerebral când a început studiul. Cercetatorii au urmat evolutia acestor participanți timp de șase ani. Au vrut sa afle care dintre participanti vor continua sa dezvolte boala Alzheimer și alte forme de demență. Iată ce au descoperit: Cei care au avut deficit moderat de vitamina D au avut un risc cu 53% mai mare de a dezvolta dementa de orice fel. Dar la cei care au fost grav deficit s-a văzut cum riscul lor explodeaza la 125%.
Iar rezultatele au fost similare pentru boala Alzheimer. Cei din grupul moderat deficitar au avut 69% mai multe sanse de a dezvolta boala. Iar la cei cu un deficit sever s-a văzut riscul crescand cu 122%.
Dr. David Llewellyn, cercetator, a declarat: „Ne-am așteptat să găsim o asociere intre nivelurile de vitamina D scăzute și riscul de dementa si a bolii Alzheimer. Dar rezultatele au fost surprinzătoare. Noi de fapt, am constatat că asociația a fost de două ori la fel de puternica fata de ce am anticipat! „
Aceste studii oferă pondere semnificativă a puterii de vitamina D3(adica a luminii solare) pentru a proteja creierul . Aceste studii spun că va ajuta la prevenirea demenței și Alzheimer – și că ar putea fi un tratament puternic, de asemenea.
Nu ar trebui sa surpinda totusi pe nimeni , avand in vedere si rolul antimicrobian pe care il are vitamina D, care o face utila si in cancer si in alte boli microbiene.
Desi puteti începe să luați vitamina D3 astăzi( 5000 UI pe zi vor ajuta /muta nivelurile sanguine de 70+ optim ng / ml. ) expunerea pielii la soare aduce avantaje NET superioare.
3) Protocolul Bob Beck cu deparazitare hepatica O curatire si energizare a ficatului este parte a protocolului Photon, astfel încât curatarea ficatului în această secțiune este necesară doar în cazul în care o persoană nu utilizează atât Protocolul Photon cat si Protocolul Bob Beck impreuna.
Protocolul Bob Beck este mai ieftin și, de fapt unele elemente ale protocolului Bob Beck pot fi aplicate efectiv la domiciliu.Protocolul Bob Beck poate curăța sângele de microbi, si ar trebui sa ajute pacientii cu Alzheimer foarte mult di nacest punct de vedere(la fel si alte tehnici de curatare a sangelui de microbi , precum terapiile cu ozon(inclusiv apa ozonata si apa ionizata alcalina) , vitamina C IntraVenos, etc) dar nu poate scapa de unele dintre cei mai importanti paraziti din ficat și alți paraziți care nu sunt circula in sange, fara deparazitarea ficatului.
Combinat cu Protocolul Bob Beck ar trebui să fie o curatare de ficat si o curatare a vezicii biliare.
Curatarea vezicii biliare trebuie să protejeze vezica biliare de ceea ce se elimina din ficat.
Aceasta este Protocolul Bob Beck (de fapt mai multe articole): Bob Beck-link
Aici este un articol de pe valuri hepatice: undele hepatice – Liver Flush
În plus față de a scăpa de microorganismele nocive (și deșeurile lor extrem de acide), este de asemenea necesar de a intari flora intestinala(microorganisme benefice). Dupa ce scapati de microbii nocivi, ar trebui să utilizati un produs probiotic de calitate pentru a obține bacterii bune înapoi în corp(inclusiv muraturi precum varza murata, etc).
În timp ce produsele reziduale de microbii bune nu sunt neaparat alcaline, fac crea un mediu bun în creier.
90% din serotonina este produsa in stomac. Serotonina este numita si „hormonul fericirii”, de unde si expresia : „Intestine fericite, minte fericita”
Amintiți-vă ca scopul principal de a scăpa de microbi este de a elibera funcția de semnalizare radio a creierului.
În timp ce este foarte important de a rezolva si problema metalelor grele si aluminiului, etc., toxine sunt cauza dominanta de Alzheimer in aproape toate cazurile.
Alzheimer, aluminiu, mercur , plumb si alte metale grele
Plumbul poate contribui la Alzheimer, dar mercurul este, probabil, un factor mai mare.
„Cantitatea maximă de mercur pe care Agenția pentru Protecția Mediului permite oamenilor să fie expuși la este de 5000 de ori mai mică decât valoarea admisă de expunerea la plumb;cu alte cuvinte, APE aparent consideră mercurul să fie de 5000 de ori mai toxic decât plumbul. „Marcia Basciano DDS la reuniunea anuala a IAOMT San Diego 1994.” http://www.whale.to/a/toxic_dentistry.html
Deci, de la ce provine mercurul?
„Oamenii de știință au demonstrat că urme de mercur poate provoca tipul de deteriorare a nervilor, care este caracteristică prejudiciului constatat în boala Alzheimer. Nivelul de expunere la mercur utilizat în test a fost cu mult sub nivelurile ce se găsesc în plombele cu amalgame mercur / argint a multor oameni . Cercetările efectuate la Universitatea din Calgary , Facultatea de Medicina a constatat ca expunerea la mercur a cauzat formarea de „incurcaturi neurofibrilare”, care sunt unul dintre cei 2 markeri de diagnostic pentru boala Alzheimer.Studiile anterioare au aratat ca mercurul poate provoca si formarea celuilalt marker Alzheimer, „placi amiloide.” Cercetătorii au expus și testat nervii si la alte elemente, incluzând aluminiu, dar a constatat că numai mercurul a cauzat prejudiciul în concordanță cu boala Alzheimer. „http://www.whale.to/m/Alzheimers.html
Amalgamul dentar, plombele argintii pe care stomatologi le folosesc, poate fi o cauza semnificativa de Alzheimer si dementa.
„La nivel mondial există peste 4000 de lucrări de cercetare ce indică mercur ca fiind o substanță extrem de toxică.Cum pot fi atât de neatenti stomatologi pentru a plasa unul dintre cele mai mortale toxine existente la „doi” inch din creierul nostru? „Tom Warren (http://www.whale.to/a/toxic_dentistry.html)
Cât de mult mercur se pune de stomatologi în gură?
„Dr.Richard D. Fischer, un dentist ce practica în Annandale, VA și fostul presedinte al Academiei Internaționale de Medicină Orală și Toxicologie, a depus mărturie sa în fața Congresului la 14 noiembrie 2002. Dr. Fischer a publicat numeroase lucrări științifice și în 1998 a fost enumerat în Cine e Cine în Medicină și Sănătate.Acestea sunt extrase din mărturia sa: „plombele cu amalgame dentare (” argintii „) contribuie cu mai mult mercur la sarcina corpului la om decât toate celelalte surse (dieta, aer, apă, vaccinuri, etc.) combinate. Aceste umpluturi conțin 50% mercur – care este de o mai mare neurotoxicitate decat plumb, cadmiu, sau chiar arsenic. Pentru a pune acest lucru în perspectivă, cantitatea de mercur conținută într-o umplere mărime medie depășește standardul USEPA pentru expunerea la om de peste 100 de ani. Exprimat în alți termeni, este nevoie de doar ½ gram de mercur (o plomba) pentru a contamina toți peștii dintr-un lac de 10 acri. „http://www.gifam.org/article-4.htm
Amalgamul dentar este clar o sursă de mercur, așa cum este peștele în unele cazuri, dar există și alte surse. Acesta este un citat dintr-un raport despre mercur și aluminiu:
„Potrivit Hugh Fudenberg, MD (http://members.aol.com/nitrf), cel mai mare immunogeneticist din lume și al 13-lea cel mai citat biolog al vremurilor noastre (aproape 850 de lucrări în reviste de evaluare inter pares), în cazul în care o persoană a avut cinci vaccinuri consecutive gripa între 1970 și 1980 (anii studiati) sansele sale de a face boala Alzheimer este de zece ori mai mare decât în cazul în care au avut 1-2 sau deloc.L-am întrebat Dr. Fudenberg de ce acest lucru a fost așa și el a spus că a fost din cauza mercurului și aluminiului, care este în fiecare vaccin antigripal (și de acele vaccinuri din copilarie).Mercur și aluminiu treptat se acumuleaza in creier si cauzeaza disfunctii cognitive. „ http://www.whale.to/vaccines/flu11.html
Combinația dintre aluminiu și fluorură -cauza Alzheimer
Problema aluminiu si boala Alzheimer are nevoie de mai mult de discuții!
„Evident boala Alzheimer a venit după ce oamenii au inceput sa foloseasca vase de aluminiu.Isabel Jansen, RN, a scris un experiment simplu privind utilizarea de vase de aluminiu, unde este ușor de dovedit că aluminiul și conținutul de fluor in apa, ambele cresc in mod dramatic . „In ianuarie 1987, experimentele efectuate la Medical Research Departamentul Endocrinologie, Newcastle , Tyne, Anglia, și Dept Fizica a Univ. de Ruhana, Sri Lanka, au aratat ca apa cu fluor la 1 ppm, atunci când este utilizata la gatit in vase de aluminiu, creste concentratia de aluminiu până la 600 ppm, în timp ce apa fără fluor nu. (Science News 131: 73) Ei au sugerat ca, din cauza cunoscuta ca aluminiul este neuro-toxic si este în concentrații anormal de mari in creier si in alte victime de boli neurologice, inclusiv SIDA si Alzheimer , aceste constatari ridica semne de intrebare cu privire la adăugarea de fluor la alimentarea cu apă potabila a comunităților pentru a reduce cariile dentare (similar paste de dinti)…
Datorita acestor constatari, un test a fost facut din Antigo, Wisconsin , pe apă in care a fost introdus fluor timp de 33 de ani. Apa a fost examinat de un laborator certificat Wisconsin, si a aratat ca, atunci cand a fost folosit la gatit in vase de aluminiu, se concentrează aluminiul de 833 ori și a crescut conținutul de fluor cu 100% (Nota:. Eliminati vasele de gatit din aluminiu și rezolvati cumva si problema fluorului din apa (macar testati apa)!!) Conținutul maxim permis de aluminiu a apei este stabilit de Organizația Mondială a Sănătății, la 200 de micrograme pe litru. Acest lucru face ca apa Antigo , atunci când este gătită impreuna cu aluminiu, sa aiba nivelul aluminiu de 75 de ori peste nivelul maxim. Nici un test nu a fost realizat cu apă distilată, după cum Antigo Apa Departamentul nu eliberează și apă distilată. „ http://www.whale.to/a/alz.html
Aluminiu nu este din punct de vedere tehnic un metal greu, dar este atât de toxic incat mulți oameni il clasifica impreuna cu metale grele, ca și mine. Aș putea continua, dar cred ca e destul : plumb, mercur, aluminiu și fluor, și alte metale grele, pot fi o cauză majoră de Alzheimer.
Metalele grele ajung în organism prin diferite moduri, dar în primul rând :
1) amalgame dentare (mercur),
2) ustensile de gătit (aluminiu cu apa cu fluor, vase de fier),
3) Vaccinări (mercur),
4) Alimentele pe care le consumam si lucrurile pe care le bem (mercur, aluminiu, etc.),
5) aerul pe care il respiram, adică poluare (plumb și multe altele),
Boala „Alzheimer”, „a existat doar de aproximativ 25-30 de ani.(a nu se confunda cu dementa) pacientii cu Alzheimer dupa autopsie au avut în mod constant nivelul de aluminiu din SNC ridicat.Utilizarea pe scara larga de cutii de aluminiu din băuturi bere și răcoritoare a început acum aproximativ 30 de ani.„Întâmplător” industria aluminiului a lansat recent o campanie de publicitate TV de preempțiune scump citând toate multe beneficii de recipiente de aluminiu pentru utilizare in depozitarea alimentelor.Sloganul lor spune „nimic pastreaza alimentele proaspăt decât aluminiul.” [Fără îndoială, în speranța de a ajunge la toți acei băuturi răcoritoare neinformati, directorii companiilor îmbuteliere care încă nu au auzit de această opțiune minunatade ambalaje noi!] APE prevede că apa publică avea 50ppb [parti pe miliard] de aluminiu, si ca băuturi conservate contin 6160ppb. „Site-ul poate fi inaccesibil
Cauză Alheimer : Plumb
Există multe expuneri de uz casnic , în special în casele mai vechi. Cu toate acestea, este mult mai ușor de a studia expunerea la plumb la locul de muncă.
„Cercetare prezentata la reuniunea din mai 2000 a Academiei Americane de Neurologie a evidențiat o corelație dramatic între expunerea la plumb la locul de muncă și riscul de aparitie a bolii Alzheimer în anii următori.Istoriile profesionale a 185 de persoane cu boala Alzheimer au fost comparate cu 303 de oameni, fara boala Alzheimer.Rezultatele au aratat ca persoanele aveau de pana la 3,4 ori mai multe sanse de a dezvolta Alzheimer, dacă ar fi lucrat în locuri de muncă cu expunere la un nivel ridicat de plumb – fie prin inhalarea prafului de plumb sau prin contactul direct cu pielea „. http://www.fi.edu/brain/metals.htm
Și un alt articol de pe plumb și Alzheimer:
„Un studiu de patru ani a analizat 535 de foști angajați de fabrica chimică, care au avut o medie de opt ani de expunere la plumb la locul de muncă și o medie de 16 ani de la ultima expunere plumb .Nivelurile sanguine și osoase de plumb au fost măsurate și teste neurologice s-au efectutat – și în comparație cu 118 de persoane care nu sunt expuse .Muncitorii expusi la plumb nu a avut doar scaderi mai mari in rezultatele testelor, dar, de asemenea, declin în functiile creierului normale legate de vârstă .„Efectele nivelului mediu de plumb gasite in oasele foștilor lucrători cu plumb au fost : cinci ani îmbătrânire mai avansata asupra creierului”, a spus dr Schwartz. „http://www.fi.edu/brain/metals.htm
Cauza Alzheimer: medicamente chimice
Aici este o altă listă lungă de elemente. Rețineți că aspirina si paracetamol sunt menționate în acest articol:
„Acum un an am aflat că ALDEHIDE, cum ar fi acetaldehida păreau să agraveze simptomele de ALS.De asemenea, am citit un raport de materiale periculoase precum mercur care a avertizat împotriva utilizării acetilenă în prezența mercurului.În afacerea mea am folosit o cantitate substanțială de acetonă ca solvent de curățare.(Am folosit, de asemenea, o mare cantitate de xilen, despre care voi explica mai târziu) Cuvintele mă bântuie;Acetona, acetaldehida, acetilenă, și altele care ar putea fi mult mai familiare pentru dvs, acid acetilsalicilic (Aspirina) acetaminofen (paracetamol) și poate chiar N-acetil cisteina.După ce faci ceva mai multe sapaturi/cercetari, am constatat că toate aceste substanțe (cu posibila excepție a N-acetil cisteina, deoarece nu am putut găsi defalcarea chimice oriunde.) au un lucru în comun tulburător:Toate au un grup metil CH3 atașat … „site poate fi inaccesibile
Oprirea daunelor
Desigur dieta este esențială pentru a face față cu Alzheimer. Aspartam, MSG, proteine vegetale hidrolizate (HVP), cisteina și altele, trebuie neapărat să fie eliminate din dieta unei persoane cu Alzheimer.
De asemenea, toate oalele și tigăile si foliile si ambalajele bauturi din aluminiu ar trebui să fie eliminate Toate cutiile din aluminiu trebuie îndepărtate. Tot plasticul ar trebui îndepărtat (Notă:. La partea de jos a tuturor containerelor din plastic este un triunghi cu un număr în ea – nici un om ar trebui să consume un produs dintr-o sticlă de plastic cu un număr de 3 sau mai mare Doar un 1 sau 2 sunt mai sănătoși. Dacă nu există triunghi cu un număr, nu-l folositi!)
Sau optati pentru un filtru de apa
De asemenea, după cum sa menționat mai sus, drojdiile și ciupercile ar trebui omorate prin diverse suplimente. Acest lucru este foarte important și vor functiona în colaborare cu eliminarea tuturor zahărurilor si făinurilor și produselor lactate, practic, toate alimentele rafinate !!
Politica bolii
Persoana normala(asa cum am fost si eu odata, persoana nascuta si educata de societatea „moderna” care plateste asigurare medicala acestei societati, societate in care medicii se duc la facultate depun un juramant ca vor avea grija de sanatatea oamenilor bolnavi, etc si care normal, cand are o problema de sanatate isi pune toata increderea in medicina alopata pentru care se aloca bani grei si pentru ca ar trebui sa aiba oameni instruti, asta pina cand vede ca realitatea contrazice de multe teoria predata in scoli de medicina , mai in cazul bolilor unde se afirma „nu se cunoaste cauza, asa ca avem nevoie de mai multi bani pentru a cerceta ; intre timp; luati cimicalul x „) nu are absolut nici o idee despre ceea ce se întâmplă în ceea ce privește boala.
Corporațiile gigant care fac intenționat oale din aluminiu si tigai, altele raspandesc produse chimice mortale și periculoase pe plante, agențiile guvernamentale suprima adevărul despre pericolele de substanțe mortale, FDA pune oamenii în închisoare pentru tratarea cancerului, plantele modificate genetic apar peste tot, și așa mai departe.
Mass-media este de gardian al corupției, nevoia de bolnavi pe termen lung a concernelor farma etc etc.
Numai „teorii conspirationiste”, ce sa mai. Nu mai intru in detalii . O mai spun pe cea finala:
Asa cum EXISTA DUMNEZEU, exista si diavol si nu te joci cu diavolul.
Dar sa nu uitam ca EXISTA DUMNEZEU!!!
Revin la tratamentul altor afecțiuni cerebrale precum Alzheimer ce poate fi un proces în două etape. Primul pas este identificarea a ceea ce este cauza prejudiciului. Daca paguba este aluminiu sau mercur, de exemplu, chelarea de metale grele poate fi necesară (a se vedea capitolul următor).
Dacă problema sunt celulele creierului moarte, produsul Aloe imunitar (pe care FDA l-a persecutat puternic ca daor e planta, face rau!!!), care inunda corpul cu celule stem naturale, poate ajuta organismul la regenerarea de noi celule ale creierului.Si lucruri precum turmericul pot avea impact in acest sens
Pentru a restaura energia celulară o serie de lucruri, mai ales discutate în capitolul următor, pot ajuta. Exemple sunt suplimente cheie, Dieta Budwig, etc.
Pe scurt, trăim într-o lume coruptă în care se pare că aproape toată lumea este interesată de bani, dar nimeni nu este interesat de sănătatea dumneavoastră. Desigur, medicina alopata de multe ori nu da 2 lei ref sanatatea dvs (acest lucru este, la urma urmei, un site de cancer și pacienții cu cancer, de asemenea, care au invatat această lecție cu mari greutati;desi sunt convins ca exista „O GRAMADA” de medici care sunt convinsi ca fac bine ce fac(pentru ca asa ii invata in scoala) , ar fi bine ca acesti medici sa isi puna niste intrebari in momentul in care oamenii mor pe banda rulanta si se sustine cu nerusinare de catre scoli ca boli precum cancerul sunt incurabile, desi exista nu numai mii de argumente si dovezi stiintifice in sens contrar ci si cazuri reale).
Tot ce puteti face este să învațati elementele de bază și sa faceti două lucruri:
1) Analiza la ceea ce pacientul a fost expus (de exemplu vase de aluminiu și tigăi, bea o mulțime de băuturi răcoritoare, lucrează cu substanțe chimice, etc.), Precum și
2) Încercați diferite lucruri pentru a vedea ce funcționează in cazul dvs.
Următoarele câteva capitole sperăm că va oferi mai multe detalii pentru a vă ghida
Alzheimer eBook
Capitolul 1: Care sunt cauzele bolii Alzheimer? (Acest articol)
Aceasta este o carte excelentă cu o mulțime de informații despre Alzherimer lui, studii de caz și o gamă largă de idei.Acest lucru este foarte recomandat:
1Departamento de Docencia e Investigación, Facultad de Ciencias Médicas, Pontificia Universidad Católica Argentina, Buenos Aires 1007, Argentina
2Centro de Neuropsiquiatría y Neurología de la Conducta, Hospital de Clínicas „José de San Martín”, Facultatea de Medicina, Universidad de Buenos Aires, Buenos Aires 1121, Argentina
Boala Alzheimer (AD) este o problemă majoră de sănătate și există o recunoaștere din ce în ce mai mare că eforturile de prevenire a acesteia trebuie întreprinse atât de organizații guvernamentale, cât și de organizații neguvernamentale. În acest context, produsul pineal, melatonina, are o semnificație promițătoare datorită proprietăților sale cronobiotice/citoprotectoare potențial utile pentru o serie de aspecte ale AD. Una dintre caracteristicile înaintării în vârstă este scăderea treptată a nivelului circulant de melatonina. Un număr limitat de studii terapeutice au indicat că melatonina are o valoare terapeutică ca medicament neuroprotector în tratamentul AD și afectarea cognitivă minimă (care poate evolua către AD). Atât in vitro, cât și in vivo, melatonina a prevenit neurodegenerarea observată în modelele experimentale de AD. Pentru ca aceste efecte să apară, sunt necesare doze de melatonină cu aproximativ două ordine de mărime mai mari decât cele necesare pentru a afecta somnul și ritmicitatea circadiană. Mai recent, atenția s-a concentrat asupra dezvoltării de analogi potenți ai melatoninei cu efecte prelungite, care au fost folosiți în studiile clinice la pacienții cu tulburări de somn sau depresive în doze considerabil mai mari decât cele utilizate pentru melatonina. Având în vedere că potențele relative ale analogilor sunt mai mari decât cele ale compusului natural, studiile clinice care utilizează melatonina în intervalul 50-100 mg/zi sunt urgent necesare pentru a evalua validitatea sa terapeutică în tulburările neurodegenerative precum AD
Abstract grafic
1. Introducere
Tulburările neurodegenerative sunt un grup de boli cronice și progresive caracterizate prin pierderi selective și simetrice ale neuronilor în sistemele cognitive, motorii sau senzoriale. Boala Alzheimer (AD) este reprezentantul cel mai relevant din punct de vedere clinic al acestui tip de tulburări. Deși originea neurodegenerării specifice în AD rămâne nedefinită, trei procese majore și frecvent interconectate, și anume deteriorarea mediată de radicali liberi, disfuncția mitocondrială și excitotoxicitatea, stau la baza mecanismelor fiziopatologice care duc la moartea neuronală [1
] .AD are un impact foarte mare asupra sănătății. Într-o publicație recentă [
2] organizația Alzheimer’s Disease International (ADI) afirmă că 44 de milioane de oameni trăiesc cu demență în prezent și că această cifră va crește la 135 de milioane de oameni până în 2050. Costul economic actual al demenței este de 604 miliarde de dolari anual. Trebuie menționat că noile estimări reprezintă o creștere de 17% față de cifrele publicate în 2009, îmbătrânirea populației fiind principalul motor al creșterii proiectate. Raportul ADI prezice, de asemenea, o schimbare în distribuția poverii globale a demenței până în 2050, 71% dintre toate persoanele afectate trăind în țări cu venituri mici sau medii. Se estimează că 10% din cazurile de demență pot fi evitate prin campanii care vizează obezitatea, hipertensiunea, diabetul, subactivitatea și fumatul, precum și prin educație și îmbunătățirea cognitivă. Prin urmare,Aportul regulat de antioxidanți de către vârstnici a fost una dintre strategiile recomandate pentru prevenirea bolilor neurodegenerative asociate vârstei, mediate de radicali liberi, dar eficacitatea acestui tratament este dezbătută [3
] . În acest context, utilizarea melatoninei ca agent citoprotector devine de mare interes. Melatonina este un metoxiindol bine conservat care se găsește în majoritatea filelor și are acțiuni citoprotectoare remarcabile pe lângă proprietățile cronobiotice. Sursa melatoninei circulante este glanda pineală, iar o cantitate substanțială de date susține că scăderea melatoninei plasmatice este una dintre caracteristicile înaintării în vârstă [
4 ].]. Vom trece în revistă pe scurt câteva aspecte relevante ale biologiei melatoninei în ceea ce privește procesul neurodegenerativ. Apoi, vor fi discutate teoria și modul de acțiune al melatoninei în AD. În cele din urmă, studiile clinice care susțin utilizarea terapeutică a melatoninei în AD vor fi evaluate critic.
2. Biologia de bază a melatoninei relevante pentru neurodegenerare
Melatonina circulantă la mamifere derivă aproape exclusiv din glanda pineală [
5 ]. În plus, melatonina este sintetizată local în multe celule, țesuturi și organe, inclusiv limfocite, măduvă osoasă, timus, tractul gastrointestinal, piele și ochi (vezi [6,
) . Faptul că apariția melatoninei nu este limitată la vertebrate, ci este prezentă aproape omniprezent în numeroși taxoni, inclusiv, de exemplu, bacterii, eucariote unicelulare și plante [6, 8], subliniază că această moleculă a câștigat
suplimentare în cursul evolutiei.Atât la alte animale, cât și la oameni, melatonina participă la diverse funcții fiziologice semnalând nu numai durata nopții, ci și facilitând, printre altele, eliminarea radicalilor liberi și răspunsul imun, prezentând astfel proprietăți citoprotectoare relevante [6
] . Într-adevăr, citoprotecția poate explica bine prezența melatoninei în majoritatea organismelor vii. Funcțiile sistemice și intracelulare ale melatoninei diferă în cantitățile necesare de agonist. În general, efectele melatoninei asupra ritmurilor biologice sunt exercitate la concentrații nanomolare și prin mecanisme mediate de receptor. Efectele citoprotectoare ale melatoninei necesită cu 100–1000 de concentrații mai mari, compatibile cu cantitățile atinse intracelular de melatonina în mai multe țesuturi [
9 ] și este metabolizată în principal în ficat, în principal prin hidroxilare la poziția C6. Acesta este catalizat selectiv de citocromul microzomal hepatic P450 1A2 (CYP1A2), cu contribuții minore din CYP2C19 hepatic și CYP1A1 și CYP1B11A în mare parte extrahepatic [
12 ]. 6-hidroximelatonina formată este apoi conjugată cu sulfat sau glucuronid pentru a fi excretată în urină. Un alt metabolit important este
N -acetilserotonina care se formează prin
O -demetilare și poate reprezenta până la 20% din doza administrată de melatonină [
13 ].Melatonina este, de asemenea, metabolizată în țesuturi prin scindarea inelului pirolic oxidativ în derivați de chinuramină. Produsul primar de scindare este
N1 – acetil-
N2 -formil-5-metoxichinuramina (AFMK), care este deformilată, fie de arilaminformamidază, fie de hemoperoxidază, la
N1
– acetil -5-metoxicinuramină (AMK) [
14 ]. S-a propus că AFMK este metabolitul activ primar al melatoninei pentru a media citoprotecția [
15 ]. Melatonina este, de asemenea, transformată în 3-hidroximelatonină ciclică într-un proces care curăță direct doi radicali hidroxil [
15]. Datorită metabolismului său rapid, melatonina are un timp de înjumătățire scurt de aproximativ o oră, deși există o variație inter-individuală marcată a nivelurilor plasmatice ale melatoninei după administrarea orală [16
2 se găsesc în membrana celulară ca dimeri și heterodimeri. GPR50, un ortolog al receptorului melatoninei cuplat cu proteina G care nu leagă melatonina în sine, dimerizează cu receptorii MT
Receptorul MT2 uman prezintă o afinitate mai mică decât receptorul MT1 uman
şi devine desensibilizat după expunerea la melatonină, probabil prin internalizare.Ca reprezentanți ai familiei de receptori cuplati cu proteina G, receptorii MT
1 și MT
2 acționează printr-un număr de mecanisme de transducție a semnalului [
1 este cuplat la proteinele G care mediază inhibarea adenililciclazei și activarea fosfolipazei C. Receptorul MT2 este, de asemenea, cuplat la inhibarea adenilil ciclazei și, în plus, inhibă calea guanilil ciclază solubilă
.Prin utilizarea autoradiografiei receptorilor cu ligand neselectiv 2-[125I]iodomelatonină și reacție cantitativă în lanț a polimerazei-transcripție inversă în timp real pentru a marca ARNm a receptorului melatoninei, s-au identificat receptorii MT
1 și MT
2 regiuni ale creierului. La nivelul hipocampusului, receptorii MT
2 au fost detectați în neuronii piramidali CA3 și CA4, care primesc inputuri excitatoare glutamatergice din cortexul entorinal, în timp ce receptorii MT
1 au fost exprimați predominant în CA1 [
6 ].Melatonina se leagă, de asemenea, la factorii de transcripție aparținând superfamiliei receptorilor acidului retinoic, în special, variantele de îmbinare ale RORα (RORα1, RORα2 și RORα izoforma d) și RZRβ [21
] . Subformele receptorului acidului retinoic sunt exprimate în mod omniprezent în țesuturile mamiferelor [
22 ].Melatonina este un antioxidant puternic care captează radicalii ∙OH, precum și alte specii de oxigen radical (ROS) și specii de azot radical (RNS) și care dă naștere cascadei de metaboliți menționați mai sus care împărtășesc proprietăți antioxidante [23
] . Melatonina acționează, de asemenea, indirect pentru a promova expresia genică a enzimelor antioxidante și pentru a inhiba expresia genică a enzimelor prooxidante [
27 ]. În special, acest lucru este valabil pentru glutation peroxidază (GPx) și pentru glutation reductază (GRd), probabil ca răspuns la creșterile dependente de GPx ale GSSG, forma oxidată a glutationului (GSH). Melatonina contribuie la menținerea nivelurilor normale de GSH a creierului [
28] prin stimularea biosintezei GSH prin γ-glutamilcisteina sintetază și glucozo-6-fosfat dehidrogenază [
29 ]. Melatonina are o superioritate demonstrată față de vitamina C și E în protecția împotriva daunelor oxidative și în eliminarea radicalilor liberi [
23 ].Melatonina are proprietăți antiinflamatorii semnificative, probabil prin inhibarea legării factorului nuclear κB (NFκB) de ADN, scăzând astfel sinteza citokinelor proinflamatorii prin inhibarea ciclooxigenazei (Cox) [30], în principal
33 ]. Deși acțiunea directă a melatoninei ca agent antioxidant este în mare parte independentă de interacțiunea receptorului [
34 ], reglarea în sus a enzimelor antioxidante de către metoxiindol implică transcripția nucleară și, în unele cazuri, receptorii RZR/RORα [
35] .].Eficacitatea melatoninei în inhibarea daunelor oxidative a fost testată într-o varietate de modele de boli neurologice. În plus față de modelele animale de AD discutate mai jos, s-a demonstrat că melatonina reduce daunele neuronale din cauza toxicității cadmiului [
51].Proprietățile neuroprotectoare ale melatoninei, precum și efectele sale reglatoare asupra tulburărilor circadiene, validează beneficiile acesteia ca substanță terapeutică în tratamentul preventiv al AD. Mai mult, melatonina exercită efecte anti-excitatoare și, la doze suficiente, efecte sedative [
53 ], astfel că poate exista un al doilea mod de acțiune neuroprotector care implică sistemul ergic de acid y-aminobutiric (GABA) ca mediator. Acest punct de vedere este susținut de studii care indică faptul că melatonina protejează neuronii de toxicitatea peptidei amiloid-β (Aβ) (o neurotoxină principală implicată în AD) prin activarea receptorilor GABA [54
] .Studiile timpurii privind acțiunile anti-excitotoxice ale melatoninei au folosit kainatul, un agonist al receptorilor ionotropi de glutamat și au susținut ipoteza că melatonina previne moartea neuronală indusă de aminoacizii excitatori [55
] . De asemenea, s-a raportat că administrarea de melatonină reduce leziunile neuronilor CA1 hipocampali cauzate de ischemia tranzitorie a creierului anterior [
59 ].Diferitele tipuri de toxicități enumerate mai sus pot duce la moarte neuronală prin necroză sau apoptoză. Moartea neuronală apoptotică necesită sinteza de ARN și proteine și epuizarea factorilor trofici. Apoptoza implică, de asemenea, rupturi monocatenar ale ADN-ului și s-a descoperit că factorii neurotrofici salvează neuronii de acest tip de moarte [
60 ]. Ele pot acționa prin intermediul componentelor celulare anti-apoptotice, cum ar fi proteina proto-oncogenă a limfomului B (Bcl-2).
Studiile in vitro indică faptul că melatonina îmbunătățește expresia Bcl-2 și previne apoptoza [
66].Procesul de îmbătrânire al creierului este cunoscut în prezent suficient de detaliat [
67 ]. O serie de procese de senescență se bazează pe deteriorarea mitocondrială, inclusiv apoptoza prin peroxidarea cardiolipinei, eliberarea citocromului C și defalcarea mtPTP și reducerea masei mitocondriale. Blocarea lanțului de transport de electroni duce la o aprovizionare insuficientă cu energie și la afectarea funcției și viabilității celulelor. Generarea crescută de ROS și RNS provoacă leziuni concomitente la endotelii, proteinele ADN și lipide. În plus, o supraexcitare neuronală cu supraîncărcare cu calciu și activarea microgliei apar odată cu vârsta. Ca o consecință a deteriorării ADN-ului, are loc o uzură crescută a telomerilor împreună cu o reducere a celulelor cu capacitate proliferativă ridicată, cum ar fi celulele progenitoare imune, ducând astfel la imunosenescență [67]
.]. Această situație este însoțită de creșterea răspunsurilor autoimune și de un proces inflamator care duce la creșterea citokinelor proinflamatorii.Dovezile derivate din studiile pe animale indică faptul că melatonina poate reduce majoritatea aspectelor îmbătrânirii creierului [
67].]. În primul rând, prin acțiunea asupra oscilatorilor circadieni centrali și periferici, melatonina crește amplitudinile ritmului deprimat și coordonarea slabă a ritmurilor observate la persoanele în vârstă. Melatonina afectează expresia proteinei de ceas și poate modula perturbarea sensibilității metabolice în senescență. Prin susținerea fluxului de electroni mitocondriali și reducerea debordării electronilor, melatonina îmbunătățește eficiența respiratorie și aprovizionarea cu energie și previne apoptoza. Reducerea ROS și RNS exercitată prin eliminarea radicalilor, reglarea în sus a enzimelor antioxidante, inhibarea enzimelor prooxidante, creșterea GSH și formarea de radicali mai mici duce la o deteriorare redusă a proteinelor, lipidelor și ADN-ului, împreună cu inhibarea telomerilor induși de oxidanți. uzură și supraexcitare neuronală. Acțiunile antiinflamatorii ale melatoninei sunt cruciale în reducerea proceselor legate de îmbătrânire, precum și în îmbunătățirea sensibilității la insulină și a contracarării sindromului metabolic atribuit metoxiindolului. În cele din urmă, melatonina modulează imunitatea naturală și adaptivă și îmbunătățește imunosenescența, parțial printr-un număr mai mare de celule cu capacitate proliferativă ridicată, inclusiv leucocite, celule stem și celule progenitoare.
3. Prezentare generală a terapiei cu melatonina pentru boala Alzheimer – Teorie și mod de acțiune
Plăcile senile extracelulare, formate în principal din depozite de Aβ, și încurcăturile neurofibrilare intracelulare, rezultate în principal din proteina asociată microtubulilor (MAP) anormal hiperfosforilată, sunt caracteristica patologică majoră a AD. Ap joacă un rol important în promovarea degenerării neuronale în AD transformând neuronii vulnerabili la creșterile legate de vârstă ale nivelurilor de stres oxidativ și un metabolism energetic celular alterat.Aβ este compus din 39–43 de resturi de aminoacizi derivate din precursorul său, proteina precursor amiloid (APP) [
68 ]. APP este procesat proteolitic de către α- sau β-secretaze în diferite căi. Calea α-non-amiloidogenă implică scindarea APP de către α-secretază pentru a elibera un fragment
N -terminal al APP, care după scindarea de către γ-secretază împiedică formarea Ap [
68 ]. Calea β-amiloidogenă include β-secretaza, care are ca rezultat formarea peptidei Aβ intacte și este mediată de scindarea secvenţială a β-secretazei și γ-secretazei la N- și C-terminal al
secvenţei Aβ
[ 68
] .Melatonina a inhibat nivelurile normale de secreție de APP solubilă în diferite linii celulare interferând cu maturarea APP [
69 ]. În plus, administrarea melatoninei reduce eficient generarea și depunerea Ap
74 ]. În general, rezultatele la șoarecii transgenici susțin opinia conform căreia melatonina ar trebui administrată într-o fază timpurie pentru a regla metabolismul APP și Aβ în principal prin prevenirea formării lor, cu un efect anti-amiloid redus ulterior. Astfel, terapia cu melatonină la șoarecii bătrâni Tg2576 începând cu vârsta de 14 luni nu a putut preveni depunerea suplimentară de Aβ.
75 ] în timp ce un tratament similar începând cu vârsta de a patra lună a fost eficient pentru a o reduce [
70 ]. Deoarece patologia plăcii de amiloid este observată în mod obișnuit la șoarecii Tg2576 cu vârsta de 10 până la 12 luni [
76 ], datele indică eficacitatea melatoninei în prevenirea formării timpurii a plăcii de amiloid, mai degrabă decât ulterior.Modul în care melatonina își exercită efectul inhibitor asupra generării de Ap rămâne nedefinit. Scindarea proteolitică a APP de către calea α-secretazei este reglată de mulți stimuli fiziologici și patologici, în special prin activarea proteinei kinazei (PK) C și clivajul mediat de secretază a APP. Inhibarea glicogen sintazei kinazei-3 (GSK-3) și reglarea în sus a kinazei
N -terminale c-Jun au ca rezultat o activitate ridicată a metaloproteinazelor matriceale cu degradarea crescândă a Ap [
77 ]. Activitatea enzimei de degradare a insulinei (IDE) care reglează nivelurile de insulină, Aβ și APP, a scăzut după creșterea Apei [
78] .]. GSK-3 interacționează cu presenilin-1, un cofactor al y-secretazei, fosforilarea GSK-3 de către PKC ducând la inactivarea y-secretazei. Într-adevăr, GSK-3 poate fi una dintre căile de semnalizare comune, crescând generarea de Ap și hiperfosforilarea tau, iar melatonina ar putea regla procesarea APP prin căile PKC și GSK-3 (
80 ]. Această interacțiune între melatonină și Aβ pare să depindă mai degrabă de caracteristicile structurale ale melatoninei decât de proprietățile sale antioxidante, deoarece nu ar putea fi imitată de alți captatori de radicali liberi [
79 ]. Prin blocarea formării foilor secundare, melatonina nu numai că reduce neurotoxicitatea, ci și facilitează clearance-ul peptidei, crescând degradarea proteolitică, de exemplu, prin IDE.
Figura 1. Efectul melatoninei asupra semnalizării afectate a insulinei cerebrale în AD. Figura schematizează procesele care leagă deficiența insulină/receptorul de insulină cu plăcile senile și încurcăturile neurofibrilare. Între paranteze, acțiunea demonstrată a melatoninei, așa cum este discutată în text.Stresul oxidativ joacă un rol central în neurotoxicitatea indusă de Ap și moartea celulelor. Acumularea datelor susține că melatonina protejează eficient celulele împotriva daunelor oxidative induse de Aβ și a morții celulare
85.]. În celulele și animalele tratate cu Ap, melatonina și-ar putea exercita activitatea protectoare printr-un efect antioxidant, în timp ce în celulele transfectate cu APP și modelele animale transgenice, mecanismul de bază poate implica în primul rând inhibarea generării de foi de β și/sau fibrile de amiloid. Ap agregat generează ROS care produc moartea neuronală prin deteriorarea lipidelor membranei neuronale, proteinelor și acizilor nucleici. S-a observat protecție împotriva toxicității Aβ de către melatonină, în special la nivel mitocondrial [
89 ]. Amploarea patologiei neurofibrilare se corelează cu severitatea demenței la pacienții cu AD. Nivelul de tau hiperfosforilat este de trei până la patru ori mai mare în creierul pacienților cu AD decât în creierul adulților normali [
91 ].Melatonina atenuează eficient hiperfosforilarea tau prin afectarea protein kinazelor și fosfatazelor într-un număr de modele experimentale, inclusiv expunerea celulelor neuroblastomului N2a și SH-SY5Y la wortmannin [92], calyculin
101 ]. Inhibarea biosintezei melatoninei la șobolani nu numai că a dus la afectarea memoriei spațiale, dar a indus și o creștere a fosforilării tau, un efect prevenit de suplimentarea cu melatonina [102]
.].Se știe că stresul oxidativ influențează starea de fosforilare a tau [
104 ]. Acumularea de proteine greșite și agregate în neuronii creierului ai AD este considerată o consecință a stresului oxidativ, în plus față de modificările structurale moleculare datorate vârstei [
105 ]. Deoarece melatonina previne, ca antioxidant și captator de radicali liberi, supraproducția de radicali liberi, pare fezabil ca prevenirea fosforilării tau de către melatonina să aibă loc parțial datorită activității sale antioxidante. În plus, mai multe studii au indicat că melatonina poate acționa ca un modulator al enzimelor într-un mod care nu are legătură cu proprietățile sale antioxidante. Acestea includ reglarea de către melatonina a PKA [
120 ].Un alt eveniment major în patogeneza AD este deficitul funcției colinergice [
121 ]. Neuronii din nucleul bazal al lui Meynert, principala sursă de inervație colinergică a cortexului cerebral și a hipocampului, suferă o degenerare profundă și selectivă în creierul AD [
122 ]. Nivelurile de acetilcolină (ACh) sunt reduse în stadiul incipient al AD, în timp ce activitățile enzimelor de sinteză și degradare a colin acetiltransferaza (ChAT) și acetilcolinesteraza (AChE) nu se modifică până la o fază târzie a AD [123
] . Deoarece scăderea activității ChAT în neocortexul pacienților cu AD sa corelat cu severitatea demenței, inhibitorii AChE au devenit un tratament standard al AD ușoară până la moderată [125]
.].Melatonina are un efect protector asupra sistemului colinergic. Previne inhibarea indusă de peroxinitrit a transportului de colină și a activității ChAT în sinaptozomi și vezicule sinaptice [
126 ]. Tratamentul cu melatonină al șoarecilor transgenici APP695 în vârstă de opt luni a îmbunătățit semnificativ reducerea profundă a activității ChAT în cortexul frontal și hipocampus [
81 ]. Melatonina antagonizează, de asemenea, deficitul de memorie spațială și scăderea activității ChAT găsite la șobolanii adulți ovariectomizați [
127 ]. Cu toate acestea, la șobolanii perfuzați intracerebroventricular cu Aβ timp de 14 zile, melatonina nu a putut restabili activitatea ChAT [
128 ]. Melatonina a inhibat creșterea indusă de lipopolizaharide și streptozotocină a activității AChE [
129]. Recent, hibrizi ai inhibitorului AChE tacrină și melatonină au fost sintetizați ca noi candidați la medicamente pentru tratarea AD [
131 ]. Acești hibrizi au prezentat o activitate antioxidantă și de conservare colinergică mai bună decât tacrina sau melatonina în monoterapie. Administrarea directă intracerebrală a unuia dintre acești hibrizi a scăzut moartea celulară indusă și încărcarea Aβ în parenchimul creierului de șoarece APP/PS1, însoțită de o recuperare a funcției cognitive [
131 ].Există un interes din ce în ce mai mare pentru rolul semnalizării insulinei cerebrale afectate în patologia AD. S-a sugerat că perturbarea semnalizării creierului insulinei/factorului de creștere asemănător insulinei 1 (IGF-1) este un eveniment cauzal cheie care stă la baza AD, fiind legată atât de prezența plăcilor senile, cât și a încurcurilor neurofibrilare [132
] . Acest punct de vedere, însă, nu este universal susținut [
135 ]. O semnalizare afectată a insulinei/receptorului de insulină (IR) duce la scăderea activării mediate de insulină a activității de semnalizare fosfoinozitid 3-kinaze (PI-3K)/Akt, ducând la supraactivarea GSK-3 care promovează direct hiperfosforilarea tau și acumularea de Aβ și plăcile senile. formarea (
Figura 1). Activitatea IDE, care reglează nivelurile de insulină, Aβ și APP, a scăzut după depunerea de Aβ [
78 ]. Administrarea melatoninei reduce semnele sindromului metabolic, cum ar fi hiperglicemia, dislipidemia, hiperinsulinemia, rezistența la insulină, creșterea în greutate și hipertensiunea [
137 ]. Melatonina restabilește mecanismele receptorilor de insulină/insulină și crește activitatea de semnalizare a fosfoinozitidei 3-kinaze/Akt cu o inhibare rezultată a GSK-3 și mai puțină acumulare de Aβ și hiperfosforilare tau (vezi [ 6 )
.]). În plus, întreruperea semnalizării insulinei duce la o scădere a expresiei transportorului de glucoză-1 (GLUT-1) și -3 (GLUT-3), culminând cu absorbția/metabolismul glucozei cerebrale afectate, un alt eveniment contracarat de melatonină. Metabolismul neuronal restabilit al glucozei mărește acilarea tau
N -acetilglucozamină, reducând astfel hiperfosforilarea tau (
Figura 2 rezumă principalele ținte posibile pentru medicație în AD. Food and Drug Administration (FDA) a aprobat doar inhibitori de AChE și blocanți ai receptorilor
N -metil-
D -aspartat (NMDA) pentru uz clinic. După cum sa discutat mai sus, melatonina are proprietatea unică de a afecta toate mecanismele fiziopatologice descrise în
Figura 2 . Prin urmare, potențialul său ca neuroprotector în AD merită să fie explorat.
Figura 2. Ținte posibile pentru medicație în AD. După cum se discută în text, melatonina are potențialul de a afecta toate mecanismele descrise în figură. (*) Aprobat de FDA.
4. Aspecte clinice ale aplicației melatoninei în AD
La pacienții cu dementa, severitatea deficienței mintale s-a corelat semnificativ cu producerea de melatonină nocturnă afectată [ 138 ]. Acesta poate fi un semn precoce al bolii, deoarece nivelurile de melatonina din lichidul cefalorahidian (LCR) sunt reduse la pacienții cu AD autopsiați în stadiul preclinic, stadiul Braak-1. Prin urmare, reducerea melatoninei LCR poate fi un marker pentru detectarea precoce a AD [ 139 ].În plus, nivelurile plasmatice ale melatoninei sunt mai scăzute la pacienții cu AD [ 140 , 141 , 142 , 143 ] și s-au postulat că conexiunile retiniene-SCN-pineale afectate funcțional sunt cauza acestei scăderi [ 144]. Expresia și activitatea genei monoaminoxidazei au crescut, iar nivelurile de ARNm a adrenoceptorului β1 au scăzut, în glanda pineală a pacienților cu AD [ 145 ]. În plus, au apărut modificări în imunoreactivitatea receptorului de melatonină în hipocampul AD [ 146 , 147 ]. În mod colectiv, rezultatele indică faptul că înlocuirea nivelurilor de melatonină poate fi o strategie terapeutică pentru oprirea progresiei AD.„Apusul soarelui”, un fenomen cronobiologic care include gândirea dezorganizată, capacitatea redusă de a menține atenția la stimulii externi, rătăcirea, agitația și tulburările perceptuale și emoționale, este o situație observată frecvent la pacienții cu AD. Ca fenomen legat de timpul zilei, care apare după-amiaza târziu sau seara devreme [
149 ], posibilul efect terapeutic al melatoninei a fost întreținut.
Tabelul 1 rezumă datele care indică faptul că melatonina, ca agent cronobiotic, poate fi eficientă în tratarea apusului soarelui și a ciclurilor neregulate de somn-veghe la pacienții cu AD.O revizuire a rezultatelor publicate privind utilizarea melatoninei în AD [ 150 ] a dat opt rapoarte (cinci studii deschise, două rapoarte de caz) ( N = 89 de pacienți) care susțin o posibilă eficacitate a melatoninei: calitatea somnului s-a îmbunătățit și la pacienții cu apus de soare. a fost redusă și decăderea cognitivă a încetinit progresia. În șase studii dublu-orb, randomizate controlate cu placebo, au fost examinați un număr total de 210 de pacienți cu AD. Calitatea somnului a crescut și apusul soarelui a scăzut semnificativ, iar performanța cognitivă s-a îmbunătățit în patru studii ( N = 143), în timp ce a existat absența efectelor în două studii ( N = 67) [ 150 ].O altă căutare sistematică a studiilor publicate între 1985 și aprilie 2009 privind melatonina și apusul soarelui la pacienții cu AD a fost publicată [ 151 ]. Toate lucrările despre tratamentul cu melatonina în demență au fost preluate și efectele melatoninei asupra tulburărilor de ritm circadian au fost notate prin evaluarea comportamentului de apus/agitat, calitatea somnului și funcționarea în timpul zilei. Au fost revizuite un total de 9 lucrări, inclusiv 4 studii randomizate controlate ( N = 243) și 5 serii de cazuri ( N = 87) ( Tabelul 1 ). Două dintre studiile controlate randomizate au constatat o îmbunătățire semnificativă a comportamentului de apus/agitat. Toate cele cinci serii de cazuri au găsit o îmbunătățire [ 151]. Astfel, dacă melatonina are vreo valoare în tratarea AD rămâne incertă. Probabil, eterogenitatea grupului examinat în AD complet dezvoltat face dificilă dezvăluirea oricărui efect terapeutic în studiile care includ un număr mic de pacienți [
152 ]. Mai mult, expresia hipocampală redusă a receptorilor de melatonină MT 2 la pacienții cu AD [ 146 ] și a receptorilor MT 1 din aparatul circadian poate explica de ce tratamentul cu melatonina este neregulat în acest stadiu avansat al bolii [ 153 , 154 ].
Tabel 1. Studii care includ tratamentul pacienților cu boala Alzheimer (AD) cu melatonină.
Deficiența cognitivă ușoară (MCI) este denumirea dată semnelor și simptomelor diagnosticate la cei care au un deficit obiectiv și măsurabil în funcțiile cognitive, dar cu o relativă conservare a activităților zilnice. Ratele anuale de conversie la AD sunt de până la 10%–15% [
169 ] și MCI reprezintă o etapă importantă clinic pentru identificarea și tratarea persoanelor cu risc. Într-adevăr, procesul degenerativ din creierul AD începe cu 20-30 de ani înainte de debutul clinic al AD [
173 .]. După cum sa menționat mai sus, nivelurile de melatonina LCR scad în stadiile preclinice ale AD, ceea ce sugerează că reducerea melatoninei LCR poate fi un declanșator și un marker precoce pentru AD [ 139 , 145 ]. Deși nu se știe dacă deficiența relativă de melatonină este fie o consecință, fie o cauză a neurodegenerării, pierderea de melatonină probabil agravează boala.Am raportat anterior o analiză retrospectivă în care 3-9 mg zilnic dintr-un preparat de melatonină cu eliberare rapidă per os (po) la culcare timp de până la trei ani au îmbunătățit semnificativ performanța cognitivă și emoțională și ciclul zilnic de somn/veghe la 25 de pacienți cu MCI [174] . Recent am raportat date de la o altă serie de 96 de pacienți cu MCI, dintre care 61 au primit zilnic 3-24 mg de un preparat de melatonină cu eliberare rapidă po, la culcare, timp de 15 până la 60 de luni, în comparație cu un grup similar de 35 de pacienți cu MCI care nu au primit.[ 175 ]. În plus, toți pacienții au primit medicația standard individuală considerată adecvată de către medicul psihiatru curant.Pacienții tratați cu melatonină au prezentat performanțe semnificativ mai bune la mini-examenul stării mentale și la subscala cognitivă a scalei de evaluare a AD. După aplicarea unei baterii neuropsihologice care cuprinde un test Mattis, testul Digit-simbol, sarcinile Trail A și B și testul verbal Rey, s-au găsit performanțe mai bune la pacienții tratați cu melatonină pentru fiecare parametru testat [175 ] . Scorurile anormal de mari ale Beck Depression Inventory au scăzut la pacienții tratați cu melatonină, concomitent cu îmbunătățirea calității somnului și a stării de veghe [ 175 ]. Aceste rezultate susțin în continuare că melatonina este un medicament suplimentar util pentru tratarea MCI într-un mediu clinic.Astfel, o inițiere precoce a tratamentului cu melatonină poate fi decisivă pentru succesul terapeutic [ 75 ]. În Tabelul 2 , sunt rezumate datele publicate privind tratamentul cu melatonină în MCI. Șase studii dublu-orb, randomizate controlate cu placebo și două studii retrospective deschise ( N = 782) au arătat în mod constant că administrarea zilnică de melatonină seara îmbunătățește calitatea somnului și performanța cognitivă la pacienții cu MCI. Prin urmare, tratamentul cu melatonină ar putea fi eficient în stadiile incipiente ale bolii neurodegenerative.Există două motive pentru care este benefică utilizarea melatoninei la pacienții cu MCI. În cursul procesului neurodegenerativ, deteriorarea legată de vârstă în organizarea circadiană devine semnificativ exacerbată și este responsabilă de probleme de comportament precum apusul soarelui [
176 ]. Declinul cognitiv legat de vârstă la adulții în vârstă sănătoși poate fi prezis prin fragmentarea ritmului circadian în comportamentul locomotor. Prin urmare, înlocuirea nivelurilor scăzute de melatonină care apar în creier [ 139 , 145 ] poate fi foarte benefică la pacienții cu MCI.Pe de altă parte, cea mai mare parte a informațiilor despre proprietățile neuroprotectoare ale melatoninei derivate din studii experimentale (vezi pentru [ 177 , 178 ]) face ca este foarte de dorit să se utilizeze doze farmacologice la pacienții cu MCI cu scopul de a opri sau încetini progresia bolii. Deși glanda pineală secretă melatonina care circulă în sânge și LCR [ 179 ], date recente susțin ipoteza că, la ovine și probabil la oameni, numai melatonina LCR, și nu melatonina din sânge, poate furniza cea mai mare parte a melatoninei cerebrale. țesut în concentrații mari [ 180]. Au fost demonstrati gradienti de concentratie semnificativi orientati de la ventricul (aproape de LCR) catre tesutul cerebral, cu concentratii variind cu un factor de la 1 la 125. Prin urmare, pe lângă susținerea rolului LCR în disponibilitatea fiziologică a melatoninei [ 179 ], aceste rezultate implică faptul că trebuie administrate cantități mari, farmacologice de melatonină pentru a avea acces la creier.Mecanismele care explică efectul terapeutic al melatoninei la pacienții cu MCI rămân de definit. Tratamentul cu melatonina promovează în principal somnul cu unde lente la vârstnici [
181 ] și poate fi benefic în MCI prin creșterea fazelor de restaurare ale somnului, inclusiv secreția crescută de GH și neurotrofine.După cum sa subliniat mai sus, melatonina acționează la diferite niveluri relevante pentru dezvoltarea și manifestarea AD. Efectele antioxidante, mitocondriale și antiamiloidogenice pot interfera eventual cu debutul bolii. Prin urmare, momentul în care începe tratamentul cu melatonină poate fi decisiv pentru răspunsul final [ 75 ].Un aspect important care trebuie luat în considerare este doza de melatonină folosită, care poate fi inutil de scăzută atunci când se iau în considerare afinitățile de legare, timpul de înjumătățire și potențele relative ale diferiților agoniști de melatonină de pe piață. Pe lângă faptul că sunt în general mai puternici decât molecula nativă, analogii melatoninei sunt folosiți în cantități considerabil mai mari [ 182 ]. Dozele autorizate de ramelteon agonist al receptorului melatoninei variază de la 8 la 32 mg/zi, în timp ce agomelatina a fost autorizată pentru tratamentul tulburării depresive majore în doze de 25-50 mg/zi. În studiile clinice care implică subiecți umani sănătoși, tasimelteon (Vanda Pharmaceuticals, Washington, DC, SUA), un alt agonist al receptorului de melatonină aprobat recent de FDA este administrat în doze de 20 până la 100 mg/zi [ 183 .] în timp ce farmacocinetica, farmacodinamia și siguranța agonistului receptorului de melatonină TIK-301 (Tikvah Pharmaceuticals, Atlanta, GA, SUA) au fost examinate într-un studiu controlat cu placebo utilizând 20 până la 100 mg/zi [184 ] . Prin urmare, studiile MCI cu doze de melatonină în intervalul 100-300 mg/zi sunt justificate în continuare.O terapie combinată a melatoninei cu un agonist al receptorului melatoninei ar putea fi teoretic benefică pentru pacienții cu AD, mai ales dacă agonistul receptorului melatoninei are un profil farmacocinetic mai bun decât melatonina. Cu toate acestea, agoniştii receptorilor melatoninei au demonstrat beneficii limitate în modelele murine de AD [ 185 ].Deoarece melatonina este eliminată foarte rapid din fluxul sanguin, strategiile cum ar fi inhibarea degradării melatoninei și eliminarea din organism ar putea fi un supliment util la tratamentul cu melatonina exogenă. Într-adevăr, nivelurile plasmatice farmacologice ale melatoninei ar putea fi afectate ca urmare a expunerii concomitente la substanțe chimice care modulează expresia CYP1A2. De exemplu, fluvoxamina, un inhibitor puternic al CYP1A2 și, într-o măsură mai mică, al CYP2C19, a crescut nivelurile plasmatice de melatonina [
187 ]. De asemenea, consumul concomitent de cofeină al cărei metabolism este catalizat în principal de CYP1A2, a dublat nivelurile plasmatice și a crescut biodisponibilitatea melatoninei [
188] .]. Un alt candidat este 5-metoxipsoralen, un medicament utilizat pentru tratamentul psoriazisului, care crește nivelurile plasmatice ale melatoninei endogene și exogene [
191 ].Trebuie subliniat faptul că melatonina are un profil ridicat de siguranță, este de obicei remarcabil de bine tolerată și, în unele studii, a fost administrată la pacienți în doze foarte mari [16 , 192 , 193, 194 195 ] . Melatonina (300 mg/zi) timp de până la trei ani a scăzut stresul oxidativ la pacienții cu scleroză laterală amiotrofică [ 192 ].La copiii cu distrofie musculară, 70 mg/zi de melatonină au redus citokinele și peroxidarea lipidelor [ 193 ]. Doze de 80 mg melatonină pe oră timp de 4 ore au fost administrate bărbaților sănătoși, fără alte reacții adverse în afară de somnolență [ 16 ]. La femeile sănătoase, cărora li sa administrat 300 mg melatonină/zi timp de patru luni, nu au existat efecte secundare [ 194 ]. Un studiu clinic randomizat, controlat, dublu-orb, pe 50 de pacienți trimiși pentru o intervenție chirurgicală hepatică, a indicat că o singură doză enterală preoperatorie de 50 mg/kg melatonină (adică, echivalentul a 3 g pentru un adult de 60 kg) a fost sigură și bine tolerată.
195 ].Un alt rezultat al studiului raportat recent [ 175 ] a fost că atunci când se folosește melatonina, sunt necesare mult mai puține benzodiazepine pentru a trata tulburările de somn în MCI. Deoarece, după cum s-a menționat mai sus, melatonina și benzodiazepinele au împărtășit unele proprietăți neurochimice (
adică , interacțiunea cu mecanismele mediate de GABA în creier [ 196 ]) și comportamentale (de exemplu, o activitate anxiolitică similară dependentă de zi [ 46 ]), terapia cu melatonină a fost postulată să să fie un instrument eficient pentru a scădea doza de benzodiazepine necesară la pacienți [
199]. O analiză retrospectivă recentă a unei baze de date germane de prescripții a identificat 512 pacienți care au inițiat tratament cu melatonină cu eliberare prelungită (2 mg) pe o perioadă de 10 luni [200
] . Din 112 pacienți din acest grup care au utilizat anterior benzodiazepine, 31% au întrerupt tratamentul cu benzodiazepine la trei luni după începerea tratamentului cu melatonină cu eliberare prelungită. Rata de întrerupere a fost mai mare la pacienții care au primit două sau trei rețete de melatonină [
200 ]. Utilizarea prelungită a benzodiazepinelor și agoniștilor receptorilor de benzodiazepină (
Z-droguri) este legată de simptome severe de sevraj și de dependență potențială, care a devenit o problemă de sănătate publică, ducând la mai multe campanii de scădere a consumului acestor medicamente. Un studiu farmacoepidemiologic recent a concluzionat că aceste campanii au eșuat în general atunci când nu au fost asociate cu disponibilitatea și piața melatoninei [
În concluzie, întrebarea dacă melatonina are o valoare terapeutică în prevenirea sau tratarea MCI, care afectează inițierea bolii sau progresia neuropatologiei și mecanismele implicate, merită să fie analizată în continuare. Sunt necesare studii multicentrice dublu-orb pentru a explora și investiga potențialul și utilitatea melatoninei ca medicament antidemență în stadiul incipient al bolii.Deoarece melatonina prezintă atât proprietăți hipnotice, cât și cronobiotice, a fost utilizată terapeutic pentru tratamentul insomniei legate de vârstă, precum și al altor insomnii primare și secundare [
203 ]. Un consens al Asociației Britanice pentru Psihofarmacologie privind tratamentul bazat pe dovezi al insomniei, parasomniei și tulburărilor de somn de ritm circadian a concluzionat că melatonina în doză de 2 mg este tratamentul de primă alegere atunci când un hipnotic este indicat la pacienții cu vârsta peste 55 de ani [
204 ] .După cum arată afinitățile de legare, timpul de înjumătățire și potențele relative ale diferiților agoniști ai melatoninei de pe piață, este clar că studiile care utilizează aceste doze mici de melatonină nu sunt adecvate pentru a oferi o comparație adecvată cu efectul compușilor menționați mai sus, care, în plus pentru a fi în general mai puternice decât molecula nativă sunt folosite în cantități considerabil mai mari [
205 ]. Prin urmare, sunt necesare studii suplimentare care folosesc doze mai mari de melatonină pentru a clarifica potențialele sale implicații neuroprotectoare terapeutice la om. Din studiile pe animale, este clar că o serie de efecte preventive ale melatoninei, cum ar fi cele în tulburările neurodegenerative, au nevoie de doze mari de melatonină pentru a deveni evidente [
177] .]. Dacă ne așteptăm ca melatonina să fie un neuroprotector eficient, este probabil ca dozele mici de melatonină folosite până acum să nu fie foarte benefice.
Tabel 2. Studii care includ tratamentul pacienților cu deficiență cognitivă ușoară (MCI) cu melatonină.
Abrevieri
Ach
acetilcolina
Durere
acetilcolinesteraza
ANUNȚ
Boala Alzheimer
ADI
Alzheimer Disease International
AFMK
N1 – acetil- N2 – formil-5-metoxicinuramină
Akt
protein kinaza identificată în virusul AKT
AMK
N1 – acetil-5-metoxicinuramină
APP
proteine precursoare de amiloid
Ap
β-amiloid agregat
Bcl-2
Proteina proto-oncogenă a limfomului cu celule B
Conversație
colin acetiltransferaza
Cox
ciclooxigenaza
LCR
fluid cerebrospinal
CYP1A1
citocromul P450 1A1
CYP1A2
citocromul P450 1A2
CYP2C19
citocromul P450 2C19
CYP1B1
citocromul P450 1B1
GABA
acid y-aminobutiric
GPR50
G-protein receptor 50 ortholog
GLUT-1
transportor de glucoză-1
GLUT-3
transportor de glucoză-3
GPx
glutation peroxidază
GRd
glutation reductază
GSH
glutation redus
GSK-3
glicogen sintază kinaza-3
IDE
enzimă degradatoare de insulină
iNOS
sintaza de oxid nitric inductibil
IGF-1
Factorul de creștere asemănător insulinei 1
HARTĂ
proteine asociate microtubulilor
MCI
deficit cognitiv minor
mPTP
porul de tranziție al permeabilității mitocondriale
ARNm
acid ribonucleic mesager
MT 1
receptor de melatonina 1
MT 2
receptor de melatonina 2
NFkB
factorul nuclear κB
NMDA
N -metil- d -aspartat
nNOS
oxid nitric sintază neuronală
NU
oxid de azot
NSAD
medicamente antiinflamatoare nesteroidiene
PI3-K
fosfoinozitid 3-kinaza
PK
protein kinaza
REM
miscare rapida a ochilor
RNS
specii reactive de azot
ROR
receptorul orfan legat de receptorul acidului retinoic
ROS
specii reactive de oxigen
RZR
receptorul retinoid Z
SCN
nuclei suprachiasmatici
GAZON
superoxid dismutaza
Mulțumiri
Studiile în laboratoarele autorilor au fost susținute de granturi de la Agencia Nacional de Promoción Científica y Tecnológica, Argentina și Universitatea din Buenos Aires. DPC este un laureat pentru cariera de cercetare de la Consiliul de Cercetare din Argentina (CONICET) și profesor emerit, Universitatea din Buenos Aires. DEV și LIB sunt premiați pentru cariera de cercetare de la CONICET.
Contribuții ale autorului
DPC a scris schița generală a recenziei. Toți ceilalți autori au citit și au fost de acord cu versiunea.
Conflicte de interes
Autorii nu declară niciun conflict de interese.
Referințe
Reiter, RJ; Garcia, JJ; Pie, J. Toxicitate oxidativă în modele de neurodegenerare: răspunsuri la melatonina. Restaurare. Neurol. Neurosci. 1998 , 12 , 135–142. [ Google Scholar ] [ PubMed ]
Johnson, EJ; Vishwanathan, R.; Johnson, MA; Hausman, DB; Davey, A.; Scott, TM; Verde, RC; Miller, LS; Gearing, M.; Woodard, J.; et al. Relația dintre carotenoizii serici și cerebrali, concentrațiile de α-tocoferol și retinol și performanța cognitivă la cel mai vechi vechi din studiul Georgia Centenarian. J. Aging Res. 2013 , 2013 . [ Google Scholar ] [ CrossRef ]
Bubenik, GA; Konturek, SJ Melatonina și îmbătrânirea: perspective pentru tratamentul uman. J. Physiol Pharmacol. 2011 , 62 , 13–19. [ Google Scholar ] [ PubMed ]
Claustrat, B.; Brun, J.; Chazot, G. Fiziologia de bază și patofiziologia melatoninei. Sleep Med. Rev. 2005 , 9 , 11–24. [ Google Scholar ] [ CrossRef ]
Venegas, C.; Garcia, JA; Escames, G.; Ortiz, F.; Lopez, A.; Doerrier, C.; Garcia-Corzo, L.; Lopez, LC; Reiter, RJ; Acuña-Castroviejo, D. Melatonina extrapineală: Analiza distribuției sale subcelulare și a fluctuațiilor zilnice. J. Pineal Res. 2012 , 52 , 217–227. [ Google Scholar ] [ CrossRef ]
Paredes, SD; Korkmaz, A.; Manchester, LC; Tan, DX; Reiter, RJ Phytomelatonin: O revizuire. J. Exp. Bot. 2009 , 60 , 57–69. [ Google Scholar ] [ PubMed ]
Cardinali, DP; Lynch, HJ; Wurtman, RJ Legarea melatoninei de proteinele plasmatice umane și de șobolan. Endocrinologie 1972 , 91 , 1213–1218. [ Google Scholar ] [ CrossRef ]
Ma, X.; Idle, JR; Krausz, KW; Gonzalez, FJ Metabolismul melatoninei de către citocromii umani p450. Drug Metab. Dispos. 2005 , 33 , 489–494. [ Google Scholar ] [ CrossRef ]
Facciola, G.; Hidestrand, M.; von Bahr, C.; Tybring, G. Izoformele citocromului P450 implicate în metabolismul melatoninei în microzomii ficatului uman. EURO. J. Clin. Pharmacol. 2001 , 56 , 881–888. [ Google Scholar ] [ CrossRef ]
Skene, DJ; Papagiannidou, E.; Hashemi, E.; Snelling, J.; Lewis, DF; Fernandez, M.; Ioannides, C. Contribuția CYP1A2 în metabolismul hepatic al melatoninei: Studii cu preparate microzomale izolate și felii de ficat. J. Pineal Res. 2001 , 31 , 333–342. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Young, IM; Leone, RM; Francis, P.; Stovell, P.; Silman, RE Melatonina este metabolizată în N -acetil serotonină și 6-hidroximelatonină la om. J. Clin. Endocrinol. Metab. 1985 , 60 , 114–119. [ Google Scholar ] [ CrossRef ]
Hardeland, R.; Tan, DX; Reiter, RJ Kynuramines, metaboliți ai melatoninei și alți indoli: învierea unei clase aproape uitate de amine biogene. J. Pineal Res. 2009 , 47 , 109–126. [ Google Scholar ] [ CrossRef ]
Tan, DX; Manchester, LC; Terron, deputat; Flores, LJ; Reiter, RJ O moleculă, mulți derivați: o interacțiune fără sfârșit a melatoninei cu speciile reactive de oxigen și azot? J. Pineal Res. 2007 , 42 , 28–42. [ Google Scholar ] [ CrossRef ]
Waldhauser, F.; Waldhauser, M.; Lieberman, HR; Deng, MH; Lynch, HJ; Wurtman, RJ Biodisponibilitatea melatoninei orale la om. Neuroendocrinology 1984 , 39 , 307–313. [ Google Scholar ] [ CrossRef ]
Aldhous, M.; Franey, C.; Wright, J.; Arendt, J. Concentrațiile plasmatice de melatonina la om în urma absorbției orale a diferitelor preparate. Br. J. Clin. Pharmacol. 1985 , 19 , 517–521. [ Google Scholar ] [ CrossRef ]
Fourtillan, JB; Brisson, AM; Gobin, P.; Ingrand, I.; Decourt, JP; Girault, J. Biodisponibilitatea melatoninei la om după administrarea de zi a melatoninei D7 . Biopharm. Eliminarea medicamentelor. 2000 , 21 , 15–22. [ Google Scholar ] [ CrossRef ]
Dubocovich, ML; Delgrange, P.; Krause, DN; Sugden, D.; Cardinali, DP; Olcese, J. Uniunea Internațională de Farmacologie de bază și clinică. LXXV. Nomenclatura, clasificarea și farmacologia receptorilor de melatonină cuplați cu proteina G. Pharmacol. Rev. 2010 , 62 , 343–380. [ Google Scholar ] [ CrossRef ]
Levoye, A.; Dam, J.; Ayoub, MA; Guillaume, JL; Couturier, C.; Delgrange, P.; Jockers, R. Receptorul orfan GPR 50 inhibă în mod specific funcția receptorului de melatonină MT 1 prin heterodimerizare. EMBO J. 2006 , 25 , 3012–3023. [ Google Scholar ] [ CrossRef ]
Wiesenberg, I.; Missbach, M.; Kahlen, JP; Schrader, M.; Carlberg, C. Activarea transcripțională a receptorului nuclear RZR alfa de către melatonina, hormonul glandei pineale și identificarea CGP 52608 ca ligand sintetic. Acizi nucleici Res. 1995 , 23 , 327–333. [ Google Scholar ] [ CrossRef ]
Lardone, PJ; Guerrero, JM; Fernandez-Santos, JM; Rubio, A.; Martin-Lacave, I.; Carrillo-Vico, A. Melatonina sintetizată de limfocitele T ca ligand al receptorului orfan legat de acidul retinoic. J. Pineal Res. 2011 , 51 , 454–462. [ Google Scholar ] [ CrossRef ]
Galano, A.; Tan, DX; Reiter, RJ Melatonina ca un aliat natural împotriva stresului oxidativ: o examinare fizico-chimică. J. Pineal Res. 2011 , 51 , 1–16. [ Google Scholar ] [ CrossRef ]
Antolin, I.; Rodriguez, C.; Sainz, RM; Mayo, JC; Uria, H.; Kotler, ML; Rodriguez-Colunga, MJ; Tolivia, D.; Menendez-Pelaez, A. Neurohormone melatonina previne deteriorarea celulelor: efect asupra expresiei genelor pentru enzimele antioxidante. FASEB J. 1996 , 10 , 882–890. [ Google Scholar ] [ PubMed ]
Pablos, MI; Reiter, RJ; Ortiz, GG; Guerrero, JM; Agapito, MT; Chuang, JI; Sewerynek, E. Ritmurile glutation peroxidazei și glutation reductazei în creierul puiului și inhibarea lor de către lumină. Neurochema. Int. 1998 , 32 , 69–75. [ Google Scholar ] [ CrossRef ]
Rodriguez, C.; Mayo, JC; Sainz, RM; Antolin, I.; Herrera, F.; Martin, V.; Reiter, RJ Reglarea enzimelor antioxidante: un rol semnificativ pentru melatonina. J. Pineal Res. 2004 , 36 , 1–9. [ Google Scholar ] [ CrossRef ]
Jimenez-Ortega, V.; Cano, P.; Cardinali, DP; Esquifino, AI Variația de 24 de ore în expresia genelor a enzimelor căii redox în hipotalamusul de șobolan: Efectul tratamentului cu melatonina. Redox Rep. 2009 , 14 , 132–138. [ Google Scholar ] [ CrossRef ]
Subramanian, P.; Mirunalini, S.; Pandi-Perumal, SR; Trakht, I.; Cardinali, DP Tratamentul cu melatonina imbunatateste statutul antioxidant si scade continutul de lipide din creier si ficat la sobolani. EURO. J. Pharmacol. 2007 , 571 , 116–119. [ Google Scholar ] [ CrossRef ]
Kilanczyk, E.; Bryszewska, M. Efectul melatoninei asupra enzimelor antioxidante în fibroblastele pielii diabetice umane. Celulă. Mol. Biol. Lett. 2003 , 8 , 333–336. [ Google Scholar ] [ PubMed ]
Cardinali, DP; Ritta, MN; Fuentes, AM; Gimeno, MF; Gimeno, AL Eliberarea de prostaglandină E de către hipotalamusul bazal medial de șobolan in vitro . Inhibarea de către melatonină la concentrații submicromolare. EURO. J. Pharmacol. 1980 , 67 , 151–153. [ Google Scholar ] [ CrossRef ]
Deng, WG; Tang, ST; Tseng, HP; Wu, KK Melatonina suprimă macrofagele ciclooxigenazei-2 și expresia inductibilă a oxidului nitric sintazei prin inhibarea acetilării și legarii p52. Blood 2006 , 108 , 518–524. [ Google Scholar ] [ CrossRef ]
Costantino, G.; Cuzzocrea, S.; Mazzon, E.; Caputi, AP Efectele protectoare ale melatoninei în inflamația labei indusă de plasmă activată de zimosan. EURO. J. Pharmacol. 1998 , 363 , 57–63. [ Google Scholar ] [ CrossRef ]
Tan, D.; Reiter, RJ; Chen, LD; Poeggeler, B.; Manchester, LC; Barlow-Walden, LR Atât nivelurile fiziologice, cât și cele farmacologice ale melatoninei reduc formarea aductului ADN indusă de safrolul cancerigen. Carcinogenesis 1994 , 15 , 215–218. [ Google Scholar ] [ CrossRef ]
Leon-Blanco, MM; Guerrero, JM; Reiter, RJ; Pozo, D. Expresia ARN a subunităților de telomerază umană TR și TERT este afectată diferențial de agoniştii receptorului de melatonină din linia celulară tumorală MCF-7. Cancer Lett. 2004 , 216 , 73–80. [ Google Scholar ] [ CrossRef ]
Urata, Y.; Honma, S.; Mergi la S.; Todoroki, S.; Iida, T.; Cho, S.; Honma, K.; Kondo, T. Melatonina induce sintetaza y-glutamilcisteină mediată de proteina activatoare-1 în celulele endoteliale vasculare umane. Radic liber. Biol. Med. 1999 , 27 , 838–847. [ Google Scholar ] [ CrossRef ]
Poliandri, AH; Esquifino, AI; Cano, P.; Jimenez, V.; Lafuente, A.; Cardinali, DP; Duvilanski, BH Efectul protector in vivo al melatoninei asupra modificărilor induse de cadmiu în echilibrul redox și expresia genelor în hipotalamusul de șobolan și pituitara anterioară. J. Pineal Res. 2006 , 41 , 238–246. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Jimenez-Ortega, V.; Cano, P.; Scacchi, PA; Cardinali, DP; Esquifino, AI Perturbarea indusă de cadmiu în expresia de 24 de ore a genelor de ceas și enzime redox în hipotalamusul bazal medial de șobolan. Prevenirea prin melatonină. Față. Neurol. 2011 , 2 . [ Google Scholar ] [ CrossRef ]
Shaikh, AY; Xu, J.; Wu, Y.; El, L.; Hsu, CY Melatonina protejează celulele endoteliale cerebrale bovine de deteriorarea ADN-ului indusă de hiperoxie și de moarte. Neurosci. Lett. 1997 , 229 , 193–197. [ Google Scholar ] [ CrossRef ]
Pablos, MI; Reiter, RJ; Chuang, JI; Ortiz, GG; Guerrero, JM; Sewerynek, E.; Agapito, MT; Melchiorri, D.; Lawrence, R.; Deneke, SM Melatonina administrată acut reduce daunele oxidative în plămâni și creier induse de oxigenul hiperbaric. J. Apl. Physiol. 1997 , 83 , 354–358. [ Google Scholar ] [ PubMed ]
Princ, FG; Juknat, AA; Maxit, AG; Cardalda, C.; Batlle, A. Protecția antioxidantă a melatoninei împotriva daunelor oxidative induse de acidul δ-aminolevulinic în cerebelul șobolanului. J. Pineal Res. 1997 , 23 , 40–46. [ Google Scholar ] [ CrossRef ]
Carneiro, RC; Reiter, RJ Peroxidarea lipidelor indusă de acidul δ-aminolevulinic în rinichi și ficat de șobolan este atenuată de melatonină: un studiu in vitro și in vivo . J. Pineal Res. 1998 , 24 , 131–136. [ Google Scholar ] [ CrossRef ]
Onuki, J.; Almeida, EA; Medeiros, MH; Di, MP Inhibarea deteriorării ADN-ului induse de acidul 5-aminolevulinic de către melatonină, N1 – acetil- N2 – formil-5-metoxicinuramină, quercetină sau resveratrol. J. Pineal Res. 2005 , 38 , 107–115. [ Google Scholar ] [ CrossRef ]
Erol, FS; Topsakal, C.; Ozveren, MF; Kaplan, M.; Ilhan, N.; Ozercan, IH; Yildiz, OG Efectele protectoare ale melatoninei și vitaminei E în leziunile cerebrale datorate radiațiilor gamma: un studiu experimental. Neurochirurgie. Rev. 2004 , 27 , 65–69. [ Google Scholar ] [ CrossRef ]
Shirazi, A.; Haddadi, GH; Asadi-Amoli, F.; Sakhaee, S.; Ghazi-Khansari, M.; Avand, A. Efectul radioprotector al melatoninei în reducerea stresului oxidativ în lentilele de șobolan. Cell J. 2011 , 13 , 79–82. [ Google Scholar ] [ PubMed ]
Taysi, S.; Memisogullari, R.; Koc, M.; Yazici, AT; Aslankurt, M.; Gumustekin, K.; Al, B.; Ozabacigil, F.; Yilmaz, A.; Tahsin, OH Melatonina reduce stresul oxidativ din lentila de șobolan din cauza leziunii oxidative induse de radiații. Int. J. Radiat. Biol. 2008 , 84 , 803–808. [ Google Scholar ] [ CrossRef ]
Lee, EJ; Wu, TS; Lee, MEA; Chen, TY; Tsai, YY; Chuang, JI; Chang, GL Tratamentul întârziat cu melatonină îmbunătățește recuperarea electrofiziologică după ischemia cerebrală focală tranzitorie la șobolani. J. Pineal Res. 2004 , 36 , 33–42. [ Google Scholar ] [ CrossRef ]
Tai, SH; Hung, YC; Lee, EJ; Lee, AC; Chen, TY; Shen, CC; Chen, HY; Lee, MEA; Huang, SY; Wu, TS Melatonina protejează împotriva ischemiei cerebrale focale tranzitorii atât la femelele active din punct de vedere reproductiv, cât și la femelele cu deficit de estrogen: impactul estrogenului circulant asupra răspunsului său hormetic la doză. J. Pineal Res. 2011 , 50 , 292–303. [ Google Scholar ] [ CrossRef ]
Beni, SM; Kohen, R.; Reiter, RJ; Tan, DX; Shohami, E. Neuroprotecția indusă de melatonină după leziuni ale capului închis este asociată cu creșterea antioxidanților din creier și activarea atenuată în fază târzie a NF-kB și AP-1. FASEB J. 2004 , 18 , 149–151. [ Google Scholar ] [ PubMed ]
Tsai, MC; Chen, WJ; Tsai, MS; Ching, CH; Chuang, JI Melatonina atenuează insulta oxidativă indusă de contuzia cerebrală, inactivarea traductorilor de semnal și a activatorilor transcripției 1 și reglarea în sus a supresoarelor de semnalizare a citokinei-3 la șobolani. J. Pineal Res. 2011 , 51 , 233–245. [ Google Scholar ] [ CrossRef ]
Kabadi, SV; Maher, TJ Posttratamentul cu uridină și melatonină în urma unei leziuni cerebrale traumatice reduce edemul în diferite regiuni ale creierului la șobolani. Ann. NY Acad. Sci. 2010 , 1199 , 105–113. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Reiter, RJ; Manchester, LC; Tan, DX Neurotoxine: Mecanisme de radicali liberi si protectie cu melatonina. Curr. Neurofarmacol. 2010 , 8 , 194–210. [ Google Scholar ] [ CrossRef ]
Golombek, DA; Pevet, P.; Cardinali, DP Efectele melatoninei asupra comportamentului: posibilă mediere de către sistemul central GABAergic. Neurosci. Comportament biologic. Rev. 1996 , 20 , 403–412. [ Google Scholar ] [ CrossRef ]
Caumo, W.; Levandovski, R.; Hidalgo, MP Efectul anxiolitic preoperator al melatoninei și clonidinei asupra durerii postoperatorii și a consumului de morfină la pacienții supuși histerectomiei abdominale: un studiu dublu-orb, randomizat, controlat cu placebo. J. Pain 2009 , 10 , 100–108. [ Google Scholar ] [ CrossRef ]
Louzada, PR; Paula Lima, AC; Mendonca-Silva, DL; Noel, F.; de Mello, FG; Ferreira, ST Taurina previne neurotoxicitatea agoniștilor receptorilor beta-amiloid și glutamat: activarea receptorilor GABA și posibile implicații pentru boala Alzheimer și alte tulburări neurologice. FASEB J. 2004 , 18 , 511–518. [ Google Scholar ] [ CrossRef ]
Giusti, P.; Lipartiti, M.; Franceschini, D.; Schiavo, N.; Floreani, M.; Manev, H. Neuroprotecția prin melatonina de la excitotoxicitatea indusă de kainat la șobolani. FASEB J. 1996 , 10 , 891–896. [ Google Scholar ] [ PubMed ]
Manev, H.; Uz, T.; Kharlamov, A.; Cagnoli, CM; Franceschini, D.; Giusti, P. Protecția in vivo împotriva apoptozei induse de kainat de către hormonul pineal melatonina: Efectul melatoninei exogene și al ritmului circadian. Restaurare. Neurol. Neurosci. 1996 , 9 , 251–256. [ Google Scholar ] [ PubMed ]
Cho, S.; Joh, TH; Baik, HH; Dibinis, C.; Volpe, BT Administrarea melatoninei protejează neuronii hipocampali CA1 după ischemia tranzitorie a creierului anterior la șobolani. Brain Res. 1997 , 755 , 335–338. [ Google Scholar ] [ CrossRef ]
Kilic, E.; Ozdemir, YG; Bolay, H.; Kelestimur, H.; Dalkara, T. Pinealectomia agravează iar administrarea melatoninei atenuează afectarea creierului în ischemia focală. J. Cereb. Fluxul de sânge Metab. 1999 , 19 , 511–516. [ Google Scholar ] [ PubMed ]
Furio, AM; Fontao, R.; Falco, N.; Ruiz, JI; Caccuri, RL; Cardinali, DP Efectul neuroprotector al melatoninei asupra toxicității glucocorticoizilor în hipocampul de șobolan. Deschide Physiol. J. 2008 , 1 , 23–27. [ Google Scholar ] [ CrossRef ]
Dodd, S.; Maes, M.; Anderson, G.; Dean, OM; Moylan, S.; Berk, M. Putative neuroprotective agents in neuropsihiatric disorders. Prog. Neuropsihofarmacol. Biol. Psihiatrie 2013 , 42 , 135–145. [ Google Scholar ] [ CrossRef ]
Jiao, S.; Wu, MM; Hu, CL; Zhang, ZH; Mei, YA 2-iodomelatonina agonistul receptorului de melatonina previne apoptoza neuronilor granulelor cerebeloase prin inhibarea curentului K + . J. Pineal Res. 2004 , 36 , 109–116. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Koh, PO Melatonina previne reglarea în jos a fosfoproteinei astrocitare PEA-15 în leziunile cerebrale ischemice. J. Pineal Res. 2011 , 51 , 381–386. [ Google Scholar ] [ CrossRef ]
Radogna, F.; Diederich, M.; Ghibelli, L. Melatonin: O moleculă pleiotropă care reglează inflamația. Biochim. Pharmacol. 2010 , 80 , 1844–1852. [ Google Scholar ] [ CrossRef ]
Peng, TI; Hsiao, CW; Reiter, RJ; Tanaka, M.; Lai, YK; Jou, MJ mtDNA T8993G Inhibarea complexului mitocondrial V indusă de mutația MJ mărește modificările dependente de cardiolipină ale dinamicii mitocondriale în timpul insultelor oxidative, Ca 2+ și lipidice la cibrizii NARP: o țintă potențială terapeutică pentru melatonină. J. Pineal Res. 2012 , 52 , 93–106. [ Google Scholar ] [ CrossRef ]
Jou, MJ Melatonina păstrează tranziția tranzitorie a permeabilității mitocondriale pentru protecție în timpul stresului mitocondrial de Ca 2+ în astrocite. J. Pineal Res. 2011 , 50 , 427–435. [ Google Scholar ] [ CrossRef ]
Andrabi, SA; Sayeed, eu; Siemen, D.; Wolf, G.; Horn, TF Inhibarea directă a porului de tranziție a permeabilității mitocondriale: un posibil mecanism responsabil pentru efectele anti-apoptotice ale melatoninei. FASEB J. 2004 , 18 , 869–871. [ Google Scholar ] [ PubMed ]
Hardeland, R. Melatonina și teoriile îmbătrânirii: o evaluare critică a rolului melatoninei în mecanismele antiîmbătrânire. J. Pineal Res. 2013 , 55 , 325–356. [ Google Scholar ] [ PubMed ]
Selkoe, DJ Biologia celulară a plierii greșite a proteinelor: exemplele bolilor Alzheimer și Parkinson. Nat. Cell Biol. 2004 , 6 , 1054–1061. [ Google Scholar ] [ CrossRef ]
Lahiri, DK; Ghosh, C. Interacțiuni între melatonină, specii reactive de oxigen și oxid nitric. Ann. NY Acad. Sci. 1999 , 893 , 325–330. [ Google Scholar ] [ CrossRef ]
Matsubara, E.; Bryant-Thomas, T.; Pacheco, QJ; Henry, TL; Poeggeler, B.; Herbert, D.; Cruz-Sanchez, F.; Chyan, YJ; Smith, MA; Perry, G.; et al. Melatonina crește supraviețuirea și inhibă patologia oxidativă și amiloidă într-un model transgenic al bolii Alzheimer. J. Neurochem. 2003 , 85 , 1101–1108. [ Google Scholar ] [ CrossRef ]
Lahiri, DK; Chen, D.; Ge, YW; Bondy, SC; Sharman, EH Suplimentarea alimentară cu melatonină reduce nivelul de β-peptide amiloide în cortexul cerebral murin. J. Pineal Res. 2004 , 36 , 224–231. [ Google Scholar ] [ CrossRef ]
Song, W.; Lahiri, DK Melatonina modifică metabolismul proteinei precursoare beta-amiloid din linia celulară neuroendocrină PC12. J. Mol. Neurosci. 1997 , 9 , 75–92. [ Google Scholar ] [ CrossRef ]
Olivieri, G.; Hess, C.; Savaskan, E.; Ly, C.; Meier, F.; Baysang, G.; Brockhaus, M.; Muller-Spahn, F. Melatonina protejează celulele neuroblastomului SHSY5Y de stresul oxidativ indus de cobalt, neurotoxicitatea și secreția crescută de β-amiloid. J. Pineal Res. 2001 , 31 , 320–325. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Quinn, J.; Kulhanek, D.; Nowlin, J.; Jones, R.; Pratico, D.; Rokach, J.; Stackman, R. Terapia cu melatonina cronica nu reuseste sa modifice sarcina de amiloid sau deteriorarea oxidativa la soarecii vechi Tg2576: Implicatii pentru studiile clinice. Brain Res. 2005 , 1037 , 209–213. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Hsiao, K.; Chapman, P.; Nilsen, S.; Eckman, C.; Harigaya, Y.; Younkin, S.; Yang, F.; Cole, G. Deficiențe de memorie corelative, creșterea Apei și plăci de amiloid la șoarecii transgenici. Science 1996 , 274 , 99–102. [ Google Scholar ] [ CrossRef ]
Donnelly, PS; Caragounis, A.; Du, T.; Laughton, KM; Volitakis, I.; Cherny, RA; Sharples, RA; Hill, AF; Li, QX; Masters, CL; et al. Eliberarea intracelulară selectivă a ionilor de cupru și zinc din complecșii bis (thiosemicarbazonato) reduce nivelurile peptidei amiloid-β ale bolii Alzheimer. J. Biol. Chim. 2008 , 283 , 4568–4577. [ Google Scholar ] [ CrossRef ]
Farris, W.; Mansourian, S.; Chang, Y.; Lindsley, L.; Eckman, EA; Frosch, deputat; Eckman, CB; Tanzi, RE; Selkoe, DJ; Guenette, S. Enzima de degradare a insulinei reglează nivelurile de insulină, β-proteinei amiloid și domeniul intracelular al proteinei precursoare de β-amiloid in vivo . Proc. Natl. Acad. Sci. SUA 2003 , 100 , 4162–4167. [ Google Scholar ] [ CrossRef ]
Poeggeler, B.; Miravalle, L.; Zagorski, MG; Wisniewski, T.; Chyan, YJ; Zhang, Y.; Shao, H.; Bryant-Thomas, T.; Vidal, R.; Frangione, B.; et al. Melatonina inversează activitatea profibrilogenă a apolipoproteinei E4 asupra peptidei amiloid Aβ Alzheimer. Biochimie 2001 , 40 , 14995–15001. [ Google Scholar ] [ CrossRef ]
Pappolla, M.; Bozner, P.; Soto, C.; Shao, H.; Robakis, NK; Zagorski, M.; Frangione, B.; Ghiso, J. Inhibarea β-fibrilogenezei Alzheimer de către melatonina. J. Biol. Chim. 1998 , 273 , 7185–7188. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Feng, Z.; Chang, Y.; Cheng, Y.; Zhang, BL; Qu, ZW; Qin, C.; Zhang, JT Melatonina atenuează deficitele comportamentale asociate cu apoptoza și disfuncția sistemului colinergic în modelul de șoarece transgenic APP 695 al bolii Alzheimer. J. Pineal Res. 2004 , 37 , 129–136. [ Google Scholar ] [ CrossRef ]
Zatta, P.; Tognon, G.; Carampin, P. Melatonina previne formarea radicalilor liberi datorită interacțiunii dintre peptidele β-amiloide și ionii metalici [Al III , Zn II , Cu II , Mn II , Fe II ]. J. Pineal Res. 2003 , 35 , 98–103. [ Google Scholar ] [ CrossRef ]
Furio, AM; Cutrera, RA; Castillo Thea, V.; Perez, LS; Riccio, P.; Caccuri, RL; Brusco, LL; Cardinali, DP Efectul melatoninei asupra modificărilor ritmului activității locomotorii la hamsterii sirieni injectați cu peptida β-amiloid 25-35 în nucleele suprachiasmatice. Celulă. Mol. Neurobiol. 2002 , 22 , 699–709. [ Google Scholar ] [ CrossRef ]
Shen, YX; Xu, SY; Wei, W.; Soare, XX; Yang, J.; Liu, LH; Dong, C. Melatonina reduce modificările de memorie și daunele oxidative neuronale la șoarecii tratați cu d -galactoză. J. Pineal Res. 2002 , 32 , 173–178. [ Google Scholar ] [ CrossRef ]
Rosales-Corral, S.; Tan, DX; Reiter, RJ; Valdivia-Velazquez, M.; Martinez-Barboza, G.; Acosta-Martinez, JP; Ortiz, GG Melatonina administrată oral reduce stresul oxidativ și citokinele proinflamatorii induse de peptida amiloid-β în creierul de șobolan: un studiu comparativ, in vivo versus vitamina C și E. J. Pineal Res. 2003 , 35 , 80–84. [ Google Scholar ] [ CrossRef ]
Olcese, JM; Cao, C.; Mori, T.; Mamcarz, MB; Maxwell, A.; Runfeldt, MJ; Wang, L.; Zhang, C.; Lin, X.; Zhang, G.; et al. Protecție împotriva deficitelor cognitive și a markerilor neurodegenerării prin administrarea orală pe termen lung a melatoninei într-un model transgenic al bolii Alzheimer. J. Pineal Res. 2009 , 47 , 82–96. [ Google Scholar ] [ CrossRef ]
Dragicevic, N.; Copes, N.; O’Neal-Moffitt, G.; Jin, J.; Buzzeo, R.; Mamcarz, M.; Tan, J.; Cao, C.; Olcese, JM; Arendash, GW; et al. Tratamentul cu melatonina restabilește funcția mitocondrială la șoarecii Alzheimer: un rol protector mitocondrial al semnalizării receptorilor membranei melatoninei. J. Pineal Res. 2011 , 51 , 75–86. [ Google Scholar ] [ CrossRef ]
Brion, JP; Anderton, BH; Authelet, M.; Dayanandan, R.; Leroy, K.; Lovestone, S.; Octave, JN; Pradier, L.; Touchet, N.; Tremp, G. Încurcăturile neurofibrilare și fosforilarea tau. Biochim. Soc. Symp. 2001 , 81–88. [ Google Scholar ]
Billingsley, ML; Kincaid, RL Fosforilarea și defosforilarea reglementate a proteinei tau: Efecte asupra interacțiunii microtubulilor, a traficului intracelular și a neurodegenerării. Biochim. J. 1997 , 323 , 577–591. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Khatoon, S.; Grundke-Iqbal, I.; Iqbal, K. Nivelurile creierului de proteină tau asociată cu microtubuli sunt crescute în boala Alzheimer: un test radioimuno-slot-blot pentru nanogramele proteinei. J. Neurochem. 1992 , 59 , 750–753. [ Google Scholar ] [ CrossRef ]
Iqbal, K.; Alonso, AC; Chen, S.; Chohan, MO; El-Akkad, E.; Gong, CX; Khatoon, S.; Li, B.; Liu, F.; Rahman, A.; et al. Patologia Tau în boala Alzheimer și alte tauopatii. Biochim. Biophys. Acta 2005 , 1739 , 198–210. [ Google Scholar ] [ CrossRef ]
Deng, YQ; Xu, GG; Duan, P.; Zhang, Q.; Wang, JZ Efectele melatoninei asupra hiperfosforilării tau induse de wortmannin. Acta Pharmacol. Păcat. 2005 , 26 , 519–526. [ Google Scholar ] [ CrossRef ]
Li, SP; Deng, YQ; Wang, XC; Wang, YP; Wang, JZ Melatonina protejează celulele neuroblastomului SH-SY5Y de afectarea neurofilamentului indusă de caliculina A și neurotoxicitatea. J. Pineal Res. 2004 , 36 , 186–191. [ Google Scholar ] [ CrossRef ]
Li, XC; Wang, ZF; Zhang, JX; Wang, Q.; Wang, JZ Efectul melatoninei asupra hiperfosforilării tau induse de calyculin A. EURO. J. Pharmacol. 2005 , 510 , 25–30. [ Google Scholar ] [ CrossRef ]
Xiong, YF; Chen, Q.; Chen, J.; Zhou, J.; Wang, HX Melatonina reduce afectarea transportului axonal și axonopatia indusă de calyculin A. J. Pineal Res. 2011 , 50 , 319–327. [ Google Scholar ] [ CrossRef ]
Benitez-King, G.; Tunez, I.; Bellon, A.; Ortiz, GG; Anton-Tay, F. Melatonina previne modificările citoscheletice și stresul oxidativ indus de acidul okadaic în celulele N1E-115. Exp. Neurol. 2003 , 182 , 151–159. [ Google Scholar ] [ CrossRef ]
Montilla-Lopez, P.; Munoz-Agueda, MC; Feijoo, LM; Munoz-Castaneda, JR; Bujalance-Arenas, I.; Tunez-Finana, I. Comparația melatoninei versus vitamina C asupra stresului oxidativ și activității enzimelor antioxidante în boala Alzheimer indusă de acid okadaic în celulele neuroblastomului. EURO. J. Pharmacol. 2002 , 451 , 237–243. [ Google Scholar ] [ CrossRef ]
Montilla, P.; Feijoo, M.; Munoz, MC; Munoz-Castaneda, JR; Bujalance, I.; Tunez, I. Efectul melatoninei asupra stresului oxidativ în celulele N1E-115 nu este mediat de receptorii MT1. J. Physiol. Biochim. 2003 , 59 , 263–268. [ Google Scholar ] [ CrossRef ]
Liu, SJ; Wang, JZ Fosforilarea tau asemănătoare Alzheimer indusă de wortmannin in vivo și atenuarea acesteia de către melatonină. Acta Pharmacol. Păcat. 2002 , 23 , 183–187. [ Google Scholar ] [ PubMed ]
Wang, XC; Zhang, J.; Yu, X.; Han, L.; Zhou, ZT; Zhang, Y.; Wang, JZ Prevenirea hiperfosforilării tau indusă de izoproterenol de către melatonină la șobolan. Sheng Li Xue Bao 2005 , 57 , 7–12. [ Google Scholar ] [ PubMed ]
Zhu, LQ; Wang, SH; Ling, ZQ; Wang, DL; Wang, JZ Efectul inhibării biosintezei melatoninei asupra reținerii memoriei spațiale și fosforilării tau la șobolan. J. Pineal Res. 2004 , 37 , 71–77. [ Google Scholar ] [ CrossRef ]
Gomez-Ramos, A.; Diaz-Nido, J.; Smith, MA; Perry, G.; Avila, J. Efectul produsului de peroxidare a lipidelor acroleină asupra fosforilării tau în celulele neuronale. J. Neurosci. Res. 2003 , 71 , 863–870. [ Google Scholar ] [ CrossRef ]
Lovell, MA; Xiong, S.; Xie, C.; Davies, P.; Markesbery, WR Inducerea tau hiperfosforilată în culturile de neuroni corticali primari de șobolan mediată de stresul oxidativ și glicogen sintaza kinaza-3. J. Alzheimer Dis. 2004 , 6 , 659–671. [ Google Scholar ] [ PubMed ]
Kenyon, CJ Genetica îmbătrânirii. Natura 2010 , 464 , 504–512. [ Google Scholar ] [ CrossRef ]
Schuster, C.; Williams, LM; Morris, A.; Morgan, PJ; Barrett, P. Receptorul uman de melatonină MT 1 stimulează producția de cAMP în celulele SH-SY5Y ale liniei celulare de neuroblastom uman printr-o cale de transducție a semnalului calciu-calmodulină. J. Neuroendocrinol. 2005 , 17 , 170–178. [ Google Scholar ] [ CrossRef ]
Peschke, E.; Muhlbauer, E.; Musshoff, U.; Csernus, VJ; Chankiewitz, E.; Peschke, D. Influența mediată de receptorul (MT1 ) a melatoninei asupra concentrației de cAMP și secreției de insulină a celulelor de insulinom de șobolan INS-1. J. Pineal Res. 2002 , 33 , 63–71. [ Google Scholar ] [ CrossRef ]
Witt-Enderby, PA; MacKenzie, RS; McKeon, RM; Carroll, EA; Bordt, SL; Melan, MA Inducerea melatoninei a structurilor filamentoase în celulele non-neuronale care este dependentă de expresia receptorului uman de melatonină MT 1 . Cell Motil. Citoschelet. 2000 , 46 , 28–42. [ Google Scholar ] [ CrossRef ]
Rivera-Bermudez, MA; Gerdin, MJ; Earnest, DJ; Dubocovich, ML Reglarea ritmicității bazale în activitatea protein kinazei C de către melatonină în celulele nucleului suprachiasmatic de șobolan imortalizate. Neurosci. Lett. 2003 , 346 , 37–40. [ Google Scholar ] [ CrossRef ]
Benitez-King, G.; Rios, A.; Martinez, A.; Anton-Tay, F. Inhibarea in vitro a activității kinazei II dependente de Ca2 + /calmodulină de către melatonină. Biochim. Biophys. Acta 1996 , 1290 , 191–196. [ Google Scholar ] [ CrossRef ]
Chan, AS; Lai, FP; Lo, RK; Voyno-Yasenetskaya, TA; Stanbridge, EJ; Receptorii Wong, YH Melatonina MT 1 și MT 2 stimulează kinaza N -terminală c-Jun prin proteinele G sensibile și insensibile la toxina pertussis. Semnal celular. 2002 , 14 , 249–257. [ Google Scholar ] [ CrossRef ]
Arends, YM; Duyckaerts, C.; Rozemuller, JM; Eikelenboom, P.; Hauw, JJ Microglia, amiloid și demență în boala alzheimer. Un studiu corelativ. Neurobiol. Îmbătrânire 2000 , 21 , 39–47. [ Google Scholar ] [ PubMed ]
Combadiere, C.; Feumi, C.; Raoul, W.; Keller, N.; Rodero, M.; Pezard, A.; Lavalette, S.; Houssier, M.; Jonet, L.; Picard, E.; et al. Acumularea de celule microglia subretiniene dependentă de CX3CR1 este asociată cu caracteristicile cardinale ale degenerescenței maculare legate de vârstă. J. Clin. Investig. 2007 , 117 , 2920–2928. [ Google Scholar ] [ CrossRef ][ Versiunea verde ]
Streit, WJ; Mrak, RE; Griffin, WS Microglia și neuroinflamație: o perspectivă patologică. J. Neuroinflamm. 2004 , 1 . [ Google Scholar ] [ CrossRef ][ Versiunea verde ]
Shen, Y.; Zhang, G.; Liu, L.; Xu, S. Efectele supresive ale melatoninei asupra activării gliale induse de amiloid-β în hipocampul șobolanului. Arc. Med. Res. 2007 , 38 , 284–290. [ Google Scholar ] [ CrossRef ]
Stuchbury, G.; Munch, G. Inflamația asociată cu Alzheimer, potențialele ținte de droguri și terapiile viitoare. J. Transm. Neural. 2005 , 112 , 429–453. [ Google Scholar ] [ CrossRef ]
Park, SY; Jin, ML; Kim, YH; Kim, Y.; Lee, SJ Efecte antiinflamatorii ale turmeronului aromatic prin blocarea căilor de semnalizare MAPK NF-κB, JNK și p38 în microglia stimulată de beta-amiloid. Int. Imunofarmacol. 2012 , 14 , 13–20. [ Google Scholar ] [ CrossRef ]
Samuel, W.; Masliah, E.; Hill, LR; Unturi, N.; Terry, R. Hipocampal conectivitate și demența Alzheimer: Efectele pierderii sinapselor și frecvența încurcăturii într-un model cu două componente. Neurologie 1994 , 44 , 2081–2088. [ Google Scholar ] [ CrossRef ]
Terry, AV, Jr.; Buccafusco, JJ Ipoteza colinergică a vârstei și a deficitelor cognitive legate de boala Alzheimer: provocări recente și implicațiile lor pentru dezvoltarea de noi medicamente. J. Pharmacol. Exp. Acolo. 2003 , 306 , 821–827. [ Google Scholar ] [ CrossRef ]
Rinne, JO; Laine, M.; Hiltunen, J.; Erkinjuntti, T. Luarea deciziilor semantice în AD probabile timpurii: Un studiu de activare PET. Brain Res. Cogn. Brain Res. 2003 , 18 , 89–96. [ Google Scholar ] [ CrossRef ]
Spencer, JP; Middleton, LJ; Davies, CH Investigarea eficacității inhibitorului de acetilcolinesterază, a donepezilului și a noilor agenți procognitivi pentru a induce oscilații γ în feliile de hipocamp de șobolan. Neurofarmacologie 2010 , 59 , 437–443. [ Google Scholar ] [ CrossRef ]
Guermonprez, L.; Ducrocq, C.; Gaudry-Talarmain, YM Inhibarea sintezei acetilcolinei și nitrarea tirozinei induse de peroxinitrit sunt prevenite în mod diferențial de antioxidanți. Mol. Pharmacol. 2001 , 60 , 838–846. [ Google Scholar ] [ PubMed ]
Feng, Z.; Cheng, Y.; Zhang, JT Efectele pe termen lung ale melatoninei sau 17 β-estradiol asupra îmbunătățirii performanței memoriei spațiale la șobolani adulți ovariectomizați, cu deficiențe cognitive. J. Pineal Res. 2004 , 37 , 198–206. [ Google Scholar ] [ CrossRef ]
Tang, F.; Nag, S.; Shiu, SY; Pang, SF Efectele melatoninei și extractului de Ginkgo biloba asupra pierderii memoriei și activităților colin acetiltransferazei în creierul șobolanilor infuzați intracerebroventricular cu β-amiloid 1-40. Life Sci. 2002 , 71 , 2625–2631. [ Google Scholar ] [ CrossRef ]
Agrawal, R.; Tyagi, E.; Shukla, R.; Nath, C. Un studiu al receptorilor de insulină din creier, activitatea AChE și stresul oxidativ în modelul de șobolan al demenței induse de ICV STZ. Neurofarmacologie 2009 , 56 , 779–787. [ Google Scholar ] [ CrossRef ]
Fernandez-Bachiller, MI; Perez, C.; Campillo, NE; Paez, JA; Gonzalez-Munoz, GC; Usan, P.; Garcia-Palomero, E.; Lopez, MG; Villarroya, M.; Garcia, AG; et al. Hibrizii tacrină-melatonină ca agenți multifuncționali pentru boala Alzheimer, cu proprietăți colinergice, antioxidante și neuroprotectoare. ChemMedChem 2009 , 4 , 828–841. [ Google Scholar ] [ CrossRef ]
Spuch, C.; Antequera, D.; Isabel Fernandez-Bachiller, M.; Isabel Rodriguez-Franco, M.; Carro, E. Un nou hibrid tacrina-melatonina reduce sarcina de amiloid si deficitele comportamentale intr-un model de soarece al bolii Alzheimer. Neurotox. Res. 2010 , 17 , 421–431. [ Google Scholar ] [ CrossRef ]
O’Neill, C.; Kiely, AP; Coakley, MF; Manning, S.; Long-Smith, CM Insulină și semnalizare IGF-1: longevitate, homeostazia proteinelor și boala Alzheimer. Biochim. Soc. Trans. 2012 , 40 , 721–727. [ Google Scholar ] [ CrossRef ]
Hildreth, KL; van Pelt, RE; Schwartz, RS Obezitatea, rezistența la insulină și boala Alzheimer. Obezitate 2012 , 20 , 1549–1557. [ Google Scholar ] [ CrossRef ]
De la Monte, SM Rezistența creierului la insulină și deficiența ca ținte terapeutice în boala Alzheimer. Curr. Alzheimer Res. 2012 , 9 , 35–66. [ Google Scholar ] [ CrossRef ]
Thambisetty, M.; Jeffrey, ME; Yang, A.; Dolan, H.; Marano, C.; Zonderman, AB; Troncoso, JC; Zhou, Y.; Wong, DF; Ferrucci, L.; et al. Intoleranța la glucoză, rezistența la insulină și caracteristicile patologice ale bolii Alzheimer în studiul longitudinal Baltimore asupra îmbătrânirii. JAMA Neurol. 2013 , 70 , 1167–1172. [ Google Scholar ] [ CrossRef ]
Cardinali, DP; Cano, P.; Jimenez-Ortega, V.; Esquifino, AI Melatonina și sindromul metabolic: implicații fiziopatologice și terapeutice. Neuroendocrinologie 2011 , 93 , 133–142. [ Google Scholar ] [ CrossRef ]
Cardinali, DP; Bernasconi, PA; Reynoso, R.; Toso, CF; Scacchi, P. Melatonina poate reduce sindromul metabolic: Studii asupra sindromului metabolic inițial și complet stabilit indus de fructoză la șobolani. Int. J. Mol. Sci. 2013 , 14 , 2502–2514. [ Google Scholar ] [ CrossRef ]
Magri, F.; Locatelli, M.; Balza, G.; Molla, G.; Cuzzoni, G.; Fioravanti, M.; Solerte, SB; Ferrari, E. Modificări ale ritmurilor circadiene endocrine ca markeri ai îmbătrânirii creierului fiziologic și patologic. Chronobiol. Int. 1997 , 14 , 385–396. [ Google Scholar ] [ CrossRef ]
Zhou, JN; Liu, RY; Kamphorst, W.; Hofman, MA; Swaab, DF Modificările neuropatologice precoce ale bolii Alzheimer la persoanele în vârstă sunt însoțite de scăderea nivelului de melatonină în lichidul cefalorahidian. J. Pineal Res. 2003 , 35 , 125–130. [ Google Scholar ] [ CrossRef ]
Skene, DJ; Vivien-Roels, B.; Sparks, DL; Hunsaker, JC; Pevet, P.; Ravid, D.; Swaab, DF Variația zilnică a concentrației de melatonină și 5-metoxitriptofol în glanda pineală umană: Efectul vârstei și al bolii Alzheimer. Brain Res. 1990 , 528 , 170–174. [ Google Scholar ] [ CrossRef ]
Ohashi, Y.; Okamoto, N.; Uchida, K.; Iyo, M.; Mori, N.; Morita, Y. Ritmul zilnic al nivelurilor serice de melatonina si efectul expunerii la lumina la pacientii cu dementa de tip Alzheimer. Biol. Psihiatrie 1999 , 45 , 1646–1652. [ Google Scholar ] [ CrossRef ]
Liu, RY; Zhou, JN; van Heerikhuize, J.; Hofman, MA; Swaab, DF Scăderea nivelului de melatonina în lichidul cefalorahidian postmortem în raport cu îmbătrânirea, boala Alzheimer și genotipul apolipoproteinei E-epsilon4/4. J. Clin. Endocrinol. Metab. 1999 , 84 , 323–327. [ Google Scholar ] [ PubMed ]
Mishima, K.; Tozawa, T.; Satoh, K.; Matsumoto, Y.; Hishikawa, Y.; Okawa, M. Tulburări ale ritmului de secreție a melatoninei la pacienții cu demență senilă de tip Alzheimer cu tulburări de somn-vegie. Biol. Psihiatrie 1999 , 45 , 417–421. [ Google Scholar ] [ CrossRef ]
Skene, DJ; Swaab, DF Ritmicitatea melatoninei: Efectul vârstei și al bolii Alzheimer. Exp. Gerontol. 2003 , 38 , 199–206. [ Google Scholar ] [ CrossRef ]
Wu, YH; Feenstra, MG; Zhou, JN; Liu, RY; Torano, JS; van Kan, HJ; Fischer, DF; Ravid, R.; Swaab, DF Modificari moleculare care stau la baza nivelurilor reduse de melatonina pineala in boala Alzheimer: modificari in stadiile preclinice si clinice. J. Clin. Endocrinol. Metab. 2003 , 88 , 5898–5906. [ Google Scholar ] [ CrossRef ]
Savaskan, E.; Ayoub, MA; Ravid, R.; Angeloni, D.; Fraschini, F.; Meier, F.; Eckert, A.; Muller-Spahn, F.; Jockers, R. Expresia redusă a receptorului melatoninei MT 2 din hipocamp în boala Alzheimer. J. Pineal Res. 2005 , 38 , 10–16. [ Google Scholar ] [ CrossRef ]
Savaskan, E.; Olivieri, G.; Meier, F.; Brydon, L.; Jockers, R.; Ravid, R.; Wirz-Justice, A.; Muller-Spahn, F. Imunoreactivitate crescută a receptorului de melatonină 1a în hipocampul pacienților cu boala Alzheimer. J. Pineal Res. 2002 , 32 , 59–62. [ Google Scholar ] [ CrossRef ]
Weldemichael, DA; Grossberg, GT Tulburări de ritm circadian la pacienții cu boala Alzheimer: o revizuire. Int. J. Alzheimer Dis. 2010 , 2010 . [ Google Scholar ] [ CrossRef ]
Klaffke, S.; Staedt, J. Sundowning și tulburări de ritm circadian în demență. Acta Neurol. Belg. 2006 , 106 , 168–175. [ Google Scholar ] [ PubMed ]
Cardinali, DP; Furio, AM; Brusco, LI Aspecte clinice ale intervenției melatoninei în progresia bolii Alzheimer. Curr. Neurofarmacol. 2010 , 8 , 218–227. [ Google Scholar ] [ CrossRef ]
De Jonghe, A.; Korevaar, JC; van Munster, BC; de Rooij, SE Eficacitatea tratamentului cu melatonină asupra tulburărilor de ritm circadian în demență. Există implicații pentru delir? O revizuire sistematică. Int. J. Geriatr. Psihiatrie 2010 , 25 , 1201–1208. [ Google Scholar ] [ CrossRef ]
Pappolla, MA; Chyan, YJ; Poeggeler, B.; Frangione, B.; Wilson, G.; Ghiso, J.; Reiter, RJ O evaluare a proprietăților antioxidante și antiamiloidogene ale melatoninei: Implicații pentru boala Alzheimer. J. Transm. Neural. 2000 , 107 , 203–231. [ Google Scholar ] [ CrossRef ]
Wu, YH; Swaab, DF Glanda pineală umană și melatonina în îmbătrânire și boala Alzheimer. J. Pineal Res. 2005 , 38 , 145–152. [ Google Scholar ] [ CrossRef ]
Wu, YH; Zhou, JN; van Heerikhuize, J.; Jockers, R.; Swaab, DF Scăderea expresiei receptorului de melatonină MT 1 în nucleul suprachiasmatic în îmbătrânire și boala Alzheimer. Neurobiol. Îmbătrânire 2007 , 28 , 1239–1247. [ Google Scholar ] [ CrossRef ]
Fainstein, I.; Bonetto, A.; Brusco, LI; Cardinali, DP Efectele melatoninei la pacienții vârstnici cu tulburări de somn. Un studiu pilot. Curr. Acolo. Res. 1997 , 58 , 990–1000. [ Google Scholar ] [ CrossRef ]
Brusco, LI; Marquez, M.; Cardinali, DP Tratamentul cu melatonina stabilizeaza simptomele cronobiologice si cognitive in boala Alzheimer. Neuro Endocrinol. Lett. 1998 , 19 , 111–115. [ Google Scholar ]
Brusco, LI; Marquez, M.; Cardinali, DP Gemeni monozigoți cu boala Alzheimer tratați cu melatonină: Raport de caz. J. Pineal Res. 1998 , 25 , 260–263. [ Google Scholar ] [ CrossRef ]
Cohen-Mansfield, J.; Garfinkel, D.; Lipson, S. Melatonina pentru tratamentul apusului soarelui la persoanele în vârstă cu demență – Un studiu preliminar. Arc. Gerontol. Geriatr. 2000 , 31 , 65–76. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Mishima, K.; Okawa, M.; Hozumi, S.; Hishikawa, Y. Administrarea suplimentară a luminii strălucitoare artificiale și a melatoninei ca tratament puternic pentru activitatea de repaus circadiană dezorganizată și sistemele autonome și neuroendocrine disfuncționale la persoanele vârstnice dementate instituționalizate. Chronobiol. Int. 2000 , 17 , 419–432. [ Google Scholar ] [ CrossRef ]
Serfaty, M.; Kennell-Webb, S.; Warner, J.; Blizard, R.; Raven, P. Studiu dublu orb, randomizat, controlat cu placebo, de melatonină în doză mică pentru tulburările de somn în demență. Int. J. Geriatr. Psihiatrie 2002 , 17 , 1120–1127. [ Google Scholar ] [ CrossRef ]
Cardinali, DP; Brusco, LI; Liberczuk, C.; Furio, AM Utilizarea melatoninei în boala Alzheimer. Neuro Endocrinol. Lett. 2002 , 23 , 20–23. [ Google Scholar ] [ PubMed ]
Singer, C.; Tractenberg, RE; Kaye, J.; Schafer, K.; Gamst, A.; Grundman, M.; Thomas, R.; Thal, LJ Un studiu multicentric, controlat cu placebo, al melatoninei pentru tulburările de somn în boala Alzheimer. Sleep 2003 , 26 , 893–901. [ Google Scholar ] [ PubMed ]
Asayama, K.; Yamadera, H.; Ito, T.; Suzuki, H.; Kudo, Y.; Endo, S. Studiu dublu-orb al efectelor melatoninei asupra ritmului somn-veghe, funcțiilor cognitive și non-cognitive în demența de tip Alzheimer. J. Nippon Med. Sch. 2003 , 70 , 334–341. [ Google Scholar ] [ CrossRef ]
Mahlberg, R.; Kunz, D.; Sutej, I.; Kuhl, KP; Hellweg, R. Tratamentul cu melatonina a tulburarilor de ritm zi-noapte si apusul soarelui in boala Alzheimer: un studiu pilot deschis folosind actigrafie. J. Clin. Psihofarmacol. 2004 , 24 , 456–459. [ Google Scholar ] [ PubMed ]
Mahlberg, R.; Walther, S. Actigrafie la pacientii agitati cu dementa. Monitorizarea rezultatelor tratamentului. Z. Gerontol. Geriatr. 2007 , 40 , 178–184. [ Google Scholar ] [ CrossRef ]
Anderson, KN; Jamieson, S.; Graham, AJ; Shneerson, JM Tulburare de comportament în somn REM tratată cu melatonină la un pacient cu boala Alzheimer. Clin. Neurol. Neurochirurgie. 2008 , 110 , 492–495. [ Google Scholar ] [ CrossRef ]
Dowling, GA; Burr, RL; van Someren, EJ; Hubbard, EM; Luxenberg, JS; Mastick, J.; Cooper, BA Melatonina și tratamentul cu lumină puternică pentru întreruperea activității de odihnă la pacienții instituționalizați cu boala Alzheimer. J. Am. Geriatr. Soc. 2008 , 56 , 239–246. [ Google Scholar ] [ CrossRef ]
Gehrman, PR; Connor, DJ; Martin, JL; Shochat, T.; Corey-Bloom, J.; Ancoli-Israel, S. Melatonin nu reușește să îmbunătățească somnul sau agitația într-un studiu dublu-orb randomizat controlat cu placebo de pacienți instituționalizați cu boala Alzheimer. A.m. J. Geriatr. Psihiatrie 2009 , 17 , 166–169. [ Google Scholar ] [ CrossRef ]
Farias, ST; Mungas, D.; Reed, BR; Harvey, D.; DeCarli, C. Progresia deficienței cognitive ușoare la demență în cohortele clinice vs comunitare. Arc. Neurol. 2009 , 66 , 1151–1157. [ Google Scholar ] [ PubMed ]
Davies, L.; Wolska, B.; Hilbich, C.; Multhaup, G.; Martins, R.; Simms, G.; Beyreuther, K.; Masters, depunerea de proteină amiloid CL A 4 și diagnosticul bolii Alzheimer: Prevalența în creierul îmbătrânit determinată prin imunocitochimie în comparație cu tehnicile neuropatologice convenționale. Neurologie 1988 , 38 , 1688–1693. [ Google Scholar ] [ CrossRef ]
Preț, JL; Morris, JC Încurcături și plăci în îmbătrânirea nedementabilă și boala Alzheimer „preclinic”. Ann. Neurol. 1999 , 45 , 358–368. [ Google Scholar ] [ CrossRef ]
Braak, H.; Braak, E. Stadializarea modificărilor neurofibrilare legate de boala Alzheimer. Neurobiol. Aging 1995 , 16 , 271–278. [ Google Scholar ] [ CrossRef ]
Braak, H.; Braak, E. Evoluția modificărilor neuronale în cursul bolii Alzheimer. J. Transm. Neural. Suppl. 1998 , 53 , 127–140. [ Google Scholar ] [ CrossRef ]
Furio, AM; Brusco, LI; Cardinali, DP Valoarea terapeutică posibilă a melatoninei în deficiența cognitivă ușoară. Un studiu retrospectiv. J. Pineal Res. 2007 , 43 , 404–409. [ Google Scholar ] [ CrossRef ]
Cardinali, DP; Vigo, DE; Olivar, N.; Vidal, MF; Furio, AM; Brusco, LI Aplicarea terapeutică a melatoninei în deficiența cognitivă ușoară. A.m. J. Neurodegener Dis. 2012 , 1 , 280–291. [ Google Scholar ] [ PubMed ]
Wu, YH; Swaab, DF Tulburări și strategii pentru reactivarea sistemului de ritm circadian în îmbătrânire și boala Alzheimer. Sleep Med. 2007 , 8 , 623–636. [ Google Scholar ] [ CrossRef ]
Pandi-Perumal, SR; BaHammam, AS; Brown, GM; Spence, DW; Bharti, VK; Kaur, C.; Hardeland, R.; Cardinali, DP Apărarea antioxidativă a melatoninei: implicații terapeutice pentru procesele de îmbătrânire și neurodegenerative. Neurotox. Res. 2013 , 23 , 267–300. [ Google Scholar ] [ CrossRef ]
Rosales-Corral, SA; Acuña-Castroviejo, D.; Coto-Montes, A.; Boga, JA; Manchester, LC; Fuentes-Broto, L.; Korkmaz, A.; Ma, S.; Tan, DX; Reiter, RJ Boala Alzheimer: mecanisme patologice și rolul benefic al melatoninei. J. Pineal Res. 2012 , 52 , 167–202. [ Google Scholar ] [ CrossRef ]
Tan, DX; Manchester, LC; Sanchez-Barcelo, E.; Mediavilla, MD; Reiter, RJ Semnificația nivelurilor ridicate de melatonină endogene în lichidul cefalorahidian al mamiferelor și în sistemul nervos central. Curr. Neurofarmacol. 2010 , 8 , 162–167. [ Google Scholar ] [ CrossRef ]
Legros, C.; Chesneau, D.; Boutin, JA; Barc, C.; Malpaux, B. Melatonina din lichidul cefalorahidian, dar nu din sânge, ajunge la țesuturile cerebrale de oaie în condiții fiziologice. J. Neuroendocrinol. 2014 . [ Google Scholar ] [ CrossRef ]
Monti, JM; Alvarino, F.; Cardinali, DP; Savio, I.; Pintos, A. Studiu polisomnografic al efectului melatoninei asupra somnului la pacienții vârstnici cu insomnie primară cronică. Arc. Gerontol. Geriatr. 1999 , 28 , 85–98. [ Google Scholar ] [ CrossRef ]
Cardinali, DP; Srinivasan, V.; Brzezinski, A.; Brown, GM Melatonin și analogii săi în insomnie și depresie. J. Pineal Res. 2012 , 52 , 365–375. [ Google Scholar ] [ CrossRef ]
Rajaratnam, SM; Polymeropoulos, MH; Fisher, DM; Roth, T.; Scott, C.; Birznieks, G.; Klerman, EB Tasimelteon agonist al melatoninei (VEC-162) pentru insomnia tranzitorie după schimbarea timpului de somn: două studii multicentre controlate randomizate. Lancet 2009 , 373 , 482–491. [ Google Scholar ] [ CrossRef ]
Mulchahey, JJ; Goldwater, DR; Zemlan, FP Un singur studiu orb, controlat cu placebo, pe grupuri de creștere a dozei privind siguranța, tolerabilitatea, farmacocinetica și farmacodinamia analogului melatoninei β-metil-6-cloromelatoninei. Life Sci. 2004 , 75 , 1843–1856. [ Google Scholar ] [ CrossRef ]
McKenna, JT; Christie, MA; Jeffrey, BA; McCoy, JG; Lee, E.; Connolly, NP; Ward, CP; Strecker, RE Tratamentul cronic cu ramelteon într-un model de șoarece al bolii Alzheimer. Arc. Ital. Biol. 2012 , 150 , 5–14. [ Google Scholar ] [ PubMed ]
Ursing, C.; Hartter, S.; von Bahr, C.; Tybring, G.; Bertilsson, L.; Rojdmark, S. Metabolismul hepatic al melatoninei afectează nivelul melatoninei serice endogen la om? J. Endocrinol. Investig. 2002 , 25 , 459–462. [ Google Scholar ] [ CrossRef ]
Hartter, S.; Grozinger, M.; Weigmann, H.; Roschke, J.; Hiemke, C. Biodisponibilitate crescută a melatoninei orale după administrarea concomitentă a fluvoxaminei. Clin. Pharmacol. Acolo. 2000 , 67 , 1–6. [ Google Scholar ] [ CrossRef ]
Hartter, S.; Nordmark, A.; Trandafir, DM; Bertilsson, L.; Tybring, G.; Laine, K. Efectele consumului de cofeină asupra farmacocineticii melatoninei, un medicament probă pentru activitatea CYP1A2. Br. J. Clin. Pharmacol. 2003 , 56 , 679–682. [ Google Scholar ] [ CrossRef ]
Souetre, E.; Salvati, E.; Belugou, JL; de Galeani, B.; Krebs, B.; Ortonne, JP; Darcourt, G. 5-Methoxypsoralen crește nivelurile plasmatice de melatonina la om. J. Investig. Dermatol. 1987 , 89 , 152–155. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Garde, E.; Micic, S.; Knudsen, K.; Angelo, HR; Wulf, HC 8-Methoxypsoralen crește nivelurile plasmatice de melatonina în timpul zilei la oameni prin inhibarea metabolismului. Fotochimie. Fotobiol. 1994 , 60 , 475–480. [ Google Scholar ] [ CrossRef ]
Mauviard, F.; Raynaud, F.; Geoffriau, M.; Claustrat, B.; Pevet, P. 5-Methoxypsoralen inhibă 6-hidroxilarea melatoninei la șobolan. Biol. Semnale 1995 , 4 , 32–41. [ Google Scholar ] [ CrossRef ]
Weishaupt, JH; Bartels, C.; Polking, E.; Dietrich, J.; Rohde, G.; Poeggeler, B.; Mertens, N.; Sperling, S.; Bohn, M.; Huther, G.; et al. Reducerea daunelor oxidative în SLA prin tratamentul cu doze mari de melatonină enterală. J. Pineal Res. 2006 , 41 , 313–323. [ Google Scholar ] [ CrossRef ]
Chahbouni, M.; Escames, G.; Venegas, C.; Sevilla, B.; Garcia, JA; Lopez, LC; Munoz-Hoyos, A.; Molina-Carballo, A.; Acuña-Castroviejo, D. Tratamentul cu melatonina normalizează citokinele proinflamatorii plasmatice și stresul nitrozativ/oxidativ la pacienții care suferă de distrofie musculară Duchenne. J. Pineal Res. 2010 , 48 , 282–289. [ Google Scholar ] [ CrossRef ]
Voordouw, BC; Euser, R.; Verdonk, RE; Alberda, BT; de Jong, FH; Drogendijk, AC; Fauser, BC; Cohen, M. Melatonina și combinațiile melatonină-progestin modifică funcția hipofizo-ovariană la femei și pot inhiba ovulația. J. Clin. Endocrinol. Metab. 1992 , 74 , 108–117. [ Google Scholar ] [ PubMed ]
Nickkholgh, A.; Schneider, H.; Sobirey, M.; Venetz, WP; Hinz, U.; Pelzle, H.; Gotthardt, DN; Cekauskas, A.; Manikas, M.; Mikalauskas, S.; et al. Utilizarea melatoninei în doze mari în rezecția hepatică este sigură: prima experiență clinică. J. Pineal Res. 2011 , 50 , 381–388. [ Google Scholar ] [ CrossRef ]
Cardinali, DP; Pandi-Perumal, SR; Niles, LP Melatonina și receptorii săi: Funcția biologică în reglementarea circadiană a somnului-veghe. În Neurochimia somnului și a stării de veghe ; Monti, JM, Pandi-Perumal, SR, Sinton, CM, Eds.; Cambridge University Press: Cambridge, Marea Britanie, 2008; p. 283–314. [ Google Scholar ]
Dagan, Y.; Zisapel, N.; Nof, D.; Laudon, M.; Atsmon, J. Inversarea rapidă a toleranței la hipnoticele benzodiazepine prin tratament cu melatonină orală: un raport de caz. EURO. Neuropsihofarmacol. 1997 , 7 , 157–160. [ Google Scholar ] [ CrossRef ]
Garfinkel, D.; Zisapel, N.; Wainstein, J.; Laudon, M. Facilitarea întreruperii benzodiazepinei de către melatonină: O nouă abordare clinică. Arc. Intern. Med. 1999 , 159 , 2456–2460. [ Google Scholar ] [ CrossRef ]
Siegrist, C.; Benedetti, C.; Orlando, A.; Beltran, JM; Tuchscherr, L.; Noseda, CM; Brusco, LI; Cardinali, DP Lipsa modificărilor serice ale prolactinei, FSH, TSH și estradiolului după tratamentul cu melatonină în doze care îmbunătățesc somnul și reduc consumul de benzodiazepine la pacienții cu tulburări de somn, de vârstă mijlocie și vârstnici. J. Pineal Res. 2001 , 30 , 34–42. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Kunz, D.; Bineau, S.; Maman, K.; Milea, D.; Toumi, M. Întreruperea benzodiazepinei cu melatonină cu eliberare prelungită: Sugestii dintr-o bază de date de prescripție longitudinală germană. Opinia expertului. Pharmacother. 2012 , 13 , 9–16. [ Google Scholar ] [ CrossRef ]
Clay, E.; Falissard, B.; Moore, N.; Toumi, M. Contribuția campaniilor de melatonină și anti-benzodiazepină cu eliberare prelungită la reducerea consumului de benzodiazepine și medicamente Z în nouă țări europene. EURO. J. Clin. Pharmacol. 2013 , 69 , 1–10. [ Google Scholar ] [ PubMed ]
Leger, D.; Laudon, M.; Zisapel, N. Excreția nocturnă a 6-sulfatoximelatoninei în insomnie și relația sa cu răspunsul la terapia de substituție cu melatonină. A.m. J. Med. 2004 , 116 , 91–95. [ Google Scholar ] [ CrossRef ]
Zhdanova, IV; Wurtman, RJ; Regan, MM; Taylor, JA; Shi, JP; Leclair, OU Tratament cu melatonina pentru insomnia legata de varsta. J. Clin. Endocrinol. Metab. 2001 , 86 , 4727–4730. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Wilson, SJ; Nutt, DJ; Alford, C.; Argyropoulos, SV; Baldwin, DS; Bateson, AN; Britton, TC; Crowe, C.; Dijk, DJ; Espie, CA; et al. Declarație de consens al Asociației Britanice pentru Psihofarmacologie privind tratamentul bazat pe dovezi al insomniei, parasomniilor și tulburărilor de ritm circadian. J. Psihofarmacol. 2010 , 24 , 1577–1601. [ Google Scholar ] [ CrossRef ]
Cardinali, DP; Furio, AM; Brusco, LI Utilizarea cronobioticelor în resincronizarea ciclului somn/veghe. Aplicație terapeutică în fazele incipiente ale bolii Alzheimer. Brevetul recent. Endocr. Metab. Drug Imun Discov. 2011 , 5 , 80–90. [ Google Scholar ] [ CrossRef ]
Jean-Louis, G.; von Gizycki, H.; Zizi, F. Efectele melatoninei asupra somnului, dispoziției și cogniției la vârstnici cu deficiențe cognitive ușoare. J. Pineal Res. 1998 , 25 , 177–183. [ Google Scholar ] [ CrossRef ]
Peck, JS; LeGoff, DB; Ahmed, I.; Goebert, D. Efectele cognitive ale administrării melatoninei exogene la persoanele în vârstă: un studiu pilot. A.m. J. Geriatr. Psihiatrie 2004 , 12 , 432–436. [ Google Scholar ] [ PubMed ]
Wade, AG; Ford, I.; Crawford, G.; McMahon, AD; Nir, T.; Laudon, M.; Zisapel, N. Eficacitatea melatoninei cu eliberare prelungită la pacienții cu insomnie cu vârsta cuprinsă între 55 și 80 de ani: calitatea somnului și rezultatele de vigilență în ziua următoare. Curr. Med. Res. Opinează. 2007 , 23 , 2597–2605. [ Google Scholar ] [ CrossRef ]
Riemersma-van der Lek, RF; Swaab, DF; Twisk, J.; Hol, EM; Hoogendijk, WJ; van Someren, EJ Efectul luminii strălucitoare și al melatoninei asupra funcției cognitive și noncognitive la rezidenții vârstnici ai unităților de îngrijire de grup: un studiu controlat randomizat. JAMA 2008 , 299 , 2642–2655. [ Google Scholar ] [ CrossRef ]
Garzon, C.; Guerrero, JM; Aramburu, O.; Guzman, T. Efectul administrării melatoninei asupra somnului, tulburărilor de comportament și întreruperii medicamentelor hipnotice la vârstnici: un studiu randomizat, dublu-orb, controlat cu placebo. Aging Clin. Exp. Res. 2009 , 21 , 38–42. [ Google Scholar ] [ CrossRef ]
Cazzola, R.; Rondanelli, M.; Faliva, M.; Cestaro, B. Efectele suplimentării cu DHA-fosfolipide, melatonină și triptofan asupra proprietăților fizico-chimice ale membranei eritrocitelor la pacienții vârstnici care suferă de deficiență cognitivă ușoară. Exp. Gerontol. 2012 , 47 , 974–978. [ Google Scholar ] [ CrossRef ]
Rondanelli, M.; Opizzi, A.; Faliva, M.; Mozzoni, M.; Antoniello, N.; Cazzola, R.; Savare, R.; Cerutti, R.; Grossi, E.; Cestaro, B. Efectele unei integrări dietetice cu o emulsie uleioasă de DHA-fosfolipide care conțin melatonină și triptofan la pacienții vârstnici care suferă de deficiență cognitivă ușoară. Nutr. Neurosci. 2012 , 15 , 46–54. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Boala Alzheimer (AD), o tulburare neurodegenerativă legată de vârstă, cu deficit de cogniție progresiv, este caracterizată prin plăci senile extracelulare (SP) de β-amiloid (Aβ) agregat și încurcături neurofibrilare intracelulare, care conțin în principal proteina tau hiperfosforilată asociată microtubulilor. Factori multipli contribuie la etiologia AD în ceea ce privește inițierea și progresia. Melatonina este un hormon produs endogen în creier și scade în timpul îmbătrânirii și la pacienții cu AD. Datele din studiile clinice indică faptul că suplimentarea cu melatonină îmbunătățește somnul, ameliorează apusul soarelui și încetinește progresia deteriorării cognitive la pacienții cu AD. Melatonina protejează eficient celulele neuronale de toxicitatea mediată de Aβ prin proprietăți antioxidante și anti-amiloide. Nu numai că inhibă generarea de Aβ, dar oprește și formarea fibrilelor de amiloid printr-o interacțiune dependentă de structură cu Aβ. Studiile noastre au demonstrat că melatonina atenuează eficient hiperfosforilarea tau asemănătoare cu Alzheimer. Deși mecanismul exact nu este încă pe deplin înțeles, se propune o influență reglatoare directă a melatoninei asupra activităților protein kinazelor și proteinelor fosfataze. În plus, melatonina joacă, de asemenea, un rol în protejarea sistemului colinergic și în antiinflamație. Scopul acestei revizuiri este de a stimula interesul pentru melatonină ca agent potențial util în prevenirea și tratamentul AD.
1. Introducere
Boala Alzheimer (AD) este o boală neurodegenerativă asociată vârstei și caracterizată prin pierderea progresivă a cogniției și alte manifestări neurocomportamentale. Semnele patologice ale AD includ plăci senile extracelulare (SP), constând în principal din β-amiloid (Aβ) și încurcături neurofibrilare intracelulare (NFT), compuse în principal din tau hiperfosforilat anormal, o proteină asociată cu microtubuli [1 ] . În ciuda unui număr mare de studii întreprinse, etiologia AD este în mare parte necunoscută. Au fost propuse multe mecanisme, inclusiv predispoziții genetice (de exemplu, niveluri de expresie și subforme ale presenilinelor (PS) și apolipoproteinei (Apo) E), procese inflamatorii asociate cu eliberarea de citokine, stres oxidativ și neurotoxicitate de către ionii metalici [ 2 – 6 ] .].
Melatonina ( N -acetil-5-metoxitriptamina), un metabolit al triptofanului și sintetizat în principal în glanda pineală, are o serie de funcții fiziologice, inclusiv reglarea ritmurilor circadiene, curățarea radicalilor liberi, îmbunătățirea imunității și, în general, inhibarea oxidării biomoleculelor. Scăderea melatoninei în ser și lichidul cefalorahidian (LCR) și pierderea ritmului diurn al melatoninei sunt observate la pacienții cu AD [ 7 – 12 ]. Mai mult, nivelul melatoninei din LCR scade odată cu progresia neuropatologiei AD, așa cum este determinat de etapele Braak [ 12] .]. Nivelurile melatoninei atât în LCR, cât și în glanda pineală umană postmortem sunt deja reduse la subiecții cu AD preclinici, care sunt încă intacte din punct de vedere cognitiv și au doar cele mai timpurii semne de neuropatologie AD [8 , 12 ] . Există o corelație puternică între conținutul pineal și nivelul melatoninei LCR [ 8 ] și între LCR și nivelurile de melatonină plasmatică [ 7 ], sugerând că un nivel redus de melatonină LCR poate servi ca un marker timpuriu pentru primele etape ale AD. La mamifere, melatonina își exercită unele dintre funcțiile sale prin doi receptori specifici de membrană cu afinitate mare, receptorul 1 al melatoninei (MT1) și receptorul 2 al melatoninei (MT2). Scăderea imunoreactivitatii MT2 și imunoreactivitatea MT1 crescută au fost raportate în hipocampul pacienților cu AD.13 , 14 ]. Deși glanda pineală a pacienților cu AD are modificări moleculare, nu s-au observat modificări ale greutății pineale, calcifierii sau conținutului total de proteine [ 8 , 15 ]. De asemenea, se arată că ARNm a receptorului β1-adrenergic a dispărut, iar activitatea și expresia genică a monoaminoxidazei (MAO) au fost reglate în sens pozitiv la pacienții cu AD, sugerând că dereglarea inervațiilor noradrenergice și epuizarea serotoninei, precursorul melatoninei, ar putea fi responsabil pentru pierderea ritmului melatoninei și reducerea nivelului de melatonina în AD [ 16]. Suplimentarea cu melatonină a fost sugerată pentru a îmbunătăți ritmicitatea circadiană, de exemplu, scăderea comportamentului agitat, a confuziei și a „apusului soarelui” și pentru a produce efecte benefice asupra memoriei la pacienții cu AD [17-21 ] . Prin urmare, suplimentarea cu melatonină, cu toxicitatea sa scăzută marcată [ 22 – 24 ], poate fi una dintre posibilele strategii pentru tratamentul simptomatic.
În AD, se crede, în general, că Aβ joacă un rol important în promovarea degenerării neuronale, făcând neuronii mai vulnerabili la creșterile legate de vârstă ale nivelurilor de stres oxidativ și deteriorări în metabolismul energetic celular [25 ] . Ca proteină principală asociată cu microtubuli, tau promovează asamblarea microtubulilor și stabilizează microtubulii. Hiperfosforilarea va reduce în mod evident abilitățile tau, ceea ce duce la perturbarea aranjamentului citoscheletic [ 26 , 27 ]. Amploarea patologiei neurofibrilare, și în special numărul de NFT cortical, se corelează pozitiv cu severitatea demenței [ 28] .]. Deoarece melatonina este capabilă să îmbunătățească unele dintre simptomele clinice ale AD, iar nivelul melatoninei scade dramatic în timpul AD, studiile privind relația dintre melatonină și patologia AD vor fi utile pentru a evalua potențialul său în prevenirea sau tratamentul AD. În această revizuire, vom aborda rolul melatoninei în hiperfosforilarea tau și toxicitatea Aβ. Deoarece deficitul colinergic și inflamația sunt implicate în patogenia AD, se introduce și protecția melatoninei asupra sistemului colinergic și a inflamației. Fiecare parte este descrisă, de la observarea fenomenului până la investigarea mecanismelor și speculații.
Tau hiperfosforilat a fost identificat în mai mult de o duzină de tulburări neurodegenerative, denumite tauopatii, inclusiv AD, boala Niemann-Pick de tip C și așa mai departe [ 29-31 ]. Dintre aceste tulburări legate de tau, AD este cea mai frecventă și cea mai bine studiată tauopatie. În creierul AD, nivelul tau-ului hiperfosforilat este de 3-4 ori mai mare decât cel al tau-ului din creierul adult normal [ 32 , 33] .]. Există 79 de situsuri presupuse de fosforilare a serinei sau treoninei în cea mai lungă izoformă tau a creierului uman și mai mult de 30 de situsuri de fosforilare au fost identificate în creierul AD, inclusiv Thr39, Ser46, Thr50, Thr69, Thr153, Thr175, Thr181, Ser184, Ser185. , Ser198, Ser199, Ser202, Thr205, Ser208, Ser210, Thr212, Ser214, Thr217, Thr231, Ser235, Ser237, Ser238, Ser245, Ser258, Ser262, Ser285, Ser262, Ser285, Ser293, Ser3, Ser3, Ser30, Ser293, Ser3, Ser3, Ser35 r377, Ser396 , Ser400, Thr403, Ser404, Ser409, Ser412, Ser413, Ser416 şi Ser422 [ 34 – 39 ]. Răspândirea patologiei tau în creier este semnul distinctiv al patogenezei AD, iar numărul de NFT este corelat pozitiv cu deficitul cognitiv clinic al pacienților cu AD [ 40 ].
Inhibarea hiperfosforilării tau este o țintă în tratamentul AD. Astfel, am studiat sistemic efectul melatoninei asupra hiperfosforilării tau și am constatat că melatonina atenuează eficient hiperfosforilarea tau indusă de wortmannin [ 41 ], calyculin A (CA) [ 42 – 44 ] și acid okadaic [ 45 ] în neuroblastom N2a și SH-SY5Y. celule. S-a demonstrat în continuare că melatonina a ameliorat semnificativ hiperfosforilarea tau provocată de wortmannin [ 46 ], izoproterenol [ 47 , 48 ], CA [ 44 ] și iluminarea constantă a luminii [ 24]] la șobolani. Pentru a elucida mecanismele care stau la baza efectului inhibitor al melatoninei asupra hiperfosforilării tau, au fost detectate modificări ale activităților protein kinazelor și fosfatazelor. Tratamentul cu melatonină nu numai că a inhibat activarea glicogen sintazei kinazei-3 (GSK-3) indusă de wortmannin, activarea proteinei kinazei A (PKA) indusă de izoproterenol și inactivarea proteinei fosfatazei-2A (PP-2A) indusă de CA, dar a antagonizat și oxidarea. stresul indus de acești agenți [ 46 , 49 , 50 ]. Aceste rezultate din studiile noastre oferă dovezi pentru eficacitatea puternică a melatoninei în inhibarea hiperfosforilării tau.
Pentru a explora dacă o scădere a melatoninei ar induce alterarea fosforilării tau, am inhibat biosinteza melatoninei prin injectarea creierului de haloperidol, un inhibitor al 5-hidroxiindol-O-metiltransferazei (o enzimă cheie în sinteza melatoninei) la șobolani [ 51 ]. S-a descoperit că inhibarea biosintezei melatoninei nu numai că a dus la afectarea memoriei spațiale la șobolani, dar a indus și o creștere a fosforilării tau cu o scădere concomitentă a activității PP-2A. Suplimentarea cu melatonină prin injectare prealabilă timp de o săptămână și întărirea în timpul perioadei de administrare a haloperidolului au îmbunătățit semnificativ deficitele de retenție a memoriei, au oprit hiperfosforilarea tau și stresul oxidativ și au restabilit activitatea PP-2A [51] .]. De asemenea, am folosit iluminare constantă pentru a întrerupe metabolismul melatoninei la șobolani. Concomitent cu scăderea melatoninei serice, șobolanii luminați în mod constant au dezvoltat deficite de memorie spațială, hiperfosforilarea tau la mai multe locuri, activarea GSK-3 și PKA, precum și suprimarea PP-1. Leziuni oxidative proeminente și leziuni de organele, demonstrate prin exprimarea crescută a proteinelor legate de stresul reticulului endoplasmatic (ER), inclusiv proteina de legare a imunoglobulinei (BiP)/GRP78 și CHOP/GADD153, au scăzut numărul de ER rugoase și ribozomi liberi, au dus la mai subțire. sinapsele și creșterea superoxid dismutazei și MAO, care au fost observate și la șobolanii expuși la lumină. Suplimentarea simultană cu melatonină a oprit parțial deficiențele comportamentale și moleculare [ 24]. Deși nu este clar dacă concentrația scăzută de melatonina este un factor cauzal în patologia AD sau doar un proces secundar, rezultatele noastre implică puternic rolul important al scăderii melatoninei în afectarea memoriei spațiale asemănătoare Alzheimer și hiperfosforilarea tau.
Agenții chimici utilizați în studiile noastre, inclusiv wortmannin, izoproterenol și CA, nu numai că au indus fosforilarea tau, dar au inițiat și stresul oxidativ, manifestat printr – un nivel crescut de malondialdehidă și o activitate alterată a superoxid dismutază (SOD) [41-43 ] . În plus, melatonina este un puternic captator direct de radicali liberi și un antioxidant indirect care acționează prin creșterea activității mai multor enzime antioxidante importante, cum ar fi SOD, glutation peroxidaza și glutation reductază [52 ] . Iluminarea constantă nu numai că a indus scăderea nivelului de melatonină seric, ci și nivelurile crescute de SOD și MAO la șobolani [ 24 ]. Se știe că stresul oxidativ influențează starea de fosforilare a tau [ 53– 55 ]. Acumularea de proteine greșite și agregate în neuronii creierului AD a fost considerată a fi legată de stresul oxidativ, împreună cu modificările structurii sale moleculare odată cu îmbătrânirea [ 56 ]. Ca antioxidant și captator de radicali liberi [ 57 – 59 ], melatonina previne supraproducția de radicali liberi și reduce daunele neuronale rezultate dintr-o varietate de procese patologice [ 60 – 62 ]. Prin urmare, este posibil ca prevenirea fosforilării tau de către melatonină să se datoreze parțial activității sale antioxidante.
Unele studii au indicat, de asemenea, că melatonina poate acționa ca un modulator al enzimei într-un mod care nu are legătură cu proprietățile sale antioxidante. Datele acumulate oferă dovezi pentru reglarea de către melatonină a protein kinazelor, inclusiv PKA [ 63 , 64 ], protein kinaza C (PKC) [ 65 , 66 ], Ca 2+ /calmodulin-dependent kinaze II (CaMKII) [ 67 , 68 ] și familia protein kinazei activate de mitogen (MAPK) [ 69 – 72 ]. Corelația documentată între melatonină și AMPc indică faptul că melatonina ar putea inhiba activitatea PKA prin inhibarea cuplată a receptorului de melatonină a adenilil ciclazei și reducerea AMPc [ 63 ]., 64 ]. Deși nu există dovezi ale unei relații directe între melatonină și activitatea GSK-3, unul dintre studiile noastre a constatat că tratamentul cu melatonină a revizuit iluminarea constantă a luminii indusă activarea GSK-3 în creierul șobolanilor [24 ] .
Aβ, care compune 39-43 de resturi de aminoacizi derivate din precursorul său, proteina precursor de amiloid (APP), joacă un rol esențial în patogeneza AD [ 25 , 73 ]. APP matur este procesat proteolitic prin căi distincte de α-secretază sau β-secretază [ 74 ]. Calea α non-amiloidogenă implică scindarea APP în secvența Ap de către α-secretază pentru a elibera un fragment APP N-terminal, care, la rândul său, este scindat de y-secretază. Astfel, scindarea de către y-secretază exclude formarea Ap. Calea β-secretazei amiloidogenă, care are ca rezultat formarea peptidei Aβ intacte, este mediată de scindarea secvenţială a β-secretazei şi γ-secretazei la nivelul N- şi C.-terminale ale secvenței Aβ, respectiv [ 75 ]. S-a descoperit că melatonina inhibă nivelurile normale de secreție de APP solubilă (sAPP) în diferite linii celulare prin interferarea cu maturarea completă a APP [ 76 ]. În plus, administrarea melatoninei a redus eficient generarea și depunerea Aβ atât in vivo [ 77 , 78 ], cât și in vitro [ 76 , 79 – 81 ]. Am demonstrat că melatonina reduce generarea de Ap în celulele N2a de neuroblastom de șoarece care adăpostesc APP695 [ 80 , 82 ].
Cu toate acestea, un studiu in vivo a arătat că melatonina nu a afectat expresia holoproteinei APP la șoarecii transgenici Tg2576 [ 77 ]. Mai mult, în ciuda atingerii unor concentrații plasmatice mari de melatonină, terapia cronică cu melatonină la șoarecii bătrâni Tg2576 inițiată la vârsta de 14 luni nu numai că nu a reușit să îndepărteze plăcile existente, dar nu a reușit să prevină depunerea suplimentară de Aβ [83 ] . Acest rezultat este în contrast cu cele ale scăderii Ap la șoarecii de tip sălbatic tratați cu melatonină [ 78 ] și nitrarea redusă a Ap și proteinei la șoarecii Tg2576 tratați cu melatonină [ 77 ]. Timpul de inițiere a tratamentului cu melatonină ar putea explica diferența dintre studiile lui Matsubara și colab. [ 77] și Quinn și colab. [ 83 ], în care a fost utilizat același model de șoarece transgenic Tg2576. Patologia plăcii de amiloid apare de obicei la șoarecii Tg2576 de 10-12 luni [ 84 ]. Tratamentul cu melatonină în studiul lui Matsubara și colab. a fost început când șoarecii aveau patru luni (înainte de apariția plăcilor hipocampale și corticale) [ 77 ], o etapă patologică mai timpurie comparativ cu vârsta de 14 luni în studiul lui Quinn și colab. [ 83]. Cu toate acestea, ambele studii sunt de acord în a găsi puține dovezi ale efectelor antioxidante puternice ale melatoninei la cei mai bătrâni șoareci. Aceste descoperiri indică faptul că melatonina are capacitatea de a regla metabolismul APP și de a preveni patologia Ap, dar nu reușește să exercite efecte anti-amiloid sau antioxidante atunci când este inițiată după depunerea Ap. Deși s-au obținut concluzii consistente, niciunul dintre studiile înrudite nu explică în continuare modul în care melatonina își exercită efectul inhibitor asupra generării Ap. Scindarea proteolitică a APP de către calea α-secretazei este reglată de mulți stimuli fiziologici și patologici, iar mecanismul dependent de PKC este unul dintre cei mai recunoscuti. Stimulii, cum ar fi agoniştii muscarinici şi metabotrofici ai receptorilor de glutamat şi esterii de forbol, au capacitatea de a stimula secreţia de APP solubilă şi de a inhiba formarea de Aβ prin activarea PKC.75 ]. Mecanismul prin care activitatea PKC crește secreția de APP solubilă este încă necunoscut, dar poate implica pași adiționali de kinază și eventuala activare a secretazelor care au mediat clivajul APP. Recent, reglarea inhibitorie prin inhibarea GSK-3 asupra generării Aβ a fost bine stabilită [ 85-87 ]. Mecanismul din spatele acestui lucru nu este clar. S-a demonstrat că inhibarea GSK-3 și reglarea în sus a kinazei N -terminale c-Jun (JNK) au ca rezultat o activitate crescută a metaloproteazei matricei și o degradare crescută a Ap [ 88 ]. Deoarece fosforilarea GSK-3 duce la inactivarea sa, datele sugerează că GSK-3 activat poate inhiba sau reduce activarea JNK de către anumiți stimuli [ 89] .]. GSK-3 interacționează cu presenilin-1 (PS1), un cofactor pentru y-secretaza, ceea ce implică faptul că GSK-3 poate funcționa ca o componentă în complexul y-secretază [90 , 91 ] . Presupunând că melatonina poate influența activitatea PKC și GSK-3 așa cum sa menționat mai devreme, se postulează că melatonina poate regla procesarea APP prin căile PKC și GSK-3. Deoarece PKC este un regulator în amonte al GSK-3, GSK-3 poate fi una dintre căile de semnal obișnuite care cresc generarea de Ap și hiperfosforilarea tau. Reglarea formării fibrilelor Aβ și proprietățile patologice importante ale Apei, cum ar fi neurotoxicitatea și rezistența la degradarea proteolitică, depind de capacitatea peptidelor de a forma structuri β-sheet și/sau fibrile de amiloid [92 , 93 ] .
Intervenția în procesul de agregare Aβ poate fi considerată o abordare pentru oprirea sau încetinirea progresiei AD. Melatonina poate interacționa cu Aβ40 și Aβ42 și poate inhiba formarea progresivă a fibrilelor β-sheet și/sau amiloid [ 94 – 96 ]. Efectul anti-fibrilogenic al melatoninei a fost demonstrat prin diferite tehnici, inclusiv spectroscopie circulară cu dicroism (CD), microscopie electronică și spectroscopie prin rezonanță magnetică nucleară (RMN) și spectrometrie de masă cu ionizare prin electrospray (ESI-MS) [95 ] . Mai mult, interacțiunea dintre melatonină și Aβ pare să depindă de caracteristicile structurale ale melatoninei, mai degrabă decât de proprietățile sale antioxidante, deoarece nu ar putea fi reprodusă de analogii melatoninei sau de alți captatori de radicali liberi.92 , 94 ]. Dovezile derivate din ESI-MS au demonstrat că a existat o interacțiune hidrofobă între Ap și melatonină, iar investigațiile proteolitice au sugerat că interacțiunea a avut loc pe cele 29-40 de reziduuri ale segmentului Aβ [95 ] . Rezultatele spectroscopiei RMN au confirmat în continuare o interacțiune specifică reziduului între melatonină și oricare dintre cele trei reziduuri de histidină și aspartat ale Ap [ 96 ]. Punțile de sare imidazol-carboxilat formate de lanțurile laterale ale reziduurilor de histidină (His+) și aspartat (Asp−) sunt esențiale pentru formarea structurilor de foi de amiloid β [97], iar ruperea acestor punți de sare promovează dizolvarea fibrilei [ 98 ] ].
Melatonina ar putea promova conversia foilor β în bobine aleatorii prin întreruperea punților de sare imidazol-carboxilat și, astfel, să prevină fibrilogeneza și agregarea Ap. Prin urmare, este posibil ca prin blocarea formării conformației secundare a foii p, melatonina nu numai că poate reduce neurotoxicitatea, ci și să faciliteze clearance-ul peptidei prin degradarea proteolitică crescută. S-a demonstrat că melatonina interacționează direct cu Aβ și previne agregarea acestuia [ 99 ]. Tratamentul cu Aβ provoacă un spectru de leziuni celulare, inclusiv creșterea peroxidării lipidelor și a concentrației de calciu liber intracelular, deteriorarea oxidativă a ADN-ului mitocondrial și apariția markerilor apoptotici [100] .]. Mitocondriile nu sunt doar locul principal de generare a speciilor reactive de oxigen (ROS), ci și ținta principală de atac pentru ROS. S-a considerat că melatonina stabilizează fluiditatea membranelor interne mitocondriale; iar legarea de membranele mitocondriale a fost dezvăluită [ 101]. Stresul oxidativ acționează sinergic cu perturbarea homeostaziei calciului intracelular. Deteriorarea membranei indusă de radicalii liberi induce un aflux suplimentar de calciu, iar influxul accentuat de calciu rezultat, la rândul său, va induce generarea de alți radicali liberi. Prin urmare, stresul oxidativ joacă un rol central în neurotoxicitatea indusă de Ap și chiar în moartea celulelor. Pe lângă faptul că Aβ provoacă stres oxidativ, s-a propus că daunele oxidative ar putea exacerba un cerc vicios, în care procesarea amiloidogenă a APP ar fi facilitată și mai mult pentru a genera mai mult Ap, care, la rândul său, crește stresul oxidativ [102 ] . Acumularea datelor implică faptul că melatonina protejează eficient celulele împotriva daunelor oxidative induse de Aβ și a morții celulare in vitro [ 103 ,104 ] şi in vivo [ 77 , 105 – 107 ]. În celulele și animalele tratate cu Aβ, melatonina își exercită activitatea de protecție în principal printr-un efect antioxidant, în timp ce în celulele transfectate cu APP și modelele animale transgenice, mecanismul de bază este atribuit nu numai proprietății sale antioxidante, ci și proprietății sale anti-amiloid. incluzând inhibarea atât a generării de Ap cât și a formării de foi de p și/sau fibrile de amiloid.
În plus, unele descoperiri sugerează un rol pentru semnalizarea perturbată a melatoninei în tulburările de somn care sunt comune la pacienții cu AD [ 108-110 ]. Prin microdializă in vivo la șoarecii Tg2576, s-a constatat că cantitatea de lichid interstițial cerebral (ISF) Ap a corelat cu starea de veghe. În plus, ISF Aβ a crescut semnificativ și în timpul privării acute de somn, dar a scăzut cu perfuzia unui antagonist dublu al receptorului de orexină. Restricția cronică a somnului a crescut semnificativ formarea plăcii Aβ [ 111 ]. Mai mult, administrarea melatoninei a redus eficient generarea și depunerea de Aβ, atât in vivo , cât și in vitro [ 76 – 81] .]. Astfel, melatonina poate inhiba generarea și încărcarea Ap, dar mecanismul necesită investigații suplimentare.
4. Protecția Melatoninei asupra sistemului colinergic
Deficitul sistemului colinergic este, de asemenea, un eveniment precoce și primar în patogeneza AD [ 112 ]. Neuronii din nucleul bazal al lui Meynert, o sursă majoră de inervație colinergică a cortexului cerebral și a hipocampului, suferă o degenerare profundă și selectivă în creierul AD [ 113 – 115 ]. Nivelul de acetilcolină (ACh) este scăzut în stadiul incipient al AD, în timp ce activitățile enzimei de sinteză, colin acetiltransferazei (ChAT) și enzimei de hidrolizare, acetilcolinesteraza (AChE), nu se modifică până în stadiul târziu al AD. 116 – 118]. Alte investigații biologice ale țesutului din biopsie și autopsie au găsit o scădere profundă a activității ChAT în neocortexul pacienților cu AD, corelând pozitiv cu severitatea demenței [ 102 ]. Deși mecanismul care duce la deficitul de ACh este încă necunoscut, inhibitorul AChE a fost folosit ca tratament și este considerat standardul de îngrijire pentru tratamentul pentru AD ușoară până la moderată [119 ] .
Melatonina are efecte protectoare asupra sistemului colinergic. Un studiu anterior a demonstrat, de asemenea, că melatonina a prevenit parțial inhibarea indusă de peroxinitrit a transportului de colină și a activității ChAT în mai multe proteine neuronale din sinaptozomi și, mai ușor, din veziculele sinaptice [120 ] . În plus, se raportează că tratamentul de patru luni cu melatonină a ameliorat semnificativ modificările neuropatologice, comportamentale și biochimice la șoarecii transgenici APP695 de opt luni cu depunere de Aβ, deficit semnificativ de învățare și memorie și o reducere profundă a activității ChAT în cortexul frontal și hipocampus [ 105]. Un alt studiu a mai arătat că un tratament similar cu melatonina a antagonizat deficitul de memorie spațială și a scăzut activitatea ChAT la șobolanii adulți ovariectomizați [ 121 ]. Cu toate acestea, la șobolanii infuzați intracerebroventricular cu Aβ timp de 14 zile, în care activitatea ChAT a fost redusă semnificativ, melatonina nu a putut restabili activitatea acestei enzime [ 122 ]. Melatonina a arătat inhibarea numai a creșterii induse de lipopolizaharide (LPS) a activității AChE, în timp ce nu au fost observate modificări la șobolanii tratați cu LCR. Aceste rezultate sunt în sprijinul influenței inhibitorii a melatoninei asupra activității AChE în demența indusă de streptozotocină [ 123 ].
În comparație cu placebo, inhibitorii de colinesterază, cum ar fi donepezilul, tacrina, rivastigmina și galantamina, care pot rezolva deprivarea de acetilcolină, sunt capabili să stabilizeze sau să încetinească declinul cogniției, funcției, comportamentului și schimbării globale [124] .]. Secreția de melatonină scade în AD, iar această scădere a fost postulată ca fiind responsabilă pentru dezorganizarea circadiană, scăderea eficienței somnului și afectarea funcției cognitive observate la acești pacienți. Înlocuirea melatoninei s-a dovedit eficientă în tratarea apusului soarelui, a deficienței cognitive ușoare (MCI), un sindrom etiologic eterogen care precede demența și alte tulburări de veghe în somn la pacienții cu AD. Pe lângă inhibarea activității AChE, perspectivele pentru melatonina ca tratament pentru AD se bazează, de asemenea, pe eliminarea speciilor reactive de oxigen și azot, rezolvarea tulburărilor de somn, scăderea toxicității Aβ și a încărcării [125] .]. Nu există dovezi clinice care să arate care este mai bun pentru pacienții cu AD în ceea ce privește inhibitorul AChE și melatonina. Cu toate acestea, combinația dintre aceste două medicamente poate avea efecte mult mai bune. Recent, hibrizii tacrină-melatonină au fost proiectați și sintetizați ca noi candidați multifuncționali pentru AD [ 126 , 127 ]. Compușii prezintă proprietăți colinergice și antioxidante îmbunătățite, fiind inhibitori mai puternici și selectivi ai AChE uman decât tacrina și captând radicalii liberi mai bine decât melatonina. Ele prezintă o toxicitate scăzută și pot fi capabile să pătrundă în sistemul nervos central [ 126 ]. Administrarea directă intracerebrală a unuia dintre acești hibrizi, N-(2-(1H-indol-3-il)etil)-7-(1,2,3,4-tetrahidroacridin-9-ilamino) heptanamidă, scăderea morții celulare induse de Ap și a încărcăturii cu amiloid în parenchimul cerebral al APP /Soareci PS1. Mai mult, reducerea patologiei Aβ a fost însoțită de o recuperare a funcției cognitive [ 127 ].
Un factor comun în patogenia AD este supraactivarea microgliei cu supraexpresia ulterioară a citokinelor proinflamatorii [ 128 – 130 ]. Acumularea de Aβ în plăci, precum și oligomerii Aβ poate produce evenimente inflamatorii/oxidative secvențiale și excitotoxicitate, provocând neurodegenerare și tulburări cognitive [ 131 ]. În plus, studiile epidemiologice au arătat că utilizarea medicamentelor antiinflamatoare nesteroidiene (AINS) scade incidența AD [ 132 ]. S-a demonstrat că Aβ acționează ca un agent proinflamator, activând multe componente inflamatorii, iar SP, înconjurat de microglia și astrocite, coexistă cu citokine și chemokine [ 133] .]. Activarea microgliei indusă de Aβ este considerată a fi una dintre sursele majore ale răspunsului inflamator [ 134 ]. S-a raportat că melatonina atenuează răspunsurile microgliale și astrogliale induse de acid kainic, așa cum este determinat de detectarea imunohistochimică a izolectinei-B4 și a proteinei acide fibrilare gliale (GFAP), markerii specifici pentru microglia și respectiv astroglia [135 ] . Administrarea orală de melatonină a atenuat, de asemenea, citokinele proinflamatorii induse de Aβ, factorul nuclear κ B (NF- κ B) și oxidul nitric (NO) în creierul șobolanului [ 107 ].
Se raportează că melatonina a redus semnificativ răspunsul proinflamator, scăzând cu aproape 50% nivelurile induse de Aβ de citokine proinflamatorii, Interleukin-1-β (IL1-β), Interleukin-6 (IL6) și factorul de necroză tumorală-α (TNF). -α), in vivo [ 107 ]. Mai mult, activitatea de legare a ADN-ului NF- κB a fost inhibată de melatonină [ 136 , 137 ]. Mai recent, s-a demonstrat că melatonina reduce IL-6 indusă de NF- κ B într-o manieră dependentă de concentrație în feliile de creier tratate cu Aβ [ 138 ]. Administrarea melatoninei este, de asemenea, raportată că reduce învățarea indusă de Ap și afectarea memoriei la șobolani, împreună cu o scădere semnificativă a celulelor gliale pozitive care exprimă NF-IL-1β indusă de κ B în plus față de complementul 1q (C1q) în hipocamp [ 139 ].
Melatonina este unul dintre cei mai puternici antioxidanți care acționează la diferite niveluri, iar nivelul melatoninei se reduce în timpul îmbătrânirii și la pacienții cu AD [ 24 , 40 , 140 – 142 ]. În plus, efectele sale antioxidante indirecte și efectele anti-amiloid se bazează pe sprijinul fazării circadiene și acțiunilor anti-excitotoxice adecvate [ 68 , 143] .]. Astfel, nu este surprinzător faptul că melatonina este protectoare în numeroase sisteme experimentale și a fost propusă ca tratament pentru AD. Studii recente de la șoareci transgenici APP au indicat că suplimentarea timpurie, pe termen lung, cu melatonină produce efecte anti-amiloid și antioxidante, dar nu se produce un astfel de efect atunci când tratamentul cu melatonină este inițiat după vârsta formării amiloidului [ 76 , 77 , 79 , 80 ] . Sunt necesare studii clinice extinse și studii cu modele transgenice pentru a confirma rolul melatoninei în stadiul patologic târziu al AD. Dacă melatonina nu are efect în stadiul târziu al AD, studiile asupra melatoninei ar trebui limitate la prevenirea AD, mai degrabă decât la tratament.
Pot apărea reacții adverse ale melatoninei, cum ar fi febră în prima zi de tratament cu melatonină, hiperkinezie sau plângeri de picioare agitate, menoragie, pigmentare la nivelul brațelor și picioarelor, dureri de cap și reacții abdominale, tromboză și somnolență [ 16 , 17 , 144 , 145 .]. În afară de aceste reacții adverse, aplicarea precoce și pe termen lung a melatoninei poate cel puțin încetini dezvoltarea AD. Pe lângă efectele pozitive în sistemele experimentale privind antagonismul deficitului colinergic, inflamația, fibrilogeneza și formarea încurcăturii, efectele de stimulare a somnului și suprimarea apusului sunt rezultate importante care justifică utilizarea melatoninei. Deși există dovezi care să postuleze melatonina ca un instrument util și terapeutic în MCI și AD, studii mai mari, dublu-orb, multicentrice, sunt necesare urgent pentru a explora și investiga în continuare potențialul și utilitatea melatoninei. Deoarece scăderea imunoreactivitatii MT2 și creșterea imunoreactivitatii MT1 au fost raportate în hipocampul pacienților cu AD [ 13 , 14] .], se așteaptă urgent regulatori specifici ai receptorilor de melatonină și noi derivați ai melatoninei.
Lars MI, Jürgen G. Amyloid-β și tau—Un pas de deux toxic în boala Alzheimer. Nat. Pr. Neurosci. 2011; 12 :67–72. [ PubMed ] [ Google Scholar ]2.
Mustapic M., Popovic Hadzija M., Pavlovic M., Pavkovic P., Presecki P., Mrazovac D., Mimica N., Korolija M., Pivac N., Muck-Seler D. Boala Alzheimer și diabetul de tip 2 : Studiul de asociere al polimorfismelor în genele factorului de necroză tumorală-alfa și apolipoproteinei E. Metab. Brain Dis. 2012; 27 :507–512. [ PubMed ] [ Google Scholar ]3.
Ajala T., Rafi J., Wray R., Whitehead MW, Zaidi J. Poate exista o legătură între colestaza intrahepatică a sarcinii și hiperlipidemia combinată familială: un raport de caz. Cauzele J. 2009; 2 doi: 10.4076/1757-1626-2-8679. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]4.
Leszek J., Sochocka M., Gasiorowski K. Factori vasculari și modificări epigenetice în patogeneza bolii Alzheimer. J. Neurol. Sci. 2012; 323 :25–32. [ PubMed ] [ Google Scholar ]5.
Rocchi A., Valensin D., Aldinucci C., Giani G., Barbucci R., Gaggelli E., Kozlowski H., Valensin G. Investigarea metabolomică RMN a astrocitelor a interacționat cu Abeta(42) sau cu complexele acestuia fie cu cuprul (II) sau zinc(II) J. Inorg. Biochim. 2012; 117 :326–333. [ PubMed ] [ Google Scholar ]6.
Rukhsana S., Butterfield DA Rolul stresului oxidativ în progresia bolii Alzheimer. J. Alzheimer Dis. 2010; 19 :341–353. [ PubMed ] [ Google Scholar ]7.
Wu YH, Swaab DF Glanda pineală umană și melatonina în îmbătrânire și boala Alzheimer. J. Pineal Res. 2005; 38 :145–152. [ PubMed ] [ Google Scholar ]8.
Wu YH, Feenstra MG, Zhou JN, Liu RY, Torano JS, van Kan HJ, Fischer DF, Ravid R., Swaab DF Modificări moleculare care stau la baza nivelurilor reduse de melatonina pineală în boala Alzheimer: Modificări în stadiile preclinice și clinice. J. Clin. Endocr. Metab. 2003; 88 :5898–5906. [ PubMed ] [ Google Scholar ]9.
Ferrari E., Arcaini A., Gornati R., Pelanconi L., Cravello L., Fioravanti M., Solerte SB, Magri F. Funcția pineală și hipofizo-adrenocorticală în îmbătrânirea fiziologică și în demența senilă. Exp. Gerontol. 2000; 35 :1239–1250. [ PubMed ] [ Google Scholar ]10.
Ohashi Y., Okamoto N., Uchida K., Iyo M., Mori N., Morita Y. Ritmul zilnic al nivelurilor serice de melatonina și efectul expunerii la lumină la pacienții cu demență de tip Alzheimer. Biol. Psihiatul. 1999; 45 :1646–1652. [ PubMed ] [ Google Scholar ]11.
Liu RY, Zhou JN, van Heerikhuize J., Hofman MA, Swaab DF Scăderea nivelului de melatonina în lichidul cefalorahidian postmortem în raport cu îmbătrânirea, boala Alzheimer și genotipul apolipoproteinei E-epsilon4/4. J. Clin. Endocr. Metab. 1999; 84 :323–327. [ PubMed ] [ Google Scholar ]12.
Zhou JN, Liu RY, Kamphorst W., Hofman MA, Swaab DF Modificările neuropatologice precoce ale Alzheimer la persoanele în vârstă sunt însoțite de scăderea nivelului de melatonină din lichidul cefalorahidian. J. Pineal Res. 2003; 35 :125–130. [ PubMed ] [ Google Scholar ]13.
Savaskan E., Olivieri G., Meier F., Brydon L., Jockers R., Ravid R., Wirz-Justice A., Muller-Spahn F. Imunoreactivitate crescută a receptorilor melatoninei 1a în hipocampul pacienților cu boala Alzheimer . J. Pineal Res. 2002; 32 :59–62. [ PubMed ] [ Google Scholar ]14.
Savaskan E., Ayoub MA, Ravid R., Angeloni D., Fraschini F., Meier F., Eckert A., Muller-Spahn F., Jockers R. Expresia redusă a receptorului de melatonină MT2 hipocampal în boala Alzheimer. J. Pineal Res. 2005; 38 :10–16. [ PubMed ] [ Google Scholar ]15.
Friedland RP, Luxenberg JS, Koss E. Un studiu cantitativ al calcificării intracraniene în demența de tip Alzheimer. Int. Psihogeriatr. 1990; 2 :36–43. [ PubMed ] [ Google Scholar ]16.
Wu YH, Fischer DF, Swaab DF Polimorfismul promotorului A în gena monoaminoxidază A este asociat cu activitatea MAOA pineală la pacienții cu boala Alzheimer. Brain Res. 2007; 1167 :13–19. [ PubMed ] [ Google Scholar ]17.
Cohen-Mansfield J., Garfinkel D., Lipson S. Melatonina pentru tratamentul apusului la persoanele în vârstă cu demență – Un studiu preliminar. Arc. Gerontol. Geriatr. 2000; 31 :65–76. [ PubMed ] [ Google Scholar ]18.
Brusco LI, Marquez M., Cardinali DP Tratamentul cu melatonina stabilizează simptomele cronobiologice și cognitive în boala Alzheimer. Neuro Endocrinol. Lett. 2000; 21 :39–42. [ PubMed ] [ Google Scholar ]19.
Brusco LI, Marquez M., Cardinali DP Gemeni monozigoți cu boala Alzheimer tratați cu melatonină: Raport de caz. J. Pineal Res. 1998; 25 :260–263. [ PubMed ] [ Google Scholar ]20.
Cardinali DP, Brusco LI, Perez Lloret S., Furio AM Melatonina în tulburările de somn și jet-lag. Neuro Endocrinol. Lett. 2002; 23 :9–13. [ PubMed ] [ Google Scholar ]21.
Cardinali DP, Brusco LI, Liberczuk C., Furio AM Utilizarea melatoninei în boala Alzheimer. Neuro Endocrinol. Lett. 2002; 23 :20–23. [ PubMed ] [ Google Scholar ]22.
Karasek M., Reiter RJ, Cardinali DP, Pawlikowski M. Future of melatonin as a therapeutic agent. Neuro Endocrinol. Lett. 2002; 23 :118–121. [ PubMed ] [ Google Scholar ]23.
Singer C., Tractenberg RE, Kaye J., Schafer K., Gamst A., Grundman M., Thomas R., Thal LJ Alzheimers cooperative, SA multicentric, studiu controlat cu placebo al melatoninei pentru tulburările de somn în boala Alzheimer . Dormi. 2003; 26 :893–901. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]24.
Ling ZQ, Tian Q., Wang L., Fu ZQ, Wang XC, Wang Q., Wang JZ Iluminarea constantă induce leziuni asemănătoare Alzheimer cu implicarea reticulului endoplasmatic și protecția melatoninei. J. Alzheimer Dis. 2009; 16 :287–300. [ PubMed ] [ Google Scholar ]25.
Selkoe DJ Biologia celulară a plierii greșite a proteinelor: exemplele bolilor Alzheimer și Parkinson. Nat. Cell Biol. 2004; 6 :1054–1061. [ PubMed ] [ Google Scholar ]26.
Brion JP, Anderton BH, Authelet M., Dayanandan R., Leroy K., Lovestone S., Octave JN, Pradier L., Touchet N., Tremp G. Neurofibrillary tangles and tau phosphorylation. Biochim. Soc. Symp. 2001; 67 :81–88. [ PubMed ] [ Google Scholar ]27.
Billingsley ML, Kincaid RL Fosforilarea și defosforilarea reglementate a proteinei tau: Efecte asupra interacțiunii microtubulilor, traficului intracelular și neurodegenerării. Biochim. J. 1997; 323 :577–591. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]28.
Braak E., Braak H., Mandelkow EM O secvență de modificări ale citoscheletului legate de formarea de încurcături neurofibrilare și fire de neuropil. Acta neuropathol. 1994; 87 :554–567. [ PubMed ] [ Google Scholar ]29.
Avila J., Perez M., Lucas JJ, Gomez-Ramos A., Santa Maria I., Moreno F., Smith M., Perry G., Hernandez F. Assembly in vitro of tau protein and its implications in Alzheimer’s boala. Curr. Alzheimer Res. 2004; 1 :97–101. [ PubMed ] [ Google Scholar ]30.
Sahara N., DeTure M., Ren Y., Ebrahim AS, Kang D., Knight J., Volbracht C., Pedersen JT, Dickson DW, Yen SH și colab. Caracteristicile speciilor Tau hiperfosforilate extractibile cu TBS: intermediari de agregare în creierul de șoarece rTg4510. J. Alzheimer Dis. 2013; 33 :249–263. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]31.
Lei P., Ayton S., Finkelstein DI, Spoerri L., Ciccotosto GD, Wright DK, Wong BX, Adlard PA, Cherny RA, Lam LQ, et al. Deficiența de Tau induce parkinsonismul cu demență prin afectarea exportului de fier mediat de APP. Nat. Med. 2012; 18 :291–295. [ PubMed ] [ Google Scholar ]32.
Khatoon S., Grundke-Iqbal I., Iqbal K. Nivelurile creierului de proteină tau asociată microtubulilor sunt crescute în boala Alzheimer: un test radioimuno-slot-blot pentru nanogramele proteinei. J. Neurochem. 1992; 59 :750–753. [ PubMed ] [ Google Scholar ]33.
Khatoon S., Grundke-Iqbal I., Iqbal K. Nivelurile de tau normal și anormal fosforilat în diferite compartimente celulare și regionale ale bolii Alzheimer și creierul de control. FEBS Lett. 1994; 351 :80–84. [ PubMed ] [ Google Scholar ]34.
Hanger DP, Betts JC, Loviny TL, Blackstock WP, Anderton BH Noi situsuri de fosforilare identificate în tau hiperfosforilat (filament elicoidal împerecheat-tau) din creierul bolii Alzheimer utilizând spectrometria de masă nanoelectrospray. J. Neurochem. 1998; 71 :2465–2476. [ PubMed ] [ Google Scholar ]35.
Hanger DP, Byers HL, Wray S., Leung KY, Saxton MJ, Seereeram A., Reynolds CH, Ward MA, Anderton BH Noile site-uri de fosforilare în tau din creierul Alzheimer susțin un rol pentru cazein kinaza 1 în patogeneza bolii. J. Biol. Chim. 2007; 282 :23645–23654. [ PubMed ] [ Google Scholar ]36.
Hasegawa M., Morishima-Kawashima M., Takio K., Suzuki M., Titani K., Ihara Y. Secvență de proteine și analize spectrometrice de masă ale tau în creierul bolii Alzheimer. J. Biol. Chim. 1992; 267 :17047–1754. [ PubMed ] [ Google Scholar ]37.
Morishima-Kawashima M., Hasegawa M., Takio K., Suzuki M., Yoshida H., Watanabe A., Titani K., Ihara Y. Hyperphosphorylation of tau in PHF. Neurobiol. Îmbătrânire. 1995; 16 :365–371. [ PubMed ] [ Google Scholar ]38.
Wang XF, Dong CF, Zhang J., Wan YZ, Li F., Huang YX, Han L., Shan B., Gao C., Han J. și colab. Proteina tau umană formează complex cu PrP și unii mutanți PrP legați de GSS și fCJD posedă activități de legare mai puternice cu tau in vitro . Mol. Celulă. Biochim. 2008; 310 :49–55. [ PubMed ] [ Google Scholar ]39.
Peng CX, Hu J., Liu D., Hong XP, Wu YY, Zhu LQ, Wang JZ Intervenția glicogen sintaza kinazei-3beta modificată de boală de către melatonină oprește patologia și deficitele de memorie la un model animal Alzheimer. Neurobiol. Îmbătrânire. 2013; 34 :1555–1563. [ PubMed ] [ Google Scholar ]40.
Peter TN, Irina A., Eileen HB, Constantin B., Heiko B., Nigel JC, Rudolph JC, Barbara JC, Peter D., Kelly DT și colab. Corelația modificărilor neuropatologice ale bolii Alzheimer cu statusul cognitiv: o revizuire a literaturii. J. neuropathol. Exp. Neurol. 2012; 71 :362–381. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]41.
Deng YQ, Xu GG, Duan P., Zhang Q., Wang JZ Efectele melatoninei asupra hiperfosforilării tau induse de wortmannin. Acta Pharmacol. Păcat. 2005; 26 :519–526. [ PubMed ] [ Google Scholar ]42.
Li XC, Wang ZF, Zhang JX, Wang Q., Wang JZ Efectul melatoninei asupra hiperfosforilării tau induse de calyculin A. EURO. J. Pharmacol. 2005; 510 :25–30. [ PubMed ] [ Google Scholar ]43.
Li SP, Deng YQ, Wang XC, Wang YP, Wang JZ Melatonina protejează celulele neuroblastomului SH-SY5Y de afectarea neurofilamentului indusă de caliculina A și neurotoxicitatea. J. Pineal Res. 2004; 36 :186–191. [ PubMed ] [ Google Scholar ]44.
Yang X., Yang Y., Fu Z., Li Y., Feng J., Luo J., Zhang Q., Wang Q., Tian Q. Melatonina ameliorează modificările patologice asemănătoare Alzheimer și afectarea reținerii memoriei spațiale induse de calyculin A. J. Psychopharmacol. 2011; 25 :1118–1125. [ PubMed ] [ Google Scholar ]45.
Wang YP, Li XT, Liu SJ, Zhou XW, Wang XC, Wang JZ Melatonina a ameliorat leziunile asemănătoare Alzheimer induse de acid okadaic. Acta Pharmacol. Păcat. 2004; 25 :276–280. [ PubMed ] [ Google Scholar ]46.
Liu SJ, Wang JZ Fosforilarea tau asemănătoare Alzheimer indusă de wortmannin in vivo și atenuarea acesteia de către melatonină. Acta Pharmacol. Păcat. 2002; 23 :183–187. [ PubMed ] [ Google Scholar ]47.
Wang DL, Ling ZQ, Cao FY, Zhu LQ, Wang JZ Melatonina atenuează supraactivarea proteinei kinazei A induse de izoproterenol și hiperfosforilarea tau în creierul șobolanului. J. Pineal Res. 2004; 37 :11–16. [ PubMed ] [ Google Scholar ]48.
Wang XC, Zhang J., Yu X., Han L., Zhou ZT, Zhang Y., Wang JZ Prevenirea hiperfosforilării tau indusă de izoproterenol de către melatonină la șobolan. Acta Pharmacol. Păcat. 2005; 57 :7–12. [ PubMed ] [ Google Scholar ]49.
Avila J. Agregarea Tau în polimeri fibrilari: Taupathies. FEBS Lett. 2000; 476 :89–92. [ PubMed ] [ Google Scholar ]50.
Gong CX, Liu F., Grundke-Iqbal I., Iqbal K. Modificări post-translaționale ale proteinei tau în boala Alzheimer. J. Transm. Neural. 2005; 112 :813–838. [ PubMed ] [ Google Scholar ]51.
Zhu LQ, Wang SH, Ling ZQ, Wang DL, Wang JZ Efectul inhibării biosintezei melatoninei asupra reținerii memoriei spațiale și fosforilării tau la șobolan. J. Pineal Res. 2004; 37 :71–77. [ PubMed ] [ Google Scholar ]52.
Reiter RJ, Acuna-Castroviejo D., Tan DX, Burkhardt S. Daune moleculare mediate de radicali liberi. Mecanisme pentru acțiunile protectoare ale melatoninei în sistemul nervos central. Ann. NY Acad. Sci. 2001; 939 :200–215. [ PubMed ] [ Google Scholar ]53.
Zhu X., Rottkamp CA, Boux H., Takeda A., Perry G., Smith MA Activarea kinazei p38 leagă fosforilarea tau, stresul oxidativ și evenimentele legate de ciclul celular în boala Alzheimer. J. Neuropat. Exp. Neur. 2000; 59 :880–888. [ PubMed ] [ Google Scholar ]54.
Gomez-Ramos A., Diaz-Nido J., Smith MA, Perry G., Avila J. Effect of the lipid peroxidation product acrolein on tau phosphorylation in neuronal cells. J. Neurosci. Res. 2003; 71 :863–870. [ PubMed ] [ Google Scholar ]55.
Lovell MA, Xiong S., Xie C., Davies P., Markesbery WR Inducerea tau hiperfosforilată în culturi primare de neuroni corticali de șobolan mediată de stresul oxidativ și glicogen sintaza kinaza-3. J. Alzheimer Dis. 2004; 6 :659–671. [ PubMed ] [ Google Scholar ]56.
Kenyon CJ Genetica îmbătrânirii. Natură. 2010; 464 :504–512. [ PubMed ] [ Google Scholar ]57.
Paradies G., Petrosillo G., Paradies V., Reiter RJ, Ruggiero FM Melatonina, cardiolipină și bioenergetica mitocondrială în sănătate și boală. J. Pineal Res. 2010; 48 :297–310. [ PubMed ] [ Google Scholar ]58.
Romero A., Egea J., Garcia AG, Lopez MG Efect neuroprotector sinergic al concentrațiilor scăzute combinate de galantamina și melatonină împotriva stresului oxidativ în celulele neuroblastomului SH-SY5Y. J. Pineal Res. 2010; 49 :141–148. [ PubMed ] [ Google Scholar ]59.
Hardeland R., Tan DX, Reiter RJ Kynuramines, metabolites of melatonin and other indoles: The resurrection of a near forgot class of biogenic amines. J. Pineal Res. 2009; 47 :109–126. [ PubMed ] [ Google Scholar ]60.
Jou MJ, Peng TI, Hsu LF, Jou SB, Reiter RJ, Yang CM, Chiao CC, Lin YF, Chen CC Vizualizarea nivelurilor multiple de protecție mitocondrială ale melatoninei împotriva tranziției de permeabilitate mediată de Ca(2+) mitocondrială și nu numai în astrocite ale creierului de șobolan. J. Pineal Res. 2010; 48 :20–38. [ PubMed ] [ Google Scholar ]61.
Hong Y., Palaksha KJ, Park K., Park S., Kim HD, Reiter RJ, Chang KT Melatonina plus terapie neuroreabilitativă bazată pe exerciții pentru leziuni ale măduvei spinării. J. Pineal Res. 2010; 49 :201–209. [ PubMed ] [ Google Scholar ]62.
Das A., McDowell M., Pava MJ, Smith JA, Reiter RJ, Woodward JJ, Varma AK, Ray SK, Banik NL Inhibarea apoptozei de către melatonină în motoneuronii VSC4.1 expuși la stres oxidativ, excitotoxicitate pe glutamat sau Toxicitatea TNF-alfa implică receptorii de melatonină de membrană. J. Pineal Res. 2010; 48 :157–169. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]63.
Schuster C., Williams LM, Morris A., Morgan PJ, Barrett P. Receptorul uman de melatonină MT1 stimulează producția de cAMP în celulele SH-SY5Y ale liniei celulare de neuroblastom uman printr-o cale de transducție a semnalului calciu-calmodulină. J. Neuroendocrinol. 2005; 17 :170–178. [ PubMed ] [ Google Scholar ]64.
Peschke E., Muhlbauer E., Musshoff U., Csernus VJ, Chankiewitz E., Peschke D. Receptorul (MT(1)) influența mediată a melatoninei asupra concentrației cAMP și secreției de insulină a celulelor de insulinom de șobolan INS-1. J. Pineal Res. 2002; 33 :63–71. [ PubMed ] [ Google Scholar ]65.
Rivera-Bermudez MA, Gerdin MJ, Earnest DJ, Dubocovich ML Reglarea ritmicității bazale în activitatea protein kinazei C de către melatonina în celulele nucleului suprachiasmatic de șobolan imortalizate. Neurosci. Lett. 2003; 346 :37–40. [ PubMed ] [ Google Scholar ]67.
Benitez-King G., Rios A., Martinez A., Anton-Tay F. Inhibarea in vitro a activității kinazei II dependente de Ca2 + /calmodulină de către melatonină. Biochim. Biophys. Acta. 1996; 1290 :191–196. [ PubMed ] [ Google Scholar ]68.
Chen S., Xu Y., Xu B., Guo M., Zhang Z., Liu L., Ma H., Chen Z., Luo Y., Huang S., Chen L. CaMKII este implicat în cadmiu activarea căilor MAPK și mTOR care conduc la moartea celulelor neuronale. J. Neurochem. 2011; 119 :1108–1118. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]69.
Chan AS, Lai FP, Lo RK, Voyno-Yasenetskaya TA, Stanbridge EJ, Wong YH Receptorii melatoninei mt1 și MT2 stimulează kinaza N -terminală c-Jun prin proteinele G sensibile și insensibile la toxina pertussis. Celulă. Semnal. 2002; 14 :249–257. [ PubMed ] [ Google Scholar ]70.
Chen L., Xu B., Liu L., Luo Y., Yin J., Zhou H., Chen W., Shen T., Han X., Huang S. Peroxidul de hidrogen inhibă semnalizarea mTOR prin activarea AMPKalfa conducând la apoptoza celulelor neuronale. Lab Invest. 2010; 90 :762–773. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]71.
Chen L., Xu B., Liu L., Luo Y., Zhou H., Chen W., Shen T., Han X., Kontos CD, Huang S. Inducerea cadmiului a speciilor reactive de oxigen activează calea mTOR , ceea ce duce la moartea celulelor neuronale. Radic liber. Biol. Med. 2011; 50 :624–632. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]72.
Xu B., Chen S., Luo Y., Chen Z., Liu L., Zhou H., Chen W., Shen T., Han X., Chen L., Huang S. Semnalizarea calciului este implicată în apoptoza neuronală indusă de cadmiu prin inducerea speciilor reactive de oxigen și activarea rețelei MAPK/mTOR. Plus unu. 2011; 6 :e19052. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]73.
Selkoe DJ Boala Alzheimer este o insuficiență sinaptică. Ştiinţă. 2002; 298 :789–791. [ PubMed ] [ Google Scholar ]74.
Selkoe DJ Biologia celulară a proteinei precursoare de beta-amiloid și a presenilinei în boala Alzheimer. Trends Cell Biol. 1998; 8 :447–453. [ PubMed ] [ Google Scholar ]75.
Fisher A., Pittel Z., Haring R., Bar-Ner N., Kliger-Spatz M., Natan N., Egozi I., Sonego H., Marcovitch I., Brandeis R. M1 agoniştii muscarinici pot modula unele dintre semnele distinctive ale bolii Alzheimer: Implicații în terapia viitoare. J. Mol. Neurosci. 2003; 20 :349–356. [ PubMed ] [ Google Scholar ]76.
Lahiri DK Melatonina afectează metabolismul proteinei precursoare beta-amiloid în diferite tipuri de celule. J. Pineal Res. 1999; 26 :137–146. [ PubMed ] [ Google Scholar ]77.
Matsubara E., Bryant-Thomas T., Pacheco Quinto J., Henry TL, Poeggeler B., Herbert D., Cruz-Sanchez F., Chyan YJ, Smith MA, Perry G. și colab. Melatonina crește supraviețuirea și inhibă patologia oxidativă și amiloidă într-un model transgenic al bolii Alzheimer. J. Neurochem. 2003; 85 :1101–1108. [ PubMed ] [ Google Scholar ]78.
Lahiri DK, Chen D., Ge YW, Bondy SC, Sharman EH Suplimentarea alimentară cu melatonină reduce nivelurile de beta-peptide amiloide în cortexul cerebral murin. J. Pineal Res. 2004; 36 :224–231. [ PubMed ] [ Google Scholar ]79.
Song W., Lahiri DK Melatonina modifică metabolismul proteinei precursoare de beta-amiloid în linia celulară neuroendocrină PC12. J. Mol. Neurosci. 1997; 9 :75–92. [ PubMed ] [ Google Scholar ]80.
Zhang YC, Wang ZF, Wang Q., Wang YP, Wang JZ Melatonina atenuează inhibarea indusă de beta-amiloid a expresiei neurofilamentului. Acta Pharmacol. Păcat. 2004; 25 :447–451. [ PubMed ] [ Google Scholar ]81.
Olivieri G., Hess C., Savaskan E., Ly C., Meier F., Baysang G., Brockhaus M., Muller-Spahn F. Melatonina protejează celulele neuroblastomului SHSY5Y de stresul oxidativ indus de cobalt, neurotoxicitatea și creșterea secretia de beta-amiloid. J. Pineal Res. 2001; 31 :320–325. [ PubMed ] [ Google Scholar ]82.
Wang XC, Zhang YC, Chatterjie N., Grundke-Iqbal I., Iqbal K., Wang JZ Efectul melatoninei și melatonilvalpromidei asupra beta-amiloidului și neurofilamentelor în celulele N2a. Neurochema. Res. 2008; 33 :1138–1144. [ PubMed ] [ Google Scholar ]83.
Quinn J., Kulhanek D., Nowlin J., Jones R., Pratico D., Rokach J., Stackman R. Terapia cronică cu melatonină nu reușește să modifice încărcarea amiloidului sau deteriorarea oxidativă la șoarecii vechi Tg2576: Implicații pentru studiile clinice. Brain Res. 2005; 1037 :209–213. [ PubMed ] [ Google Scholar ]84.
Hsiao K., Chapman P., Nilsen S., Eckman C., Harigaya Y., Younkin S., Yang F., Cole G. Deficiențe de memorie corelative, creșterea Abeta și plăci de amiloid la șoarecii transgenici. Ştiinţă. 1996; 274 :99–102. [ PubMed ] [ Google Scholar ]85.
Su Y., Ryder J., Li B., Wu X., Fox N., Solenberg P., Brune K., Paul S., Zhou Y., Liu F. și colab. Litiul, un medicament comun pentru tratamentul tulburării bipolare, reglează procesarea proteinei precursoare de amiloid-beta. Biochimie. 2004; 43 :6899–6908. [ PubMed ] [ Google Scholar ]86.
Ryder J., Su Y., Liu F., Li B., Zhou Y., Ni B. Roluri divergente ale GSK-3 și CDK5 în procesarea APP. Biochim. Biophys. Res. comun. 2003; 312 :922–929. [ PubMed ] [ Google Scholar ]87.
Phiel CJ, Wilson CA, Lee VM, Klein PS GSK-3alpha reglează producția de peptide amiloid-beta ale bolii Alzheimer. Natură. 2003; 423 :435–439. [ PubMed ] [ Google Scholar ]88.
Donnelly PS, Caragounis A., Du T., Laughton KM, Volitakis I., Cherny RA, Sharples RA, Hill AF, Li QX, Masters CL și colab. Eliberarea intracelulară selectivă a ionilor de cupru și zinc din complecșii bis (thiosemicarbazonato) reduce nivelul peptidei amiloid-beta bolii Alzheimer. J. Biol. Chim. 2008; 283 :4568–4577. [ PubMed ] [ Google Scholar ]89.
White AR, Du T., Laughton KM, Volitakis I., Sharples RA, Xilinas ME, Hoke DE, Holsinger RM, Evin G., Cherny RA, et al. Degradarea beta-peptidei amiloid din boala Alzheimer prin reglarea în sus a activității metaloproteazei dependentă de metal. J. Biol. Chim. 2006; 281 :17670–17680. [ PubMed ] [ Google Scholar ]90.
Tesco G., Tanzi RE GSK-3 beta formează un complex tetrameric cu PS1-CTF/NTF endogen și beta-catenina. Efectele mutațiilor legate de D257/D385A și FAD. Ann. NY Acad. Sci. 2000; 920 :227–232. [ PubMed ] [ Google Scholar ]91.
Takashima A., Murayama M., Murayama O., Kohno T., Honda T., Yasutake K., Nihonmatsu N., Mercken M., Yamaguchi H., Sugihara S., et al. Presenilina 1 se asociază cu glicogen sintaza kinaza-3beta și substratul său tau. Proc. Natl. Acad. Sci. STATELE UNITE ALE AMERICII. 1998; 95 :9637–9641. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]92.
Simmons LK, May PC, Tomaselli KJ, Rydel RE, Fuson KS, Brigham EF, Wright S., Lieberburg I., Becker GW, Brems DN și colab. Structura secundară a peptidei beta-amiloid se corelează cu activitatea neurotoxică in vitro . Mol. Pharmacol. 1994; 45 :373–379. [ PubMed ] [ Google Scholar ]93.
Soto C., Castano EM Conformația peptidei beta Alzheimer determină viteza de formare a amiloidului și rezistența acestuia la proteoliză. Biochim. J. 1996; 314 :701–707. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]94.
Poeggeler B., Miravalle L., Zagorski MG, Wisniewski T., Chyan YJ, Zhang Y., Shao H., Bryant-Thomas T., Vidal R., Frangione B. și colab. Melatonina inversează activitatea profibrilogenă a apolipoproteinei E4 asupra peptidei amiloid Abeta Alzheimer. Biochimie. 2001; 40 :14995–5001. [ PubMed ] [ Google Scholar ]95.
Skribanek Z., Balaspiri L., Mak M. Interacțiunea dintre amiloid-beta-peptidă sintetică (1–40) și inhibitorii săi de agregare studiați prin spectrometrie de masă cu ionizare electrospray. J. Spectrom de masă. 2001; 36 :1226–1229. [ PubMed ] [ Google Scholar ]96.
Pappolla M., Bozner P., Soto C., Shao H., Robakis NK, Zagorski M., Frangione B., Ghiso J. Inhibirea beta-fibrilogenezei Alzheimer de către melatonină. J. Biol. Chim. 1998; 273 :7185–7188. [ PubMed ] [ Google Scholar ]97.
Huang TH, Fraser PE, Chakrabartty A. Fibrillogenesis of Alzheimer’s Abeta peptides studied by fluorescence energy transfer. J. Mol. Biol. 1997; 269 :214–224. [ PubMed ] [ Google Scholar ]98.
Fraser PE, Nguyen JT, Surewicz WK, Kirschner DA tranziții structurale dependente de pH ale peptidelor amiloide Alzheimer. Biophys. J. 1991; 60 :1190–1201. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]99.
Masilamoni JG, Jesudason EP, Dhandayuthapani S., Ashok BS, Vignesh S., Jebaraj WC, Paul SF, Jayakumar R. Rolul neuroprotector al melatoninei împotriva șoarecilor injectați cu peptida beta amiloid. Radic liber. Res. 2008; 42 :661–673. [ PubMed ] [ Google Scholar ]100.
Pacchierotti C., Iapichino S., Bossini L., Pieraccini F., Castrogiovanni P. Melatonin in psychiatric disorders: A review on the melatonin involvement in psychiatry. Față. Neuroendocrină. 2001; 22 :18–32. [ PubMed ] [ Google Scholar ]101.
Yuan H., Pang SF [125I] Situri de legare a iodomelatoninei în creierul porumbeilor: caracteristici de legare, distribuție regională și variație diurnă. J. Endocrinol. 1991; 128 :475–482. [ PubMed ] [ Google Scholar ]102.
Bieschke J., Zhang Q., Powers ET, Lerner RA, Kelly JW Metaboliții oxidativi accelerează amiloidogeneza Alzheimer printr-un mecanism în două etape, eliminând cerința de nucleare. Biochimie. 2005; 44 :4977–4983. [ PubMed ] [ Google Scholar ]103.
Feng Z., Zhang JT Efectul protector al melatoninei asupra apoptozei induse de beta-amiloid în celulele C6 de astrogliom de șobolan și mecanismul acestuia. Radic liber. Biol. Med. 2004; 37 :1790–1801. [ PubMed ] [ Google Scholar ]104.
Zatta P., Tognon G., Carampin P. Melatonina previne formarea radicalilor liberi datorită interacțiunii dintre peptidele beta-amiloide și metalioni [Al(III), Zn(II), Cu(II), Mn(II), Fe(II)] J. Pineal Res. 2003; 35 :98–103. [ PubMed ] [ Google Scholar ]105.
Feng Z., Chang Y., Cheng Y., Zhang BL, Qu ZW, Qin C., Zhang JT Melatonina atenuează deficitele comportamentale asociate cu apoptoza și disfuncția sistemului colinergic în modelul de șoarece transgenic APP 695 al bolii Alzheimer. J. Pineal Res. 2004; 37 :129–136. [ PubMed ] [ Google Scholar ]106.
Shen YX, Xu SY, Wei W., Sun XX, Liu LH, Yang J., Dong C. Efectele protectoare ale melatoninei de deteriorarea oxidativă indusă de beta-peptida amiloid 25–35 la șobolanii de vârstă mijlocie. J. Pineal Res. 2002; 32 :85–89. [ PubMed ] [ Google Scholar ]107.
Rosales-Corral S., Tan DX, Reiter RJ, Valdivia-Velazquez M., Martinez-Barboza G., Acosta-Martinez JP, Ortiz GG Melatonina administrată oral reduce stresul oxidativ și citokinele proinflamatorii induse de peptida beta-amiloid la șobolan creier: un studiu comparativ, in vivo versus vitamina C și E. J. Pineal Res. 2003; 35 :80–84. [ PubMed ] [ Google Scholar ]108.
Slats D., Claassen JA, Verbeek MM, Overeem S. Interacțiuni reciproce între somn, ritmuri circadiene și boala Alzheimer: Focus pe rolul hipocretinei și melatoninei. Imbatranire Res. Rev. 2013; 12 :188–200. [ PubMed ] [ Google Scholar ]109.
Rothman SM, Mattson MP Tulburări de somn în bolile Alzheimer și Parkinson. Neuromolecular Med. 2012; 14 :194–204. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]110.
Cecon E., Markus RP Relevanța acțiunilor cronobiologice și non-cronobiologice ale melatoninei pentru creșterea eficacității terapeutice în tulburările neurodegenerative. Brevetul recent. Endocr. Metab. Drug Imun Discov. 2011; 5 :91–99. [ PubMed ] [ Google Scholar ]111.
Kang JE, Lim MM, Bateman RJ, Lee JJ, Smyth LP, Cirrito JR, Fujiki N., Nishino S., Holtzman DM Dinamica beta-amiloid este reglată de orexină și de ciclul somn-veghe. Ştiinţă. 2009; 326 :1005–1007. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]112.
Coyle JT, Price DL, DeLong MR Boala Alzheimer: O tulburare a inervației colinergice corticale. Ştiinţă. 1983; 219 :1184–1190. [ PubMed ] [ Google Scholar ]114.
Rasool CG, Svendsen CN, Selkoe DJ Degenerarea neurofibrilară a neuronilor colinergici și noncolinergici ai creierului anterior bazal în boala Alzheimer. Ann. Neurol. 1986; 20 :482–488. [ PubMed ] [ Google Scholar ]115.
Samuel W., Masliah E., Hill LR, Butters N., Terry R. Hipocampal connectivity and Alzheimer’s dementa: Effects of synapse loss and tangle frequency in a two-component model. Neurologie. 1994; 44 :2081–2088. [ PubMed ] [ Google Scholar ]116.
Davis KL, Mohs RC, Marin D., Purohit DP, Perl DP, Lantz M., Austin G., Haroutunian V. Markeri colinergici la pacienții vârstnici cu semne precoce ale bolii Alzheimer. J. Am. Med. conf. univ. 1999; 281 :1401–1406. [ PubMed ] [ Google Scholar ]117.
Rinne JO, Laine M., Hiltunen J., Erkinjuntti T. Luarea deciziilor semantice în AD probabilă timpurie: Un studiu de activare PET. Cogn. Brain Res. 2003; 18 :89–96. [ PubMed ] [ Google Scholar ]118.
Terry AV, Jr, Buccafusco JJ Ipoteza colinergică a vârstei și a deficitelor cognitive legate de boala Alzheimer: provocări recente și implicațiile lor pentru dezvoltarea de noi medicamente. J. Pharmacol. Exp. Acolo. 2003; 306 :821–827. [ PubMed ] [ Google Scholar ]119.
Spencer JP, Middleton LJ, Davies CH Investigarea eficacității inhibitorului de acetilcolinesterază, a donepezilului și a noilor agenți procognitivi pentru a induce oscilații gamma în feliile de hipocamp de șobolan. Neurofarmacologie. 2010; 59 :437–443. [ PubMed ] [ Google Scholar ]120.
Guermonprez L., Ducrocq C., Gaudry-Talarmain YM Inhibarea sintezei acetilcolinei și nitrarea tirozinei induse de peroxinitrit sunt prevenite diferențial de antioxidanți. Mol. Pharmacol. 2001; 60 :838–846. [ PubMed ] [ Google Scholar ]121.
Feng Z., Cheng Y., Zhang JT Efectele pe termen lung ale melatoninei sau 17 beta-estradiol asupra îmbunătățirii performanței memoriei spațiale la șobolani adulți cu deficiențe cognitive, ovariectomizate. J. Pineal Res. 2004; 37 :198–206. [ PubMed ] [ Google Scholar ]122.
Tang F., Nag S., Shiu SY, Pang SF Efectele melatoninei și extractului de Ginkgo biloba asupra pierderii memoriei și activităților colin acetiltransferazei în creierul șobolanilor infuzați intracerebroventricular cu beta-amiloid 1-40. Life Sci. 2002; 71 :2625–2631. [ PubMed ] [ Google Scholar ]123.
Agrawal R., Tyagi E., Shukla R., Nath C. Un studiu al receptorilor de insulină din creier, activitatea AChE și stresul oxidativ în modelul de șobolan al demenței induse de ICV STZ. Neurofarmacologie. 2009; 56 :779–787. [ PubMed ] [ Google Scholar ]124.
Hansen RA, Gartlehner G., Webb AP, Morgan LC, Moore CG, Jonas DE Eficacitatea și siguranța donepezilului, galantamina și rivastigminei pentru tratamentul bolii Alzheimer: o revizuire sistematică și meta-analiză. Clin. Interv. Îmbătrânire. 2008; 3 :211–225. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]125.
Cardinali DP, Furio AM, Brusco LI Aspecte clinice ale intervenției melatoninei în progresia bolii Alzheimer. Curr. Neurofarmacol. 2010; 8 :218–227. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]126.
Fernandez-Bachiller MI, Perez C., Campillo NE, Paez JA, Gonzalez-Munoz GC, Usan P., Garcia-Palomero E., Lopez MG, Villarroya M., Garcia AG, et al. Hibrizii tacrină-melatonină ca agenți multifuncționali pentru boala Alzheimer, cu proprietăți colinergice, antioxidante și neuroprotectoare. ChemMedChem. 2009; 4 :828–841. [ PubMed ] [ Google Scholar ]127.
Spuch C., Antequera D., Isabel Fernandez-Bachiller M., Isabel Rodriguez-Franco M., Carro E. Un nou hibrid tacrină-melatonină reduce sarcina de amiloid și deficitele comportamentale într-un model de șoarece al bolii Alzheimer. Neurotox. Res. 2010; 17 :421–431. [ PubMed ] [ Google Scholar ]128.
Arends YM, Duyckaerts C., Rozemuller JM, Eikelenboom P., Hauw JJ Microglia, amiloid și demență în boala Alzheimer. Un studiu corelativ. Neurobiol. Îmbătrânire. 2000; 21 :39–47. [ PubMed ] [ Google Scholar ]129.
Combadiere C., Feumi C., Raoul W., Keller N., Rodero M., Pezard A., Lavalette S., Houssier M., Jonet L., Picard E., et al. Acumularea de celule microglia subretiniene dependentă de CX3CR1 este asociată cu caracteristicile cardinale ale degenerescenței maculare legate de vârstă. J. Clin. Investi. 2007; 117 :2920–2928. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]130.
Streit WJ, Mrak RE, Griffin WS Microglia și neuroinflamație: O perspectivă patologică. J. Neuroinflamm. 2004; 1 doi: 10.1186/1742-2094-1-14. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]131.
Hardy JA, Higgins GA Boala Alzheimer: Ipoteza cascadei amiloide. Ştiinţă. 1992; 256 :184–185. [ PubMed ] [ Google Scholar ]132.
Stuchbury G., Munch G. Inflamația asociată cu Alzheimer, potențiale ținte de droguri și terapii viitoare. J. Neural. Transm. 2005; 112 :429–453. [ PubMed ] [ Google Scholar ]133.
Tuppo EE, Arias HR Rolul inflamației în boala Alzheimer. Int. J. Biochim. Celula B. 2005; 37 :289–305. [ PubMed ] [ Google Scholar ]134.
Park SY, Jin ML, Kim YH, Kim Y., Lee SJ Efecte antiinflamatorii ale turmeronului aromatic prin blocarea căilor de semnalizare NF-kappaB, JNK și p38 MAPK în microglia stimulată de beta-amiloid. Int. Imunofarmacol. 2012; 14 :13–20. [ PubMed ] [ Google Scholar ]135.
Chung SY, Han SH Melatonina atenuează neurodegenerarea hipocampului indusă de acid kainic și stresul oxidativ prin inhibarea microgliale. J. Pineal Res. 2003; 34 :95–102. [ PubMed ] [ Google Scholar ]136.
Mohan N., Sadeghi K., Reiter RJ, Meltz ML Neurohormonul melatonina inhibă citokinele, mitogenele și radiațiile ionizante induse de NF-kappa B. Biochem. Mol. Biol. Int. 1995; 37 :1063–1070. [ PubMed ] [ Google Scholar ]137.
Chuang JI, Mohan N., Meltz ML, Reiter RJ Efectul melatoninei asupra activității de legare a ADN-ului NF-kappa-B în splina șobolanului. Cell Biol. Int. 1996; 20 :687–692. [ PubMed ] [ Google Scholar ]138.
Lau WW, Ng JK, Lee MM, Chan AS, Wong YH Interleukin-6 semnalizarea autocrină mediază fosforilarea STAT3 Tyr(705) indusă de receptorul melatoninei MT(1/2). J. Pineal Res. 2012; 52 :477–489. [ PubMed ] [ Google Scholar ]139.
Shen Y., Zhang G., Liu L., Xu S. Efectele supresive ale melatoninei asupra activării gliale induse de beta amiloid în hipocampul șobolanului. Arc. Med. Res. 2007; 38 :284–290. [ PubMed ] [ Google Scholar ]140.
Grases F., Costa-Bauza A., Prieto RM Un rol potențial al inhibitorilor de cristalizare în tratamentul bolii Alzheimer. Med. Ipoteza. 2010; 74 :118–119. [ PubMed ] [ Google Scholar ]141.
Boala Alzheimer Wollen KA: Avantajele și dezavantajele terapiilor farmaceutice, nutriționale, botanice și stimulatoare, cu o discuție despre strategiile de tratament din perspectiva pacienților și a practicienilor. J. Clin. Acolo. 2010; 15 :223–244. [ PubMed ] [ Google Scholar ]142.
Dowling GA, Burr RL, van Someren EJ, Hubbard EM, Luxenberg JS, Mastick J., Cooper BA Melatonina și tratamentul cu lumină puternică pentru perturbarea activității de repaus la pacienții instituționalizați cu boala Alzheimer. J. Am. Geriatr. Soc. 2008; 56 :239–246. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]143.
Pappolla MA, Chyan YJ, Poeggeler B., Frangione B., Wilson G., Ghiso J., Reiter RJ O evaluare a proprietăților antioxidante și antiamiloidogene ale melatoninei: Implicații pentru boala Alzheimer. J. Neural. Transm. 2000; 107 :203–231. [ PubMed ] [ Google Scholar ]144.
Nagtegaal J., Smits M., Van Der Meer Y., Fischer-Steenvoorden M. Melatonin: Un studiu asupra reacțiilor adverse suspectate la medicamente. Somn-Trezire Res. Netherl. 1996; 7 :115–118. [ Google Scholar ]145.
Avery D., Lenz M., Landis C. Linii directoare pentru prescrierea melatoninei. Ann. Med. 1998; 30 :122–130. [ PubMed ] [ Google Scholar ]
Articole din
Jurnalul Internațional de Științe Moleculare sunt furnizate aici prin amabilitatea
Multidisciplinary Digital Publishing Institute (MDPI)
Esra Küpeli Akkol, 1 Hilal Bardakcı, 2 Çiğdem Yücel, 3 Gökçe Şeker Karatoprak, 4 Büşra Karpuz , 5 şi Haroon Khan6Afișați mai multe
Editor academic: Jayeeta Ghose
Abstract
Tulburările de somn, precum și tulburările de ritm somn-veghe, sunt simptome caracteristice ale bolii Alzheimer (AD) care pot conduce la celelalte semne clinice ale acestei boli neurodegenerative. Modificările structurale și fiziologice legate de vârstă ale creierului duc la modificări ale tiparelor de somn. Condiții precum AD afectează cortexul cerebral, procreierul bazal, locus coeruleus și hipotalamusul, schimbând astfel ciclul somn-veghe. De asemenea, tulburările de somn afectează negativ evoluția bolii. Deoarece calitatea somnului este importantă pentru buna funcționare a memoriei, somnul afectat este asociat cu probleme în zonele conexe ale creierului care joacă un rol cheie în funcțiile de învățare și memorie. Pe lângă medicamentele sintetice, utilizarea plantelor medicinale a devenit populară în tratamentul bolilor neurologice. curcuminoide, care se află într-o structură diarilheptanoidă, sunt componentele principale ale turmericului. Printre acestea, curcumina are multiple aplicații în regimurile de tratament ale diferitelor boli, cum ar fi bolile cardiovasculare, obezitatea, cancerul, bolile inflamatorii și îmbătrânirea. În plus, curcumina a fost raportată a fi eficientă în diferite tipuri de boli neurodegenerative. Studiile științifice au arătat exclusiv că curcumina duce la îmbunătățiri semnificative în procesul patologic al AD. Cu toate acestea, solubilitatea sa scăzută, prin urmare, biodisponibilitatea scăzută este principala limitare terapeutică a curcuminei. Deși studiile anterioare s-au concentrat pe diferite tipuri de nanoformulări avansate ale curcuminei, sunt necesare noi abordări pentru a rezolva problema solubilității. Această analiză rezumă datele științifice disponibile,
1. Introducere
Plantele au o istorie lungă de utilizare ca remediu pentru numeroase probleme de sănătate. Când ne uităm la tratamentele tradiționale, Curcuma longa L. (Zingiberaceae), cunoscută în mod obișnuit sub numele de turmeric, iese în prim-plan. S-a raportat că turmericul este folosit în special în Ayurveda și are importanță religioasă în hinduism [ 1 ]. Mai mult, este folosit mai ales ca condiment și agent de colorare alimentară în bucătăria din Asia de Sud-Est [ 2 ]. Curcumina, un compus natural izolat din C. longa , are multiple aplicații în tratamentul diferitelor boli precum boli cardiovasculare [ 3 ], boli hepatice [ 4 , 5 ], obezitate [ 6 , 7 ], cancer [8 ], boli inflamatorii [ 9 , 10 ] și îmbătrânire [ 11 , 12 ].
Pe lângă aceste aplicații și activități, curcumina a fost raportată a fi eficientă în multe boli neurodegenerative. Boala Alzheimer (AD) este una dintre bolile neurodegenerative și reprezintă un procent mare de decese umane la nivel mondial. Pe baza datelor epidemiologice prezentate, un grup de autorități a estimat că 24,3 milioane de oameni au AD astăzi, cu 4,6 milioane de cazuri noi de AD în fiecare an. Numărul persoanelor afectate se va dubla la fiecare 20 de ani până la 81,1 milioane până în 2040. Studiile au arătat că curcumina poate duce la multe îmbunătățiri ale procesului patologic al AD [ 13 .]. Mecanismele sale de acțiune pot fi clasificate ca (I) susținerea homeostaziei sistemului inflamator, (II) creșterea clearance-ului substanțelor toxice din creier, (III) eliminarea radicalilor liberi și inducerea elementelor antioxidante și (IV) legarea și limitarea agregării. a proteinelor pliate greșit [ 1 ]. Deși curcumina are o gamă largă de acțiuni terapeutice, solubilitatea și biodisponibilitatea sa scăzută limitează utilizarea sa în clinică. Formulări farmaceutice noi trebuiau dezvoltate pentru a rezolva parametrii farmacocinetici ai acesteia [ 13 ].
În plus față de efectele curcuminei asupra bolilor neurodegenerative, în special AD, s-a raportat că curcumina poate fi benefică și în gestionarea privării de somn (SD) și a consecințelor neurologice induse de SD [14 ] . SD sa dovedit a avea o influență negativă asupra funcțiilor fizice, psihologice și neurologice. Din păcate, problemele SD sunt în creștere în public în zilele noastre [ 15 ]. Efectele curcuminei asupra cortexului prefrontal medial (mPFC) [ 16 ], potențialul protector asupra arborilor dendritici de la scurtarea și scurgerea induse de SD cronică [ 14 ] și efectele tulburărilor de memorie induse de SD au fost demonstrate in vivo. studii [ 17 ].
Noi tratamente alternative pentru tulburările neurologice cauzate de AD sau SD, precum și formulări mai bune pentru a depăși limitările terapeutice ale curcuminei, sunt încă în curs de cercetare. Studiile recente de revizuire pe acest domeniu sunt, de asemenea, foarte importante și pot oferi cercetătorilor noi perspective.
2. Evaluarea somnului și a tulburărilor cognitive
Investigarea problemelor de somn și odihnă necesită efectuarea unui morman de tehnici detaliate de diagnostic strategic. Metodele subiective și obiective sunt cele mai preferate. Înregistrarea scalelor de maniere și somn este printre tehnicile de evaluare subiectivă. Polisomnografia și actigrafia sunt metodele obiective alese în primul rând.
Indiferent de beneficiile tehnicilor subiective, cum ar fi accesibilitatea și cheltuielile reduse, acestea pot determina rate uriașe de rezultate fals-pozitive și fals-negative care se amestecă în evaluarea clinică. Mai multe investigații au demonstrat inconsecvența dintre datele dobândite prin tehnici subiective și obiective. De exemplu, persoanele cu deficiență cognitivă ușoară (MCI) au prezentat o percepție greșită mai proeminentă a orei de început a somnului decât cei sănătoși analogi [ 18 ].
Actigrafia este o tehnică de încredere pentru a evalua calitatea și natura somnului, deși este mai puțin sensibilă decât polisomnografia. Utilizarea acestei tehnici este rezonabilă în evaluările ciclului circadian și diagnosticarea tulburării de mișcare periodică a membrelor. Beneficiile acestei metode obiective sunt rentabilitatea, ușurința în aplicare, grade mai scăzute de conflict cu calitatea somnului și șansa de a mapa întreaga activitate motrică a indivizilor pentru intervale mai lungi [19 ] .
Cu toate acestea, polisomnografia dă rezultate mai exacte în comparație cu actigrafia în recunoașterea diferitelor tulburări de somn (de exemplu, probleme de respirație, mișcare recurentă a membrelor și tulburare comportamentală a somnului cu mișcarea rapidă a ochilor (REM)), precum și modificări ale caracteristicilor somnului. Datorită taxelor mai mari și a necesității de a păstra pacienții într-un laborator de somn, practica sa este mai dificilă. În plus, atitudinile de somn, în special la adulții, cu recurență mai mare a modificărilor ritmului circadian și amânări în timpul zilei, nu au putut fi evaluate prin polisomnografie.
3. Modificări ale somnului în deficiențe cognitive ușoare
Deteriorarea memoriei, pe lângă vârstă și educație, fără a prezenta modificări funcționale critice, este caracteristica particulară a MCI. Oamenii de știință care studiază funcțiile cognitive au grupat MCI ca MCI amnestic și nonamnestic și au clasificat fiecare grup în domenii specifice și multiple în funcție de evaluările neuropsihologice. Această grupare este crucială, deoarece indivizii cu MCI amnestic cu domenii multiple prezintă o rată mai mare de progres al demenței [ 20 ]. Simptomele MCI sunt prezentate în Figura 1 .
figura 1Simptomele deficienței cognitive ușoare (MCI)
Constatările din studiile cu populație mare asociază MCI cu rate mai mari de tulburări de somn în comparație cu persoanele sănătoase de aceeași vârstă. Valoarea tulburărilor de somn la pacienții cu MCI se modifică de la 8,8% la 59% [ 21 ]. În plus, tulburările de bază ale somnului, cum ar fi problemele de respirație, mișcarea recurentă a membrelor și tulburările de comportament ale somnului REM sunt frecvent observate în MCI [ 22 ].
Corelația dintre tulburările de somn și incidența mai mare a progresiei MCI la AD a fost evidențiată de studii prospective de cohortă [ 23 ]. O meta-analiză a cercetărilor care compară ambele măsuri obiective ale somnului la pacienții cu MCI amnezic și nonamnezic, împreună cu adulții sănătoși, a arătat că ambele subtipuri MCI au prezentat un model alternant ciclic mai mare, care este o expresie a instabilității somnului și a latenței alungite a somnului împreună cu scăderea eficienței somnului și timpul total de somn. Pacienții cu MCI amnezic au exprimat un indice de excitare redus și timpul total de somn în comparație cu pacienții cu MCI nonamnezici. Nu a fost observată nicio diferență între grupurile evaluate în ceea ce privește timpul de trezire după debutul somnului, somnul cu unde lente, durata REM, REM (%) și sindromul mișcării recurente ale membrelor [19 ]]. Totuși, pacienții cu MCI demonstrează atitudini crescute în timpul nopții [ 24 ].
Performanța cognitivă cu MCI amnestic și indivizii sănătoși au fost comparate folosind teste neuropsihologice. Inițial, evaluările neuropsihologice și polisomnografice ale pacienților au fost efectuate timp de două zile înainte de somn și din urmă. Au fost efectuate teste de reamintire a perechilor de cuvinte, recunoaștere a faptelor și obiecte promițătoare, iar grupul de control a prezentat performanțe crescute de memorie după somn. Dimpotrivă, performanța memoriei nu a revenit la pacienții cu MCI amnestic; chiar și testul de reamintire a perechii de cuvinte a arătat recidiva memoriei. Pacienții cu MCI nonamnestici suferă și de tulburări de somn. Disfuncția memoriei și timpul prelungit de veghe după debutul somnului cu alte disfuncții (deficiență de atenție, inhibarea răspunsului, formarea conceptului și gândirea critică) au fost legate de discontinuitatea somnului [25] .]. Cercetările legate de caracteristicile somnului au arătat că pacienții cu MCI au scăzut somnul cu unde lente în comparație cu indivizii sănătoși [ 26 , 27 ] și o fragmentare crescută a somnului non-REM. În plus, s-au realizat diferențe ambivalente în ceea ce privește REM, timpul de trezire după debutul somnului, durata REM și calitatea somnului [ 27 ].
Evaluarea puterii EEG în timpul somnului non-REM și REM a descoperit, de asemenea, contraste între pacienții cu MCI amnestic și cei sănătoși. Inițial, a fost observată reducerea puterii benzilor de frecvență EEG delta și theta. Scădere similară a fost observată în numărătoarea rapidă a axului pe locațiile frontale. S-a descoperit corelația dintre scăderea puterii delta și theta împreună cu o scădere a performanței cognitive a somnului din urmă [ 27 ]. În plus, intervalul REM a fost diminuat cu 5% la pacienții cu MCI. Durata prelungită REM a îmbunătățit rapid performanța cognitivă [ 28 ].
4. Modificări ale somnului în demența bolii Alzheimer
În anii 1980 și 1990, studiile electroencefalografice la pacienții cu demență AD și la indivizi sănătoși au oferit date neprețuite despre tiparele de somn și tulburările neuropsihiatrice, cum ar fi depresia. Demența AD și voluntarii sănătoși au prezentat tulburări de somn similare [ 29 ]. Cu toate acestea, gravitatea schimbărilor a fost de obicei mai mare, iar somnul REM a avut diferențe explicite. La pacienții cu demență AD, somnul a fost mult deranjat noaptea, atenția a progresat, iar numărul și durata somnului au fost mult perturbate, rezultând o perioadă totală de somn mai scurtă, cu o calitate mai scăzută [30 ] . Ca o concluzie, la pacienți s-au observat raportul superior al somnului din stadiul 1 și 2 și raportul scăzut al somnului cu unde lente [ 31] .]. Interacțiunile posibile dintre tulburările de somn și AD au fost prezentate în Figura 2 .
Figura 2Interacțiuni posibile între tulburările de somn și boala Alzheimer.
În plus, demența AD este identificată cu scăderi ale perioadei de somn ușor N2 (etapa 2), de exemplu, fusurile de somn și complexele K, care devin produse inadecvat, cu magnitudini și prevalențe mai mici [31 ] . Se știe că semnificația demenței este deteriorată din cauza acestor modificări, iar dezvoltarea bolii face dificilă discriminarea N1 (stadiul 1) și N2 [ 30 , 32 ], deoarece somnul fără faze vizibile, văzut în AD din urmă ca nedeterminat nedeterminat. REM a fost aranjat [ 29 , 31 , 33 ].
În timpul procesului de progresie a AD, sistemul colinergic începe să se deterioreze, ducând astfel la modificări ale somnului REM asociate demenței. Degenerarea creierului anterior bazal în AD, induce scăderea activității corticale în timpul somnului REM [ 29 , 31 ]. În mod similar, cercetările cantitative EEG au indicat o putere mai mare în frecvențele teta și delta și o putere redusă în frecvențele alfa și beta în timpul somnului REM al pacienților cu AD. Numărul total de episoade de somn REM și latența somnului REM rămân totuși intacte [ 31 ].
Îmbunătățirea amintirilor prin somn se crede că se datorează reactivării datelor și transferului hipocampo-neocortical legat de oscilațiile exacte ale creierului în somn, cum ar fi oscilațiile lente, fusurile talamocorticale și ondulațiile hipocampale în timpul somnului cu unde lente. Mai mult, somnul cu unde lente afectează amintirile dependente de hipocamp (amintiri declarative/episodice), în timp ce cantitățile mari de somn REM afectează amintirile care nu depind de hipocamp, cum ar fi învățarea procedurală legată de circuitele fronto-striatale. Deși mecanismul este încă ipotetic, tulburările de respirație legate de somn (SRBD) au un impact asupra disfuncției neuronale prin hipoxie sau întrerupere a somnului sau ar putea ajuta la apariția sau agravarea leziunilor vasculare și/sau neurodegenerative ale trunchiului cerebral. În plus, în timpul somnului REM, Nivelurile de acetilcolină sunt crescute, ceea ce este un factor critic în consolidarea amintirilor procedurale. Degenerarea sistemului colinergic ajută la exprimarea simptomatică a AD.29 ].
Lucrările inițiale care au evaluat schema de somn la pacienții cu AD au fost efectuate fără clasificare în funcție de severitate. Cu toate acestea, au fost efectuate cercetări avansate pentru a caracteriza somnul și la pacienții cu demență AD în stadiu ușoară. În mod predominant, modificarea principală în diagramele de somn/veghe afișate a fost interferența considerabilă a stării de veghe în timpul somnului [ 30 ].
5. Tulburările de somn ca factori de risc pentru boala Alzheimer
În secolul al XX-lea, s-a susținut că tulburările de somn ar putea contribui la progresia demenței și ipoteza principală a fost inițial că aceste tulburări au apărut în tandem ca procesul de îmbătrânire. Cu toate acestea, această teorie s-a schimbat după mai multe analize de cohortă prospective ale pacienților cu demență cu istoric de probleme de somn au șanse mai mari de a dezvolta AD decât cei fără probleme de somn. Una dintre cele mai recente meta-analize a aprobat posibila contribuție a somnului slab și a tulburărilor de somn ca un prognostic al AD.
Studii de cohortă prospective recente efectuate în jurul anilor 2000 au arătat că tulburările și tulburările legate de somn ar putea fi legate de riscul crescut de tulburări cognitive și demență. Aceste studii au relevat diferite forme de tulburări de somn și, în general, au constatat că acestea sunt asociate cu progresia demenței, de exemplu, legătura dintre insomnie [ 34 ], somnolență crescută în timpul zilei [ 35 , 36 ], perioade de somn prelungite [ 37 , 38 ], somn mai scurt. perioada [ 37 , 39 , 40 ] și apariția tulburărilor cognitive și/sau a demenței. Ca rezumat, atât durata scurtă cât și cea lungă de somn ar putea cauza tulburări cognitive/demență [38 , 39 ].
Participanții au fost evaluați prin utilizarea metodelor subiective în ceea ce privește caracteristicile somnului sau evaluarea cognitivă la început, dar dezvăluind legătura dintre tulburările de somn și funcționarea cognitivă [ 35 , 41 ]. Mai târziu (începutul anilor 2000), studiile de cohortă cu un număr adecvat de participanți au oferit date mai precise despre diagnosticul demenței prin utilizarea unor scale standardizate pentru a evalua somnul, deși studiile care nu au fost urmărite mai puțin de cinci ani au evitat datele respectabile [42 ] .
În 2010, a fost publicată o literatură care dezvăluie relația dintre riscurile de afectare cognitivă și somnul mai mult de nouă ore pe noapte după 10 și 14 ani de monitorizare a pacienților. Pe de altă parte, utilizarea telefonului ca instrument de interpretare a funcțiilor cognitive a dus la chestionarea acestui studiu [ 43 ].
Tranah și colab. au demonstrat dezavantajele strategice din studiile publicate anterior în studiul lor detaliat despre evaluarea somnului și a activităților zilnice cu actigrafie a încheieturii mâinii efectuat pe 1.282 de femei sănătoase [44 ] . Însoțitorii au fost expuși la o baterie neuropsihologică și la teste de demență după o monitorizare de 4,9 ani. S-a ajuns la concluzia că ritmurile circadiene deteriorate au crescut incidența demenței și a MCI. Împreună cu instrumentele standardizate de screening a somnului, în studiile avansate au fost utilizate tehnici de actigrafie a încheieturii mâinii mai detaliate [ 45 ]. Comparația dintre simptomele demenței și MCI a fost prezentată în Figura 3 .
Figura 3Simptome de demență și MCI
Yaffe și colegii săi au demonstrat că femeile sănătoase cu probleme de respirație cu tulburări de somn au fost mai susceptibile de a dezvolta MCI sau demență decât femeile fără probleme de somn prin utilizarea [ 46 ]. Această afirmație a fost verificată prin monitorizarea unui număr mare de participanți pentru perioade mai lungi. În plus, Pase și colegii săi au descoperit că scăderea somnului REM în loc de somnul non-REM este, de asemenea, legată de demență [ 47 ].
Nu au existat astfel de studii care au continuat mai mult de 15 ani pentru a evalua asocierea tulburărilor de somn și a performanțelor cognitive, chiar dacă apariția și ameliorarea AD durează 10-15 ani. Cea mai timpurie cercetare care a durat 22,5 ani a fost efectuată pe pacienți finlandezi de cel puțin 65 de ani [ 37 ]. Performanța cognitivă diminuată a fost obținută la pacienții cu durate scurte (<7 ore/zi) și lungi (>8 ore/zi) de somn și calitate insuficientă a somnului. Pase și colegii au efectuat analize asemănătoare confirmând rezultatele cu Virta și colegii [ 47 ].
Date suplimentare au fost obținute prin mai multe urmăriri. De exemplu, este prezentată legătura dintre tulburările de somn și tulburările cognitive non-AD de origine vasculară [ 36 ]. Mai mult, Potvin și colegii au declarat asocieri timpurii de sex, performanțe cognitive și diferite tipuri de întreruperi ale somnului [ 39 ].
În urma demonstrării asocierii semnificative dintre performanța cognitivă și calitatea somnului, au fost efectuate investigații epidemiologice. Almondes și colegii săi au explicat că persoanele cu fond de insomnie au prezentat un risc mai mare de a dezvolta demență (; 95%CI 1,07–2,18) [ 48 ]. Într-o serie de cercetări autentice, care include 27 de studii cu 69.216 participanți, tulburările de somn, cu excepția insomniei, au arătat, de asemenea, un risc mai mare de a dezvolta AD (1,55 (95% CI: 1,25–1,93)), tulburări cognitive (1,65 (95% CI: 1,45–1,86) ), și AD preclinic (3,78 (IC 95%: 2,27–6,30)) [ 49]. În plus, au fost efectuate meta-analize pentru rolul diagnostic al tulburărilor de somn în frecvența tulburărilor cognitive. Shi și colegii au clasificat tulburările de somn ca insomnie, probleme de respirație, somnolență mai lungă în timpul zilei, tulburări de mișcare asociate etc. și le-au confruntat cu diferitele tipuri de performanță cognitivă diminuată, unul câte unul, folosind 18 lucrări între cele 12.926 de lucrări publicate anterior. Conform constatărilor, insomnia este asociată cu AD, iar dificultățile respiratorii legate de somn sunt asociate cu toate tipurile de demență, inclusiv cu AD și demența vasculară. [ 40 ]. Cu toate acestea, ar trebui efectuate cercetări suplimentare pentru afirmații mai semnificative.
Diagnosticul AD este o provocare, deși biomarkerii sunt utilizați în predicția bolii [ 50 ]. În acest caz, polisomnografia și puncția lombară au fost folosite pentru a investiga relația dintre somn și AD. Asocierea dintre respirația tulburată de somn și AD a fost evidențiată prin evaluarea biomarkerilor lichidului cefalorahidian [ 51 ]. Alte studii care evaluează PET cu imagistica amiloid și tau sunt în curs de desfășurare.
6. Mecanisme potențiale care leagă tulburările de somn și riscul de AD
Deși datele clinice au dezvăluit asocierea dintre caracteristicile somnului și AD, pentru a expune mecanismele care stau la baza, au fost efectuate mai multe studii in vivo și clinice. F-fluorodeoxiglucoza (FDG-PET), un biomarker care arată activitatea metabolică a creierului, este calculată în timpul stărilor de veghe și somn. Nivelurile s-au dovedit a fi identice în somnul REM și în stare de veghe; cu toate acestea, activitatea metabolică cerebrală a fost diminuată considerabil (aproximativ 40%) în timpul somnului cu unde lente [ 52 , 53 ]. S-a afirmat că această afectare este localizată în regiunile subcorticale și corticale și s-ar putea datora stării tăcute suprapolarizate a neuronilor în timpul somnului cu unde lente [ 53 ]. Descărcarea de amiloid- β (A β) în lichidul interstițial implică activitatea sinaptică [ 52 ], ceea ce este demonstrat și de alte experimente [ 54 , 55 ]. O stare de veghe mai lungă duce la o concentrație mai mare de A β și se observă o escaladare suplimentară în timpul privării acute de somn. Gradele A β atenuate se dezvoltă în somn exclusiv în timpul somnului cu unde lente, dimpotrivă [ 55 ].
Celulele gliale au un rol semnificativ în eliminarea metaboliților toxici din creier și sunt numite sistem glimfatic și funcționează analog cu sistemul limfatic. Experimentele cu rozătoare au arătat că LCR este mutat în spațiile periarteriale ale arterelor cerebrale mai largi din zonele subarahnoidiene, mergând de-a lungul acestora pe măsură ce se ramifică în arterele penetrante. LCR este condus din spațiile perivasculare în interstițiul cerebral susținut pe picioarele de capăt ale astrocitelor de către transportorii AQP4. Un sistem similar se vede de la interstițiu până la spațiile perivenoase; LCR este transportat în spațiul subarahnoidian. Diverse căi produc un flux conjunctiv de lichid interstițial (ISF) de la spațiile periarteriale la cele perivenoase prin interstițiu, facilitând eliminarea metaboliților toxici acumulați în creier, cum ar fi A.proteina β și tau [ 56 , 57 ]. O investigație efectuată în 2012 a descoperit că o extensie de 60% a interstițiului are loc în timpul somnului, ceea ce ajută la comerțul convectiv între LCR și ISF, avansând un clearance -ul A β crescut în timpul somnului [ 58 ]. În concluzie, un transfer convectiv atenuat între ISF și LCR care duce la scăderea clearance-ului A β ar putea fi cauzat de tulburările de somn.
În total, ambele rezultate posibile ale perioadelor de somn perturbate (o performanță cerebrală extinsă cu niveluri crescute de A β și funcționare diminuată a schimburilor glimfatice cu eliminarea diminuată a A β ) ar putea determina niveluri mai mari de A β solubil în ISF, care este legat de amiloid. degradare [ 59 ]. Regiunile precuneus, parietal lateral și prefrontal medial ale creierului sunt implicate în sistemul de mod implicit, care este progresiv dinamic în perioadele de repaus, și sistemul cu cea mai mare activitate neuronală [60 ] . În plus, această rețea este regiunea cel mai ușor afectată de ansamblul A β în fazele inițiale ale AD.
Tulburările de ritm circadian sunt unul dintre potențialii factori de risc în neurodegenerare și AD. Studiile au demonstrat că reglarea învățării dependentă de hipocamp, neurogeneza hipocampală diminuată, învățare și memorie afectate și biomarkeri superiori legați de afecțiunile inflamatorii au apărut după anomalii ale ritmului circadian [ 61 ]. Mai târziu, Zhou și colegii au descoperit că degenerarea nucleului suprachiasmatic este cauza elementară a anomaliilor de ritm circadian la persoanele cu AD și sindromul apusului [ 62 ].
În plus, stresul oxidativ neuronal, degenerarea barierei hemato-encefalice și perturbarea neurogenezei hipocampului sunt câteva dintre celelalte caracteristici cauzate de tulburările de somn [ 63 ].
Plăcile de amiloid sunt compuse în regiuni ale creierului care controlează somnul, care perturbă și mai mult somnul. În mod similar, duce la mai multe simptome comportamentale cu demența AD, cum ar fi modificări ale stilului de viață, expunere redusă la lumina zilei, utilizarea polifarmaciei și scăderea calității somnului. Un alt motiv pentru problemele de somn care s-ar putea amesteca în mod legitim în patogeneza AD este respirația tulburată de somn, prin hipoxie și cascade inflamatorii.
7. Considerații privind tratamentul
Tratamentul nu trebuie inițiat conform simptomelor înainte de diagnosticarea etiologiei, deoarece simptomele și bolile pot interfera sau acoperi boli existente, cum ar fi delirul și demența la persoanele geriatrice. De exemplu, produsele farmaceutice utilizate în tratamentul infecțiilor acute ale tractului urinar la pacienții cu demență cu agitație nocturnă pot duce la întreruperi ale somnului. Mai mult decât atât, fazele incipiente ale delirului la pacienții geriatrici ar putea fi legate de o alimentație deficitară, constrângeri fizice, utilizarea cateterului vezical, polifarmacie sau condiții iatrogenice [ 64 ]. Aici, diagnosticul diferenţial corect joacă un rol important.
Tulburările de somn trebuie verificate prin polisomnografie înainte de tratament în cazul absenței oricărei comorbidități medicale. În cazul în care se observă caracteristici comportamentale anterioare seara sau nopții și orice membru al familiei ar putea fi martor la situație, ar trebui examinat sindromul picioarelor neliniştite (RLS). Examinările neuroimagistice au arătat scăderea nivelurilor de dopamină striatală pe tomografia cu emisie de pozitroni la pacienții cu AD rătăcitor cu RLS [ 65 ]. În cazul RLS, medicamentele agoniste dopaminergice ar putea fi preferate. Cu toate acestea, diagnosticul ar trebui să fie precis și ar trebui testat prin măsurarea nivelurilor serice de feritină, care este mai scăzută la pacienții cu demență și RLS [ 66 ].
Caracteristicile comportamentale nocturne trebuie evaluate dacă diagnosticul tulburărilor de somn eșuează. Triazolam (medicament de tip benzodiazepină) a fost testat într-un studiu clinic la scară mică și a arătat rezultate favorabile [ 67 ]. Alegerea agoniştilor GABA A în tratamentul AD este discutabilă; există studii cu rezultate controversate [ 68 ]. Utilizarea zolpidemului în ameliorarea tulburărilor de somn, a insomniei și a rătăcirii la pacienții cu demență și boala Parkinson sa dovedit a fi eficientă în studiile anterioare [69 – 71 ] . Cu toate acestea, astăzi, mulți geriatrici nu sunt dispuși să folosească sedative-hipnotice. În principal, sedative-hipnoticele cu timpi de înjumătățire prin eliminare lungi nu sunt preferate în special, deoarece duc la mai multe efecte adverse.72 ]. Psihotropul nocturn ar putea fi diminuat fără probleme [ 73 , 74 ]. Problemele de personal au un loc esențial în dozarea PRN, ca și factorii legați de pacient [ 75 ] și o literatură de lungă durată care leagă psihotropul și căderile și fracturile de șold [ 76 , 77 ]. Zolpidem cu un timp de înjumătățire de 2,5 ore poate determina pacienții să scadă [ 78 ]. Întrucât, nu există dovezi precise care să arate asocierea dintre utilizarea sedativ-hipnotică și căderi [ 76 , 77 , 79 , 80]. Comportamentul general al indivizilor cu probleme de somn este de a folosi sedative-hipnotice; cu toate acestea, se trezesc adesea în timpul nopții în afară de medicamente. Prin urmare, căderile ar putea fi legate de somn prost în loc de medicamente. Brassington și colegii săi au dezvăluit că întreruperile somnului sunt cauza căderilor în comunitățile mai în vârstă, demonstrând această ipoteză. [ 81 ].
Prescripția inhibitorilor de colinesterază (donepezil, galantamina, rivastigmină etc.) este frecvent preferată în tratamentul AD, iar aceștia afectează și somnul. Există o legătură între somnul REM și sistemul colinergic; prin urmare, durata REM mai lungă ar putea fi observată la pacienții care utilizează inhibitori de colinesterază. Această ipoteză este confirmată de alte studii; pacienti nedementatiau folosit donepezil și indivizii sănătoși care au folosit galantamina au prezentat modificări omoloage în duratele REM [ 82 , 83 ]. Densitatea mișcărilor oculare în timpul REM la persoanele sănătoase a fost crescută prin utilizarea rivastigminei [ 84 ]. Este incert dacă astfel de impacturi polisomnografice se pot întâmpla, de asemenea, în AD. Se poate ghici că, având în vedere semnificația pentru perioada REM și pentru învățare și memorie, s-ar putea identifica creșteri ale REM.
Există rapoarte de cazuri de vise izbitoare și coșmaruri la pacienții cu AD care primesc inhibitori de colinesterază care susțin această afirmație [ 85 , 86 ]. Indiferent dacă astfel de întâlniri se reflectă suplimentar în cele anunțate frecvent, impactul nefavorabil zilnic al privării de somn este neclar, dar ar apărea în orice caz. În acest context, frecvența insomniei a fost examinată prin utilizarea diferitelor doze de donepezil; cu toate acestea, atât rivastigmina, cât și galantamina s-au dovedit a avea efecte mai mici asupra somnului [ 87 – 90] .]. Datele din farmacosupravegherea arată că utilizarea donepezilului în AD este asociată cu o rată de peste două ori mai mare de prescripții sedative-hipnotice în comparație cu pacienții cu AD care nu iau donepezil. Acest lucru se adaugă la numărul tot mai mare de dovezi că inhibitorii de colinesterază sunt susceptibili de a perturba somnul la pacienții cu AD, în ciuda utilizării lor pe scară largă. [ 91]. În ciuda faptului că rivastigmina și galantamina nu perturbă somnul, există dovezi limitate că inhibitorii colinesterazei îmbunătățesc somnul la pacienții cu Alzheimer. Pe acest fundal de dovezi care sugerează că inhibitorii de colinesterază pot avea o utilitate limitată ca medicamente explicite pentru îmbunătățirea fomentării pe timp de noapte și a somnului în boala Alzheimer, câteva rapoarte recomandă utilizarea lor în tratamentul comportamentelor de interpretare a viselor asociate cu demența cu corp Lewy (LBD) [ 92 , 93 ]. Această informație este persistentă cu pierderi colinergice corticale frecvente în LBD, care este la un moment dat, peste cea observată în AD.
8. Chimia curcuminei
Structura compușilor care apar într-un amestec numit curcuminoide, care sunt principalele componente ale turmericului, constituie de obicei aproximativ 1-6% din plantă în greutate uscată și se află în structura diarilheptanoidă. Trei compuși majori formează curcuminoizi: curcumina (1E,6E)-1,7-bis(4-hidroxi-3-metoxifenil)-1,6-heptadien-3,5-dion, demetoxicurcumină și bisdemetoxi curcumină [94 ] . Formula sa chimică este C 21 H 20 O 6 și greutatea moleculară este de 368,38 g/mol. Două sisteme inelare aromatice care conțin grupări o-metoxi fenolice, conectate printr-o legătură cu șapte atomi de carbon, inclusiv un β α , β -nesaturat-dicetonă, se formează ca trei entități chimice în structura curcuminei [ 95 ]. Transferul de atomi de hidrogen intramolecular între gruparea β -dicetonă duce la prezența formei enol-tautomer și prezintă o solubilitate mai bună în dimetil sulfoxid și cloroform, solubilitate ușoară în metanol și solubilitate limitată în apă. În condiții acide și neutre, componenta majoră se află în formă de dicetonă, acționând ca donor de protoni, în timp ce în stare alcalină, forma ceto-enol acționează ca donor de electroni ca component principal (Figura 4) [ 96 ] . Curcumina are trei protoni ionizabili în apă: protoni enolici cu aproximativ 8,5 pKa și doi protoni fenolici cu 10–10,5 pKa (în amestec de solvent alcool/apă) [ 97]. Curcumina prezintă o absorbție puternică din punct de vedere spectrofotometric, variind de la 408 la 434 nm [ 96 ].
Figura 4Curcumina: dicetona și ceto-enolul formează echilibru
Primele studii asupra biosintezei curcuminoidelor au arătat că acestea au fost sintetizate din calea fenilpropanoidelor. Calea de biosinteză a curcuminei începe cu fenilalanină și implică 2 unități de acid cinamic și un carbon central din acidul malonic și următoarea adăugare de grupări funcționale pe inelele aromatice. Fenilalanina amoniac liaza este prima enzimă din calea de biosinteză. Curcumin sintaza cu trei izomeri (CURS1,2,3) și diketid-CoA sintază (DCS) și enzimele poliketid sintază au fost izolate din C. longa. DCS este enzima care formează feruloil-dietida-CoA prin catalizarea reacției feruloil-CoA cu malonil-CoA. Curcumin sintaza are loc în reacția de condensare decarboxilativă în biosinteza curcuminoidelor. Atât DCS, cât și CURS1, 2 și 3 au substraturi inițiale pentru a fi convertite într-unul dintre curcuminoizi. Unitatea de pornire feruloil-CoA se transformă în curcumină prin CURS. CURS catalizează, de asemenea, conversia p-cumaroildiketide-CoA și feruloil-CoA pentru a da demetoxicurcumină. Enzimele p -cumaroil shikimat transferaza, p -cumaroil quinat transferaza, acidul cafeic O -metiltransferaza și cafeoil-CoA O -metil-transferaza au fost, de asemenea, implicate în calea biosintetică a curcuminoidelor (Figura 5 ) [98 ].
Figura 5Calea biosintetică a curcuminoidului [ 98 ]
9. Noi sisteme de livrare utilizate pentru a crește biodisponibilitatea curcuminei
Au fost investigate sisteme de livrare nanodimensionate care oferă oportunități superioare în gestionarea bolilor sistemului nervos central (SNC), cum ar fi AD, care pot fi modificate cu agenți de țintire pentru a lega receptorii sau transportorii exprimați la bariera hemato-encefalică (BBB), crescând astfel selectivitatea SNC. și permeabilitate [ 99 ]. Se știe că compușii bioactivi naturali, cum ar fi curcumina, sunt eficienți în tratament de mulți ani, dar depășirea biodisponibilității lor scăzute este posibilă prin utilizarea sistemelor nanopurtători [ 100 , 101 ]. În studiile anterioare, sistemele de livrare nanodimensionate au fost dezvoltate cu curcumină, care sunt utilizate pe scară largă pentru a obține un rezultat terapeutic mai bun [ 102 , 103] .]. S-a raportat că curcumina posedă activitate neuroprotectoare în special asupra bolilor neurodegenerative precum AD, care progresează odată cu acumularea de peptidă β -amiloid în plăcile senile, care este cel mai important simptom histopatologic al bolii. S-a demonstrat că curcumina are posibilitatea de a încetini progresul AD prin reducerea β- amiloidului . În același timp, curcumina și-a luat locul în multe studii pentru a fi utilizată în tratamentul AD ca un compus proeminent cu efectul său antioxidant [ 104 , 105 ]. Utilizarea nanotehnologiei în sistemele de livrare a medicamentelor îmbunătățește biodisponibilitatea medicamentelor în sistemele biologice [ 106]. Datorită eliminării sistemice rapide a curcuminei, multe sisteme de livrare nanodimensionate au fost folosite pentru a crește timpul de circulație a curcuminei în organism pentru penetrarea curcuminei în diferite regiuni ale creierului [107 ] . S-a dovedit că curcumina nanoformulată a scăzut generația de A β in vitro și in vivo și, de asemenea, a inhibat agregarea depozitelor de A β [ 108 ].
Lipozomii (LP) sunt fosfolipide sferice cu două straturi, micro- sau nanodimensionate, biocompatibile și netoxice datorită asemănării cu membranele biologice și captează atât compuși hidrofobi cât și hidrofili. LP-urile au fost utilizate în domeniul farmaceutic de mulți ani ca nanopurtător pentru a servi potențialul terapeutic al agenților activi ca curcumină în tratamentul AD [ 99 , 109 ]. Lazar et. al. au proiectat nanolipozomi biocompatibili conjugați cu curcumină (CnLs) pentru țintirea depozitelor de amiloid. Datorită faptului că este legată covalent de suprafața CnLs, curcumina ar putea interacționa direct cu depozitele de amiloid. Au fost efectuate studii in vitro și au fost găsite CnL uniforme, stabile și netoxice; secretia de A β a fost reglata si A β-s-a prevenit toxicitatea indusa. CnL-urile au marcat puternic depozitele A β pe țesutul cerebral postmortem al pacienților cu AD și șoarecii transgenici care supraexprimă mutații umane legate de AD (șoareci APPxPS1) [ 110 ].
O altă formulare diferită de curcumină lipozomală a fost preparată de Taylor și colab. pentru a investiga agregarea peptidei amiloid- β 1-42 (A β 1-42) in vitro. Autorii au formulat diferite LP nanodimensionate (nLP) care încorporează sau decorate (prin chimie prin clic) cu curcumină, derivatul de curcumină, acid fosfatidic ligand lipidic, cardiolipină sau GM1. Toate nLP-urile preparate cu curcumină sau derivatul său au fost capabile să prevină formarea de fibrile și/sau oligomerice Aβ . Tipul click-curcumină, unul dintre cei trei lipozomi, a fost cel mai eficient și nLP-urile cu liganzi lipidici au inhibat doar formarea de fibrile și oligomeri A β la un raport LP/peptidă foarte mare. [ 111 ].
Mourtas şi colab. au dezvoltat și caracterizat nLP-uri multifuncționale, care conțin derivat de curcumină și decorate cu mAb (anti-transferin Ab) ca mediator de transport în LP. NLP-urile derivate de curcumină, cu și fără anti-TrF, au arătat o afinitate ridicată pentru plăcile senile de pe probele de creier postmortem ale pacienților cu AD, iar ambele nLP-uri au arătat că reduc agregarea peptidei A β (1-42 ) folosind testul tioflavine. Cu toate acestea, nLP-urile derivate de curcumină decorate cu mAbs au îmbunătățit semnificativ aportul de către modelul celular BBB in vitro [ 112 ].
Într-un alt studiu realizat de Mourtas și colegii de muncă, nLP-urile conjugate cu curcumină au fost formulate folosind diferite metode: metoda convențională și tehnica de chimie prin clic. NLP-urile care conțin derivatul de curcumină au arătat o afinitate mare pentru fibrilele A β (1-42) și cantități semnificative de depozite A β marcate în țesutul cerebral postmortem al pacienților cu AD. Prin urmare, curcumina neplanară nu a fost conectată la A β (1-42) din cauza apariției interacțiunilor multivalente. Injecția in vivo în hipocamp și neocortexul șoarecilor transgenici a arătat că supraexprimarea mutațiilor umane asociate AD (șoareci APP/PS1) și dezvoltarea unui număr mare de depozite A β au arătat că nLP-urile conjugate cu curcumină ar putea colora în mod specific A βdepozite. În cele din urmă, aceste nLP-uri conjugate cu curcumină pot fi preferate în diagnosticul sau tratamentul AD [ 113 ].
Mecanismele de acțiune ale curcuminei în AD au fost dovedite de multe studii (Figura 6 ) [ 114 – 117 ].
Figura 6Mecanismul de acțiune al nanoformulelor de curcumină în boala Alzheimer
Alte formulări lipozomale au fost formulate cu curcumină și conjugate cu un anticorp anti-receptor de transferină sau cu peptidă TAT care pătrunde în celule. Absorbția celulară crescută și permeabilitatea mai mare într-un model BBB au fost găsite în comparație cu LP-urile nedecorate [ 108 , 118 ].
Exozomii sunt nanovesicule naturale care sunt eliberate în mediu de aproape toate celulele, ale căror membrane sunt înconjurate de un dublu strat fosfolipidic (fosfolipide și colesterol), iar dimensiunile lor variază între 30 și 100 de nanometri. Conțin diverși acizi nucleici, lipide și proteine. Exozomii, un nanofarmaceutic natural cu potențialul de a fi utilizați în diagnostic și tratament în medicină, au multe caracteristici dezirabile ale sistemului ideal de livrare a medicamentelor. Exozomii sunt vezicule derivate din celule cu potențial terapeutic și biocompatibilitate ridicată. Propunând un nou concept de lipozomi asemănătoare exozomilor pentru administrarea de medicamente, Fernandes și colegii săi au selectat o moleculă model de curcumină și au examinat eficacitatea exolipozomului curcuminei împotriva stresului oxidativ și în scăderea β .-acumularea de amiloid în tratamentul AD. S-a raportat că lipozomii de tip nou produși cu o eficiență ridicată de încapsulare au fost necitotoxici pe celulele neuronale SH-SY5Y și au potențial de neuroprotecție. Exo-lipozomii încărcați cu curcumină au fost interiorizați, iar curcumina a fost eliberată în interiorul celulelor. Acest transportor inovator este o abordare nouă și eficientă pentru a furniza medicamente către creier, deoarece este stabil, păstrează încărcătura și este reținut de celulele neuronale [ 119 ].
Nanoparticulele (NP) sunt denumiți purtători coloidali în care medicamentele sunt încapsulate sau adsorbite la suprafață. Aceste sisteme de eliberare a medicamentelor asigură livrarea susținută și controlată a medicamentelor în BBB și o durată de viață crescută a circulației. Multe modificări cu polietilen glicol (PEG), peptide, polimeri sau anticorpi pot fi utilizate fie prin adsorbție, fie prin legarea chimică pe suprafața NP-urilor pentru a îmbunătăți livrarea medicamentului. Studiile recente indică faptul că livrarea de NP, în special de NP modificate, este mai eficientă în creier [ 99 , 120 ].
Mathew et al. aptamerul de legare de amiloid (NN2) preparat conjugat cu NP-uri de curcumină acoperite cu poli(acid lactic-co-glicolic) (PLGA) pentru a se lega de plăcile de amiloid și a preveni agregarea acestora. Ei au propus să utilizeze potențialul aptamerilor ca instrumente terapeutice noi care se atașează eficient la fibrilele de amiloid și să evalueze eficacitatea NP-urilor PLGA încapsulate în curcumină împotriva plăcilor de amiloid. S-a raportat că NP-urile NN2-curcumină-PLGA atașate cu proteinele amiloide și dezagregarea proteinelor amiloide au putut fi observate la NP-uri formulate cu și fără aptamer. S-a înțeles că activitatea antiamiloidă a aptamerului atașat nu s-a schimbat din imaginile SEM. Cu toate acestea, dimensiunea agregatului de proteine a fost scăzută atunci când curcumină-PLGA NP-uri conjugate cu aptamer și agregatele de amiloid au fost incubate împreună [ 121] .].
Într-un alt studiu, Cheng et al. a preparat o nanocurcumină foarte stabilă utilizând un polimer în bloc de polietilen glicol-acid polilactic (PEG-PLA) și polivinilpirolidonă (PVP). Această nouă formulare a transportat eficient curcumina prin BBB cu o creștere de aproximativ șase ori a timpului mediu de rezidență în creier, comparativ cu curcumina neformulată. De asemenea, s-a observat o memorie mai bună la șoarecii Tg2576 în testele de condiționare a fricii contextuale tratate cu nanocurcumină [ 122 ]. Fan și colegii de muncă au proiectat NP-uri PLGA-PEG încărcate cu curcumină conjugate cu peptida B6 și pentru a evalua potențiala eficacitate a PLGA-PEG-B6 care conține curcumină în celulele HT22 și șoarecii transgenici APP/PS1 Al pentru tratamentul AD. In vitroanalizele au fost testate și s-a indicat că NP-urile PLGA-PEG-B6 încărcate cu curcumină au un diametru îngust, absorbție celulară crescută și compatibilitate bună cu sânge. În plus, aceste noi NP ar putea avansa capacitatea de învățare și memorie spațială a șoarecilor APP/PS1 în comparație cu curcumina neformulată și au proprietăți promițătoare în ameliorarea amiloidozei și a tauopatiei cu celule b cu biodisponibilitate ridicată [123 ] .
Huang şi colab. studiat pe alte NP-uri PLGA care conțin peptida S1 ca inhibitor al generării A β și curcumină. Peptida ciclică mimică de fier (CRT) este capabilă să țintească complexul proteic al transferinei și receptorul transferinei (TfR), iar această conjugare oferă o penetrare crescută a BBB, timpul de înjumătățire in vivo și efecte terapeutice. Conform acestui studiu, s-a sugerat că, în comparație cu alte NP-uri PLGA, NP-urile conjugate cu CRT care conțin peptida S1 și curcumină au arătat o creștere a permeabilității BBB a NP-urilor PLGA și peptida S1 și curcumina a jucat un rol important în tratamentul șoarecilor AD prin scăderea generației A β [ 124 ].
Într-un studiu anterior, Meng și colab. a proiectat purtătorul de lipide nanostructurat (NLC) cu lipoproteine de densitate joasă încărcat cu curcumină, modificat cu lactoferină (Lf) pentru a viza creier și a evaluat efectul acestuia asupra progresiei AD la șobolani. Autorii au folosit diferite niveluri de Lf pentru modificarea NLC. In vitrocaracterizarea, absorbția și citotoxicitatea au fost măsurate în celulele endoteliale capilare ale creierului (BCEC). Conform studiului de eliberare, nu a fost observată nicio diferență semnificativă în ceea ce privește conținutul activ eliberat din NLC-mimic Lf preparat cu diferite niveluri de Lf. Dar, absorbția crescută a Lf-mNLC de către BCEC a fost găsită în mod semnificativ în comparație cu nivelul redus de Lf modificat de NLC, ceea ce dezvăluie procesul de endocitoză mediată de receptorul Lf. O acumulare și un control mai mare al progresiei AD au fost prezentate cu Lf-mNLC în cortex și al treilea ventricul decât cele ale NLC și este posibil să spunem că Lf-mNLC poate fi un nou nanopurtător modificat pentru țintirea creierului [125 ] .
Huo și colab. au proiectat nanoparticule de seleniu curcumină încapsulate nanosfere PLGA și au evaluat efectele acestora în reducerea agregării Ap în probe de creier ale șoarecilor cu AD. S-a demonstrat că nanosferele sintetizate încărcate cu curcumină au scăzut cantitatea de A β și au îmbunătățit deficiența de memorie. Nanosferele Se-PLGA încărcate cu curcumină au fost legate în mod specific de plăci A β și au inhibat eficient plăcile. Sa raportat că aceste nanosfere PLGA încapsulate în NP ar putea fi direcționate către plăcile de amiloid și ar putea fi o alternativă eficientă pentru a îmbunătăți eficiența terapeutică în leziunile AD [ 126 ].
Tiwari și colegii au formulat o nanoparticulă PLGA încapsulată în curcumină și au evaluat eficiența acesteia asupra neurogenezei și a deficiențelor cognitive. Autorii s-au concentrat pe activarea căii Wnt/ β -cateninei, translocarea nucleară a β -cateninei și nivelurile GSK-3 β care sunt legate de reglarea neurogenezei prin îmbunătățirea expresiei genelor proneurogene. Aceste NP au fost obținute care au îmbunătățit proliferarea NSC și diferențierea neuronală in vitro și au inversat tulburările de învățare și memorie în modelele de șobolan asemănător AD induse de beta amiloid, în comparație cu curcumina în vrac neformulată [120 ] .
Ca nou studiu, Wu a realizat un studiu comparativ prin colectarea tuturor datelor folosind cuvântul cheie Morris water labirint pentru a studia AD la rozătoare prin limitarea literaturii în general între anii 2010 și 2021 și evaluând învățarea și memoria spațială a rozătoarelor, folosind meta. -metoda de analiză. Nu a fost găsită nicio diferență semnificativă statistic în eficacitatea lor. S-a ajuns la concluzia că ambele nanoparticule sunt la fel de eficiente în tratamentul AD [ 127 ].
Într-un alt studiu, datele cercetării au arătat că nanoparticulele de albumină serică umană camuflate cu membrană de celule roșii din sânge (RBC) purtând molecule de T807 și trifenilfosfină (TPP) atașate la suprafața membranei RBC (T807/TPP-RBC-NPs) cu curcumina ca model antioxidant. nanosistemul a atenuat simptomele AD prin reducerea stresului oxidativ mitocondrial și suprimarea morții neuronale atât in vitro, cât și in vivo [ 128 ].
Nanoparticulele lipidice solide (SLN) încărcate cu curcumină au fost determinate de Kakkar și colab. pentru livrarea creierului la șobolani pe cale orală. Ei au sugerat că activitatea crescută a acetilcolinesterazei și concentrația de două ori a curcuminei în creier au fost găsite în comparație cu curcumina neformulată atunci când este administrată oral [ 129 ].
O altă abordare este NP-urile magnetice conjugate cu curcumină, care se leagă în mod specific de plăcile de amiloid. NP-urile magnetice conjugate cu curcumină biocompatibile acoperite cu copolimer bloc PEG-PLA și PVP care aveau un diametru mediu sub 100 nm și-au caracterizat structura prin metode spectroscopice. NP-urile magnetice conjugate cu curcumină au fost găsite netoxice prin rinichiul canin Madin-Darby sau prin celulele diferențiate ale neuroblastomului uman. S-a constatat că coeficientul de permeabilitate aparent al NPs este–6 cm/s într-un model BBB in vitro . In vivo , după injectarea de NP, s-au găsit plăci de amiloid în imagistica prin rezonanță magnetică (IRM) a creierului de șoarece Tg2576. Din punct de vedere imunohistochimic, examinarea creierului de șoarece a arătat că NP-urile au arătat o afinitate mare pentru plăcile de amiloid și s-a crezut că NP-urile ar putea fi de ajutor pentru diagnosticarea AD folosind RMN [122 ] .
Este posibilă direcționarea creierului, îmbunătățirea proprietăților farmacocinetice și farmacodinamice și reducerea efectelor secundare prin utilizarea nanopurtătorilor pe bază de lipide. Administrarea intranazală este o modalitate convenabilă de a traversa bariera hemato-encefalică pentru a ținti medicamentele către creier. Prin acest mod neinvaziv, ocolind BBB, metabolismul presistemic poate fi eliminat [ 130 ]. În acest moment, într-un studiu, nanovesiculele AmyloLipid (ALN) încărcate cu curcumină au fost fabricate pentru a gestiona AD prin inhibarea agregării A β și a inflamației induse de A β prin livrare nazală. A fost obținut după o oră de administrare intranazală a nanoparticulelor încărcate cu curcumină care au arătatnivelurile creierului şinivelurile plasmatice din studiul in vivo și se poate spune că un ALN bine conceput care conține curcumină administrat intranazal este o abordare promițătoare [ 131 ].
Miezul/învelișul NP PLGA încapsulat în chitosan cu curcumină, ale căror aplicații clinice sunt limitate din cauza solubilității scăzute și a biodisponibilității scăzute și a metabolismului de primă trecere în ciuda potențialului său terapeutic puternic, au fost produse și evaluate cu administrare intranazală de către Dhas și Metha. Ca rezultat al studiului de eliberare și permeabilitate, s-a observat o eliberare susținută și o permeabilitate îmbunătățită a mucoasei nazale. În studiul in vivo , s-a constatat că biodistribuția NP-urilor este mai mare în creier urmând calea intranazală, demonstrând un efect semnificativ de captare a ROS asupra activității antioxidante. Cu toate aceste date, s-a văzut că NP-urile au potențialul de a reduce stresul oxidativ din creier pentru un tratament eficient al AD [ 132 ].
Zhang et. al. a produs nanoparticule de poli (acid lactic-co-glicolic) acoperite cu chitosan (CUR-CS-PLGA-NPs) și complexe de incluziune hidroxipropil- β -ciclodextrină de curcumină, care este dificil de administrat din cauza solubilității slabe și a structurii instabile și a comparat acestea prin administrare intranazală. Studiile in vitro au arătat că complexele de incluziune au fost mai stabile în condiții fiziologice decât CS-PLGA-NP și s-a obținut o absorbție celulară mai mare a curcuminei în celulele SH-SY5Y. Ambele formulări nu au fost citotoxice celulare și au prezentat un efect antioxidant comparabil. In vivostudiile farmacocinetice au arătat că, după administrarea intranazală a aceleiași doze, valorile ASC ale CUR în plasmă și creier ale complexului de incluziune, care este atât o metodă simplă, cât și structuri adecvate pentru extinderea producției, au fost de 2,57 ori și 1,12 ori mai mari decât cele ale CS-PLGA-NP, respectiv [ 133 ]. Pe baza datelor acestui studiu și a altor studii cu administrare intranazală, se poate spune că calea intranazală este o alternativă eficientă și bună pentru livrarea curcuminei și a altor medicamente/terapeutice către creier sau SNC.
Microemulsiile (ME) și nanoemulsiile (NE) sunt stabile din punct de vedere termodinamic, iar formularea limpede din punct de vedere izotrop constă din faza uleioasă și faza apoasă stabilizată prin amestecul unui surfactant și cosurfactant. În ciuda avantajelor ME/NE, puține studii privind utilizarea studiilor NE au fost disponibile pentru livrarea creierului. Sood și colab. au dezvoltat sisteme NE și mucoadezive NE (mNE) pentru a furniza curcumină pentru tratamentul AD. Conform studiilor in vitro de citotoxicitate și ciliotoxicitate nazală, au fost dezvoltate formulări netoxice și sigure. Cel mai mare flux prin mucoasa nazală de oaie a fost observat cu mNE-uri în comparație cu NE și soluția de curcumină și s-a sugerat că curcumina slab solubilă ar putea fi administrată pentru livrarea nazală pentru îmbunătățirea captării creierului [134 ] .
Din nou, purtătorii de lipide nanostructurați (NCL) încărcați cu curcumină/donepezil au fost pregătiți și utilizați cu administrare intranazală pentru livrarea la creier. O concentrație mai mare de medicamente în creier a fost găsită cu aplicarea intranazală comparativ cu aplicarea intravenoasă. S-a folosit un model de șoarece și s-a obținut că memoria și învățarea au fost îmbunătățite în comparație cu grupul tratat cu medicamentul neformulat. De asemenea, nivelurile îmbunătățite de ACh și daunele de oxidare scăzute au fost găsite în grupurile tratate cu NLC [ 108 ].
Într-un alt studiu publicat, Mandal și colab. a avut ca scop proiectarea gelului ME mucoadeziv (mMEg) pentru a îmbunătăți absorbția creierului de curcumină pe cale intranazală. După dezvoltarea mMEg, au fost efectuate studii de caracterizare, stabilitate, mucoadeziune și nazociliotoxicitate și au fost obținute mMEg transparente, stabile și neciliotoxice. Absorbția creierului de curcumină pe cale nazală a fost determinată la șobolani. Când s-a utilizat curcumină mMEg, s-a obținut de 11 ori mai multă absorbție de curcumină în bulbul olfactiv comparativ cu injectarea intravenoasă a soluției de curcumină. După administrarea nazală de curcumină mMEg, rapoartele ASC au fost semnificativ mai mari decât cele după administrarea intravenoasă de curcumină liberă [ 135 ].
Într-un studiu nou, Sharma a raportat sistemul de livrare a medicamentelor nanorobot pentru a îmbunătăți biodisponibilitatea curcuminei în timpul tratamentului AD. Au fost examinate strategia de proiectare a sistemului de livrare a medicamentelor nanorobot cu o țintă specifică și formularea farmacocinetică a reacțiilor paralele concurente asociate. Cromoforul a fost folosit ca senzor, iar imagistica computerizată și designul de control al feedback-ului poate avea ca rezultat o biodisponibilitate mai mare pentru efectul terapeutic al curcuminei pentru tratarea AD [ 136 ].
Giacomeli et al. a proiectat nanocapsule cu miez lipidic (LNC) încărcate cu curcumină și a evaluat efectele neuroprotectoare asupra unui model de AD indus de injecții intracerebroventriculare cu peptida β -amiloid1-42 (A β1-42 ). Curcumină neformulată (50 mg/kg, po) și nanocapsule de curcumină (10 sau 1 mg/kg, po) au fost date șoarecilor femele în vârstă timp de 14 zile după administrarea A β 1-42. Tratamentul oral timp de 14 zile a redus atât nivelurile, cât și expresia proteinelor citokinelor inflamatorii, care joacă un rol critic în neurodegenerarea AD. LNC a arătat efect de neuroprotecție semnificativ împotriva A β1-42-inducție care sunt asociate cu niveluri crescute de citokine. S-a observat că efectul neuroprotector al curcuminei a crescut odată cu nanoîncapsularea sa și această nanocurcumină poate fi eficientă în prevenirea și tratamentul AD [ 137 ].
Când evaluăm datele tuturor studiilor anterioare, s-a văzut că nanoformulele de curcumină au ajutat la reducerea simptomelor și la tratarea AD. Avantajele nanoformulărilor au fost eficiente prin creșterea biodisponibilității scăzute, a absorbției creierului și a efectului terapeutic al curcuminei. Se poate spune că aceste nanoformulări sunt o alternativă care poate fi benefică în diagnosticul și tratamentul AD în viitor. De asemenea, se vede că nanoformulările cu diferite căi de aplicare ajută la depășirea problemei actuale de biodisponibilitate cu nanotehnologia în curs de dezvoltare, ținând cont de conformitatea pacientului. Cu toate acestea, nu trebuie ignorate posibilele efecte toxice ale nanotehnologiei, care se dezvoltă și oferă multe posibilități.
10. Rolul curcuminei în tulburările de somn și boala Alzheimer: informații din studiile in vitro și in vivo
Curcumina este prezis a fi un agent neuroprotector, datorită activităților sale antioxidante și antiinflamatorii puternice și, prin urmare, poate fi benefică în terapia diferitelor tulburări neurologice și afecțiunilor legate de stres, cum ar fi AD.
Kumar şi colab. a efectuat cercetări pentru a investiga funcția oxidului nitric în efectele preventive ale curcuminei împotriva modificărilor comportamentale ale privării de somn de 72 de ore și a daunelor oxidative la șoareci. Extract de curcumină aplicat la 10 și 20 mg/kg (ip) la șoareci timp de 5 zile. Tratamentul cu curcumina a restabilit glutationul epuizat, activitatea catalazei și a redus peroxidarea lipidelor și nivelurile de nitriți, prevenind pierderea în greutate, scăderea activității locomotorii și efecte similare anxietății în toate paradigmele comportamentale. În plus, inhibitorul sintazei de oxid nitric a contribuit într-un mod semnificativ la eficacitatea protectoare a curcuminei [ 15 ].
Deoarece alcoolismul are ca rezultat o varietate de tulburări neuronale, cum ar fi starea de conștiență, tulburări de somn, insomnie, demență și amnezie, efectele curcuminei asupra 5-hidroxi-triptaminei (5-HT) și 5-hidroxi-indolului induse de etanol. Au fost investigate nivelurile și ritmurile acidului acetic (5-HIAA) pe SCN și pineală. În grupul tratat cu curcumină, după consumul de etanol, s-a administrat 0,002% (ad libitum) curcumină timp de 15 zile. La șobolanii aplicați cu curcumină, a fost observată o reducere a cantității de 5-HT și 5-HIAA [ 138 ].
Noorafshan și colab. a efectuat o cercetare pentru a analiza efectele potențiale ale tratamentului cu curcumină asupra lipsei de somn cu mișcarea rapidă a ochilor (REM-SD) [ 16 ]. S-a determinat că volumul, numărul de celule, reconstrucția, timpul de recunoaștere a obiectului și greutatea corporală au fost recuperate îngrup comparativ cugrup de apă. Într-un studiu diferit, Noorafshan și colab. a examinat, de asemenea, influența curcuminei asupra cortexului prefrontal medial (mPFC) al privării de somn la șobolani. Reducerea semnificativă a modificărilor atrofice mPFC în pierderea celulară și modificările dendritice au fost cercetate la animalele SD după tratamentul cu curcumină [ 17 ].
În studiul în care a fost examinată potența curcuminei asupra microbiotei intestinale a șobolanilor cu privare de somn interval (ISD), 70 mg/kg curcumină a fost aplicată șobolanilor din grupul de tratament. Conform rezultatelor experimentelor comportamentale, s-a observat un progres semnificativ în grupul de tratament cu curcumină. Acest studiu a dovedit că ISD nu numai că duce la tulburări mintale la șobolani, dar dă naștere și la modificări ale Escherichia coli , Bifidobacterium , Lactobacillus , Clostridium perfringens și Bacteroides și curcumina a schimbat dezechilibrul microbiotei [ 139 ].
Nivelurile de antioxidanti, parametrii histopatologici și efectul curcuminei asupra leziunilor hepatice cu privarea de somn la șobolanii lipsiți de somn acut și cronic au fost investigate de Saliha și colab. Potrivit rezultatelor studiului, pretratamentul cu curcumină a arătat o îmbunătățire semnificativă în grupul acut lipsit de somn, în timp ce schimbările pozitive nu au fost semnificative în grupul cronic. Descoperirile biochimice sunt susținute și de constatările histopatologice. Studiul a raportat că privarea de somn poate duce la deteriorarea stresului oxidativ în ficat, iar aportul de curcumină alimentară poate crește nivelul de antioxidanți din organism și poate reduce riscurile cauzate de privarea de somn [140 ] .
Mai recent, Erfanizadeh et al. a investigat efectul privării cronice de somn asupra modificărilor histomorfometrice ale ganglionului cervical superior și a capacității curcuminei de a preveni aceste modificări. 36 de șobolani adulți masculi au fost repartizați aleatoriu în șase grupuri, inclusiv control, curcumină, lipsiți de somn și somn-, podea grilă și grilăgrupuri. Un ml de curcumină în doza de 100 mg/kg/zi (po) curcumină aplicată grupelor de curcumină timp de 21 de zile. În ceea ce privește descoperirile stereologice, CSD a scăzut semnificativ volumul SCG și numărul total de neuroni și celule gliale satelit în comparație cu grupul de control. Tratamentul CSD cu curcumină a prevenit reducerile și, de asemenea, evaluarea TUNEL a evidențiat apoptoză semnificativă în celulele SCG din grupul CSD, iar curcumina a redus semnificativ această apoptoză. O scădere a apoptozei a fost observată în toate grupurile de control aplicate cu curcumină. Drept urmare, s-a raportat că efectele nocive ale privării de somn pot fi prevenite prin administrarea orală de curcumină [ 141 ].
Deși există puține studii despre privarea de somn cu curcumină, cele care au fost făcute arată că, atunci când este administrată în timpul privării de somn, curcumina eficientă antioxidantă previne pierderea neuronilor de privare de somn în corteza șobolanului. Cu toate acestea, doar o singură cercetare a folosit metode stereologice pentru a evalua pierderea celulară [ 141 ]. Studiile au raportat că privarea de somn crește stresul oxidativ sau că curcumina a scăzut stresul oxidativ la animalele protejate de pierderea somnului leziuni neuronale.
Un studiu recent la scară largă constată că benzodiazepinele, care sunt adesea folosite pentru a trata anxietatea și problemele de somn, sunt asociate cu un risc crescut de boala Alzheimer la persoanele în vârstă [142 ] ; puține studii au raportat efecte asupra AD. În celulele granulare cerebeloase de șoarece, ramelteonul a îmbunătățit conținutul neuronal al BDNF. Prin urmare, s-a afirmat că, dacă aplicarea ramelteonului poate regla nivelul BDNF al creierului, acesta poate fi utilizat ca un posibil agent terapeutic în bolile neurodegenerative precum AD [143 ] .
O doză mică de tranchilizant diazepam a scăzut degradarea neuronilor, observată în progresul bolii Alzheimer. S-a demonstrat că diazepamul crește învățarea spațială și reținerea memoriei și concentrațiile de proteine normalizate asociate cu descompunerea acetilcolinei și sinteza GABA la șobolani induși cu boala Alzheimer [ 144 ].
Schetinger și colab. au studiat efectul noilor 1,5 benzodiazepine asupra activităților acetilcolinesterazei (AChE) și ATPDază (apirazei) în cortexul cerebral al șobolanilor adulți. S-a raportat că 2-triclormetil-4-fenil-3H-1,5-benzodiazepină și 2-triclormetil-4(p-metil-fenil)-3H-1,5-benzodiazepină au inhibat semnificativ activitatea acetilcolinesterazei în intervalul 0,18– 0,35 mM. De asemenea, la concentrații cuprinse între 0,063 și 0,25 mM, 1,5 benzodiazepinele au inhibat hidroliza ATP și ADP de către sinaptozomii din cortexul cerebral [ 145 ].
Se știe că daunele oxidative și inflamația au loc în afecțiunile neurodegenerative corelate cu vârsta, cum ar fi AD, în timp ce curcumina posedă efecte antioxidante și antiinflamatorii care asigură o conservare semnificativă față de agenții neurotoxici și genotoxici. Pe baza acestui fapt, Frautschy et al. au folosit șobolani Sprague-Dawley în vârstă de 22 de luni pentru a compara capacitatea ibuprofenului și curcuminei de a proteja împotriva deteriorării proteinei β -amiloid (Ap). Doar curcumina dietetică (2000 ppm) a presă daune oxidative și scăderea sinaptofizinei. Microglioza din straturile corticale a fost redusă atât de ibuprofen, cât și de curcumină. În celălalt grup al studiului, șobolani femele de vârstă mijlocie au fost tratați cu 500 ppm curcumină. Rezultatele au arătat că curcumina a demonstrat capacitatea de a preveni A β-deficiențe de memorie spațială inițiate de perfuzie și pierderea densității postsinaptice și scăderea depozitelor de A β [ 146 ].
Deoarece metalele induc agregarea amiloid-beta și toxicitatea, chelatorii, desferioxamina și cliochinolul au efecte semnificative împotriva AD. Baum și Ng au evaluat afinitatea curcuminei pentru ionii de cupru, zinc și fier. S-a stabilit că curcumina leagă mai ușor metalele redox-active de fier și cuprul decât și zincul redox-inactiv. Deoarece curcumina inhibă chelarea metalelor, studiul a clarificat că poate fi protectoare împotriva toxicității Abeta și, în același timp, poate deprima leziunile inflamatorii prin păstrarea inducției metalice a NF-kappaB [147 ] .
În condiții de agregare in vitro, curcumina a fost evidențiată la o agregare limitată la o concentrație de IC 50 : 0,8 μ M și Abeta40 IC 50 fibrilar dezagregat : 1 μ M. De asemenea, în același studiu, curcumina a fost raportată a fi un inhibitor preferabil de agregare Abeta40. decât ibuprofenul și naproxenul și previne apariția oligomerului Abeta42 la o concentrație de 0,1 până la 1,0 μ M [ 148 ].
Deoarece AD este caracterizată de acumularea de beta amiloid în celulele creierului împreună cu stresul oxidativ și inflamația, Balasubramanian a investigat eficacitatea curcuminei asupra plăcilor de beta amiloid în studiul său. Balasubramanian a raportat că curcumina se leagă mai ușor de beta-amiloid, deoarece are proprietăți adecvate de încărcare și legare, în timp ce puterea antiinflamatoare și antioxidantă depinde și de caracteristicile electronice și structurale. Dintre formele ceto și enol, s-a dovedit că forma enol are capacitatea de a trece bariera hemato-encefalică și de a se lega de beta-amiloid și are toate caracteristicile pentru un antioxidant optim [149 ] .
Utilizând microscopia fotonică multiplă (MPM), Garcia-Alloza și colab. au reprezentat că curcumina trece bariera hemato-encefalică și etichetează plăcile senile și angiopatia amiloidă cerebrovasculară (CAA) la șoarecii APPswe/PS1dE9. Curcumina a fost administrată iv din vena coadă 7,5 mg/kg/zi timp de 7 zile. Curcumina a îndepărtat și a scăzut plăcile substanțiale care au fost monitorizate și, de asemenea, a provocat o inversare restrânsă, dar semnificativă, a modificărilor constituționale ale dendritelor distrofice. Rezultatele studiului au arătat că curcumina poate fi un agent preventiv eficient pentru stresul oxidativ, inflamația și neurotoxicitatea asociate cu AD [ 150 ].
Park și colab. a indus celulele PC 12 cu A β (25-35) pentru a evalua efectele curcuminei asupra neuroprotecției. Înainte de inducerea A β (25–35), curcumina a fost tratată la o concentrație de 10 μ g/mL timp de 1 oră și a inversat semnificativ activitatea A β (25–35) prin reducerea stresului oxidativ și a deteriorării ADN-ului, pe lângă cantitățile de calciu intracelular și hiperfosforilarea tau [ 151 ].
Șobolanii induși de streptozotocină intracerebroventriculară (ICV-STZ) au fost utilizați pentru a evalua activitatea curcuminei împotriva deficienței cognitive și a deteriorării oxidative, exploatând evitarea pasivă și sarcinile labirintului de apă. Pe parcursul celor trei săptămâni s-a aplicat 80 mg/kg curcumină. Deficiențele cognitive cauzate de ICV-STZ în grupul cu curcumină s-au îmbunătățit semnificativ. Tratamentul cu curcumină a crescut în mod clar nivelurile scăzute de glutation (GSH), glutation peroxidază (GPx) și glutation reductază (GR) în hipocamp și cortexul cerebral. Ishrat și colab. a raportat că curcumina este eficientă în păstrarea deficienței cognitive și poate fi utilă în tratamentul demenței sporadice de tip Alzheimer [ 152 ].
Într-un alt studiu, influența asupra fluxului sanguin cerebral (CBF), perturbarea memoriei, stresul oxidativ și disfuncția colinergică a fost investigată la șoareci cu tulburări de memorie induse de IC-STZ cu curcumină. Șoarecilor li s-au administrat 10, 20 și 50 mg/kg, (po) curcumină, timp de 21 de zile de la prima aplicare a STZ. La șoarecii induși de STZ, curcumina a crescut fluxul sanguin cerebral într-o manieră dependentă de doză, îmbunătățind astfel deteriorarea memoriei atât în mod conservator, cât și în mod terapeutic [ 153 ].
Beta-amiloid (A β 1-40) a fost folosit pentru a induce modelul Alzheimer la șobolani pentru a evalua efectul curcuminei asupra modificărilor exprimării genelor legate de apoptoză Bax și Bcl-2 în hipocamp, pe lângă modificările memoriei spațiale. și funcția cognitivă. Șobolanii au primit 300 mg/kg/zi curcumină ip timp de șapte zile. În grupul tratat cu curcumină, expresia Bax a fost atenuată semnificativ, în timp ce expresia Bcl-2 a fost crescută. Rezultatele labirintului de apă Morris au demonstrat că curcumina a avansat în mod clar învățarea spațială și funcțiile de memorie la șobolan prin scurtarea latenței medii de evadare prin modularea Bax și Bcl-2 [ 154 ].
Influența dietei cu curcumină pe termen scurt și lung (6 săptămâni, 12 săptămâni) asupra proliferării celulare hipocampice și a funcției cognitive a fost investigată la șobolani în vârstă hrăniți cu un amestec standard de hrană de 480 mg/kg concentrație de curcumină. Cercetătorii au explorat, de asemenea, modul de acțiune de bază care ar putea conecta tratamentul cu curcumină la îmbunătățirea cogniției și a neurogenezei prin analiza secvenței exonilor a transcripției ARNm corticale și hipocampale. Acest studiu a fost clarificat cu o interferență a rețelei transcripționale a genelor implicate în neurotransmisie, dezvoltarea neuronală, transducția semnalului și metabolismul după tratamentul cu curcumină. Atât tratamentele cu curcumină, cât și pe termen lung, au îmbunătățit memoria de recunoaștere socială a șobolanilor în vârstă într-un mod semnificativ. O îmbunătățire mai puternică a memoriei spațiale a fost observată după o perioadă lungă de tratament, recomandând ca aproximativ 12 mg de curcumină pe termen lung consumului pe zi poate păstra sau ușura funcția cognitivă odată cu îmbătrânirea. Mecanismul care stă la baza efectului curcuminei asupra învățării și dezvoltării memoriei în tratamentul pe termen lung constă în promovarea neurogenezei în girusul dentat, în special la șobolanii în vârstă.155 ].
Șoarece predispus la senescență 8 (șoareci SAMP8) a fost utilizat de Sun și colab. pentru a examina dacă curcumina poate vindeca defectele cognitive. Curcumina a fost administrată intragastric șoarecilor la o doză zilnică de 20 și 50 mg/kg timp de 25 de zile. În acest studiu, a fost folosit testul labirintului cu apă Morris. Învățarea și memoria șoarecilor au fost interpretate prin examinarea memoriei spațiale, a activității superoxid dismutază (SOD), a cantității de malondialdehidă (MDA), a expresiei kinazei II dependente de p-calciu/calmodulină (p-CaMKII) și a pN-metil-d. -subunitatea 1 a receptorului de aspartat (p-NMDAR1) din hipocamp. În ambele doze, curcumina a redus semnificativ întârzierile de evacuare și a redus cantitatea de MDA și, într-o manieră dependentă de doză, a îmbunătățit expresia p-CaMKII și p-NMDAR1 [156 ] .
Banji și colab. a folosit D-galactoza pentru a diferenția dinamica mitocondrială la șobolani și pentru a genera stres oxidativ în apoptoza neuronilor. Șobolanii au primit curcumină singură (50 mg/kg, po), hesperidină singură (10 mg/kg, po) și o combinație de hesperidină și curcumină (10 mg/kg, 50 mg/kg, po) cu o săptămână înainte de D- tratament cu galactoză. În plus față de studiile comportamentale, au fost evaluate enzimele ciclului acidului tricarboxilic, complexele mitocondriale, oxidarea proteinelor și lipidelor și cantitățile de glutation în fracțiunea mitocondrială a creierului. În același timp, caspaza-3 și caspaza-3 scindată au fost studiate cu analiză Western blot. Rezultatele cercetării au arătat că curcumina s-a dovedit a fi mai puternică decât hesperidina în scăderea oxidării lipidelor, a proteinelor, a exprimării caspazei-3 scindate și a cantităților de enzime mitocondriale.157 ].
Cu studiile menționate mai sus, potența curcuminei împotriva AD a fost dovedită cu diferite mecanisme in vitro și in vivo. Eficacitatea curcuminei asupra AD este încă în curs de investigare, iar cercetările recente au fost rezumate în Tabelul 1 .
tabelul 1Rezumatul studiilor in vivo și in vitro din ultimii ani.
10.1. Studii clinice asupra curcuminei
Un studiu clinic de 6 luni a fost efectuat pentru a examina efectul curcuminei asupra măsurilor biochimice și cognitive și a siguranței la pacienții cu AD din China. Curcumina a fost aplicată în capsule sau pulbere la 1 sau 4 g. Nu a fost detectată nicio diferență în examinarea stării minime. De asemenea, rezultatele indică faptul că nu a existat nicio diferență în nivelurile serice de A β 1-40 între tratamente și în grupul de tratament cu curcumină. S-a observat că agregarea AP în creier scade. În comparație cu biodisponibilitatea, s-a raportat că capsulele sunt mai bune decât pulberea și nu există nicio diferență între 1 și 4 g de curcumină în ceea ce privește metaboliții [ 168 ] (Tabelele 2 și 3 ).
tabel 2Studii clinice finalizate cu curcumină.
Tabelul 3Studii clinice nepublicate cu curcumină.
Curcumină, bisdemetoxicurcumină și amestec demetoxicurcumină numit Curcumin 3 Complex® a fost aplicată, 4 g/zi, timp de 24 de săptămâni, participanților. Nu au fost găsite date privind eficacitatea formulării în cogniție și AP, iar în LCR, nivelurile de tau și biodisponibilitatea în plasmă au fost scăzute [ 169 ].
Cox şi colab. a efectuat un studiu între adulți cu vârste cuprinse între 65 și 80 de ani. Tratamentul unei doze de 400 mg de Longvida® care conține 80 mg curcumină a fost aplicat ca acut cronic și acut pe cronic. La o oră după administrare, aplicarea curcuminei a avut un efect pozitiv asupra memoriei de lucru și asupra măsurătorilor continue a atenției. Cercetătorii au descoperit că au observat un efect similar după administrarea cronică și, de asemenea, au afirmat că nu au existat rezultate similare la 3 ore după administrarea acută. În plus, în grupul de curcumină cronică s-a dezvoltat starea de spirit; s-a observat creșterea calității și scăderea prostrației [ 170 ].
Formularea de curcumină BiocurcumaxTM a fost studiată pentru efectul său de prevenire asupra declinului cognitiv la adulții în vârstă într-o cercetare dublu orb, randomizat. 1500 mg/zi BiocurcumaxTM au fost administrate participanților timp de 12 luni. În studiu, nu a fost observată nicio diferență între grupuri pentru măsurători clinice și cognitive [ 171 ].
Theracurmin®, care conține 90 mg curcumină, a fost examinat pe 40 de participanți nedemenți și în vârstă de 51-84 de ani. Formularea și placebo au fost aplicate de două ori pe zi timp de 18 luni. Descoperirile studiului au inclus o scădere a formării de amiloid și a acumularii tau, precum și o mai bună atenție și memorie. [ 172 ].
Deși curcumina este eficientă împotriva unui număr mare de boli, s-a demonstrat că limitează eficacitatea terapeutică a biodisponibilității slabe din cauza absorbției slabe, metabolismului rapid și eliminării sistemice rapide. Ca urmare, s-au făcut numeroase eforturi pentru a îmbunătăți biodisponibilitatea curcuminei prin modificarea acestor proprietăți. Utilizarea adjuvanților pentru a restrânge calea metabolică a curcuminei este cea mai comună strategie de îmbunătățire a biodisponibilității curcuminei. De asemenea, alte abordări încurajatoare pentru a crește biodisponibilitatea curcuminei la oameni includ nanoparticule, lipozomi, complexe fosfolipide și modificări structurale [ 173 ]. Tabelul 2 enumeră studiile clinice finalizate cu curcumină, în timp ce tabelul 3 prezintă studiile clinice nepublicate cu curcumină.
10.2. Efecte secundare/Toxicitatea curcuminei
În general, studiile preclinice și clinice au raportat că curcumina nu are efecte secundare semnificative; un număr limitat de studii au demonstrat efecte adverse minore ale curcuminei la doze mari. Curcumina este considerată sigură de FDA. Datorită efectului curcuminei asupra citocromului P450, glutation-S-transferazei și UDP-glucuronoziltransferazei, s-a demonstrat că afectează metabolismul diferitelor medicamente și s-a raportat că provoacă o creștere a concentrației plasmatice [174 ] . De asemenea, în unele studii, s-a clarificat că, într-o manieră dependentă de doză, curcumina induce modificări cromozomiale și provoacă modificări ale ADN-ului în diferite linii celulare de mamifere [175, 176 ] .]. Într-un studiu clinic al lui Sharma și colab., s-au observat greață și diaree și niveluri crescute de fosfatază alcalină serică și lactat dehidrogenază pe o perioadă de 1-4 luni, cu un interval de doze de 0,45-3,6 g/zi [177 ] . Hanai și colab. a raportat, de asemenea, efecte secundare gastrointestinale minore la pacienții cu colită ulceroasă [ 178 ]. Pe de altă parte, multe studii clinice cu intervale de timp diferite și doze diferite nu au raportat efecte secundare și au evaluat curcumina ca fiind sigură [ 179 – 183 ].
11. Concluzie și perspective de viitor
Deși studiile in vitro și in vivo au arătat rezultate promițătoare, studiile clinice au demonstrat biodisponibilitatea scăzută a curcuminei; prin urmare, în doze mici, nu a fost observată nicio activitate la pacienți. Pentru a depăși această problemă, ar trebui dezvoltate noi formulări cu diferite căi de administrare. În plus, dificultățile de diagnostic și chiar diagnosticarea greșită a bolilor neurologice, în principal AD, sunt încă una dintre problemele majore, în ciuda îmbunătățirilor tehnologice. Pentru cel mai adecvat regim de tratament și o eficiență sporită a tratamentului selectat, diagnosticul și mecanismul bolii trebuie mai întâi dezvăluite corect.
Disponibilitatea datelor
Datele folosite pentru a susține concluziile acestui studiu sunt toate incluse și disponibile în articol.
Conflicte de interes
Autorii declară că nu au interese financiare concurente cunoscute sau relații personale care ar fi putut părea să influențeze munca raportată în această lucrare.
Contribuțiile autorilor
Conceptualizarea a fost realizată de EKA; redactarea manuscrisului a fost realizată de EKA, HB, Ç.Y., G.Ş.K., şi BK; revizuirea și editarea lucrării au fost efectuate de EKA și HK; revizuirea a fost făcută EKA, HB, Ç.Y., şi G.Ş.K. Toți autorii au citit și au fost de acord cu versiunea publicată a manuscrisului.
Referințe
A. Bhat, AM Mahalakshmi, B. Ray și colab., „Beneficiile curcuminei în tulburările creierului”, BioFactors , voi. 45, nr. 5, p. 666–689, 2019.Vizualizare la: Site-ul editorului | Google Academic
M. Tang și C. Taghibiglou, „Mecanismele de acțiune ale curcuminei în boala Alzheimer”, Journal of Alzheimer’s Disease , voi. 58, nr. 4, p. 1003–1016, 2017.Vizualizare la: Site-ul editorului | Google Academic
S. Jagtap, K. Meganathan, V. Wagh, J. Winkler, J. Hescheler și A. Sachinidis, „Mecanismul chimioprotector al compușilor naturali, epigallocatechin-3-O-galat, quercetină și curcumină împotriva cancerului și a bolilor cardiovasculare, ” Current Medicinal Chemistry , voi. 16, nr. 12, p. 1451–1462, 2009.Vizualizare la: Site-ul editorului | Google Academic
RA Sharma, CR Ireson, RD Verschoyle și colab., „Efectele curcuminei dietetice asupra glutation S-transferazei și aductilor malondialdehide-ADN în mucoasa ficatului și colonului de șobolan: relația cu nivelurile de medicamente”, Clinical Cancer Research, voi . 7, nr. 5, p. 1452–1458, 2001.Vizualizați la: Google Scholar
DS El-Agamy, „Efectele comparative ale curcuminei și resveratrolului asupra leziunilor hepatice induse de aflatoxină B(1) la șobolani”, Arhivele de toxicologie , voi. 84, nr. 5, p. 389–396, 2010.Vizualizare la: Site-ul editorului | Google Academic
BB Aggarwal, „Întrețirea obezității induse de inflamație și a bolilor metabolice de către curcumină și alte nutraceutice”, Annual Review of Nutrition , voi. 30, nr. 1, p. 173–199, 2010.Vizualizare la: Site-ul editorului | Google Academic
L. Alappat și A. Awad, „Curcumină și obezitate: dovezi și mecanisme”, Curcumină și obezitate: dovezi și mecanisme , voi. 68, p. 729–738, 2010.Vizualizați la: Google Scholar
A. Darvesh, B. Aggarwal și A. Bishayee, „Curcumină și cancer de ficat: o revizuire”, Current Pharmaceutical Biotechnology , voi. 13, nr. 1, p. 218–228, 2012.Vizualizare la: Site-ul editorului | Google Academic
JL Funk, JN Oyarzo, JB Frye și colab., „Extractele de turmeric care conțin curcuminoizi previn artrita reumatoidă experimentală”, Journal of Natural Products , voi. 69, nr. 3, p. 351–355, 2006.Vizualizare la: Site-ul editorului | Google Academic
B. Chandran și A. Goel, „Un studiu pilot randomizat pentru a evalua eficacitatea și siguranța curcuminei la pacienții cu poliartrită reumatoidă activă”, Phytotherapy Research , voi. 26, nr. 11, p. 1719–1725, 2012.Vizualizare la: Site-ul editorului | Google Academic
KS Lee, BS Lee, S. Semnani et al., „Curcuminul prelungește durata de viață, îmbunătățește durata sănătății și modulează expresia genelor de îmbătrânire asociate cu vârsta la Drosophila melanogaster ”, Rejuvenation Research , voi. 13, nr. 5, p. 561–570, 2010.Vizualizare la: Site-ul editorului | Google Academic
VH Liao, CW Yu, YJ Chu, WH Li, YC Hsieh și TT Wang, „Extinderea duratei de viață mediată de curcumin în Caenorhabditis elegans ”, Mecanisme de îmbătrânire și dezvoltare , voi. 132, nr. 10, p. 480–487, 2011.Vizualizare la: Site-ul editorului | Google Academic
B. Salehi, D. Calina, AO Docea et al., „Curcumin’s nanomedicine formulations for therapeutic application in neurologic diseases”, Journal of Clinical Medicine , voi. 9, nr. 2, p. 430, 2020.Vizualizare la: Site-ul editorului | Google Academic
A. Noorafshan, F. Karimi, AM Kamali, S. Karbalay-Doust și M. Nami, „Ar putea curcumina să protejeze arborii dendritici ai neuronilor CA1 de scurtarea și scurgerea induse de restricția cronică a somnului la șobolani?” Științe ale vieții , voi. 198, p. 65–70, 2018.Vizualizare la: Site-ul editorului | Google Academic
A. Kumar și A. Singh, „Posibilă modulare a oxidului nitric în efectul protector al ( Curcuma longa , Zingiberaceae) împotriva modificărilor comportamentale induse de privarea de somn și daune oxidative la șoareci”, Phytomedicine , voi. 15, nr. 8, p. 577–586, 2008.Vizualizare la: Site-ul editorului | Google Academic
A. Noorafshan, F. Karimi, S. Karbalay-Doust și AM Kamali, „Utilizarea curcuminei pentru a preveni modificările structurale și comportamentale ale cortexului prefrontal medial induse de privarea de somn la șobolani”, EXCLI Journal, voi . 16, p. 510–520, 2017.Vizualizare la: Site-ul editorului | Google Academic
A. Noorafshan, F. Karimi, AM Kamali, S. Karbalay-Doust și M. Nami, „Restaurative effects of curcumin on sleep-induced memory impairments and structurel changes of the hipocampus in a rat model,” Life Sciences, vol . . 189, p. 63–70, 2017.Vizualizare la: Site-ul editorului | Google Academic
E. Hita-Yañez, M. Atienza și JL Cantero, „Marerii de somn polisomnografici și subiectivi ai deficienței cognitive ușoare”, Sleep (Basel) , voi. 36, nr. 9, p. 1327–1334, 2013.Vizualizare la: Site-ul editorului | Google Academic
M. Hu, P. Zhang, C. Li și colab., „Turburări de somn în tulburări cognitive ușoare: o revizuire sistematică a măsurilor obiective”, Științe Neurologice , voi. 38, nr. 8, p. 1363–1371, 2017.Vizualizare la: Site-ul editorului | Google Academic
RC Petersen și JC Morris, „Deficiența cognitivă ușoară ca entitate clinică și țintă de tratament”, Archives of Neurology , voi. 62, nr. 7, p. 1160–3; discuția 1167, 2005.Vizualizare la: Site-ul editorului | Google Academic
S. Beaulieu-Bonneau și C. Hudon, „Turburări de somn la adulții în vârstă cu deficiențe cognitive ușoare”, International Psychogeriatrics , voi. 21, nr. 4, p. 654–666, 2009.Vizualizare la: Site-ul editorului | Google Academic
S. Bombois, P. Derambure, F. Pasquier și C. Monaca, „Tulburări de somn în îmbătrânire și demență”, The Journal of Nutrition, Health & Aging , voi. 14, nr. 3, p. 212–217, 2010.Vizualizare la: Site-ul editorului | Google Academic
M. Bidzan, L. Bidzan și I. Bidzan-Bluma, „Simptome neuropsihiatrice și progresie mai rapidă a deficiențelor cognitive ca predictori ai riscului de conversie a deficienței cognitive ușoare la demență”, Archives of Medical Science , voi. 13, nr. 5, p. 1168–1177, 2017.Vizualizare la: Site-ul editorului | Google Academic
W. Muangpaisan, S. Intalapaporn și P. Assantachai, „Simptome neuropsihiatrice la pacienții comunitari cu deficiențe cognitive ușoare și influența factorilor demografici”, Jurnalul Internațional de Psihiatrie Geriatrică , voi. 23, nr. 7, p. 699–703, 2008.Vizualizare la: Site-ul editorului | Google Academic
SL Naismith, NL Rogers, IB Hickie, J. Mackenzie, LM Norrie și SJG Lewis, „Sleep well, think well: sleep-wake perturbation in mild cognitive impairment”, Journal of Geriatric Psychiatry and Neurology, voi . 23, nr. 2, p. 123–130, 2010.Vizualizare la: Site-ul editorului | Google Academic
DW Kang, CU Lee și HK Lim, „Rolul tulburărilor de somn în traiectoria bolii Alzheimer”, Clinical Psychopharmacology and Neuroscience , voi. 15, nr. 2, p. 89–99, 2017.Vizualizare la: Site-ul editorului | Google Academic
CE Westerberg, EM Lundgren, SM Florczak și colab., „Somnul influențează severitatea tulburărilor de memorie în deficiența cognitivă ușoară amnestică: rezultate din autoevaluarea somnului și monitorizarea continuă a activității”, Alzheimer Disease and Associated Disorders, voi . 24, nr. 4, p. 325–333, 2010.Vizualizare la: Site-ul editorului | Google Academic
E. Hita-Yanez, M. Atienza, EL Gil-Neciga și J. Cantero, „Modele de somn perturbate la vârstnici cu deficiențe cognitive ușoare: rolul declinului memoriei și al genotipului ApoE ε 4”, Current Alzheimer Research , voi . 9, nr. 3, p. 290–297, 2012.Vizualizare la: Site-ul editorului | Google Academic
L. Peter-Derex, P. Yammine, H. Bastuji și B. Croisile, „Somnul și boala Alzheimer”, Sleep Medicine Reviews , voi. 19, p. 29–38, 2015.Vizualizare la: Site-ul editorului | Google Academic
MV Vitiello, PN Prinz, DE Williams, MS Frommlet și RK Ries, „Tulburări de somn la pacienții cu boala Alzheimer în stadiu uşoară”, Journal of Gerontology , voi. 45, nr. 4, p. M131–M138, 1990.Vizualizare la: Site-ul editorului | Google Academic
D. Petit, JF Gagnon, ML Fantini, L. Ferini-Strambi și J. Montplaisir, „Somn și EEG cantitativ în tulburările neurodegenerative”, Journal of Psychosomatic Research , voi. 56, nr. 5, p. 487–496, 2004.Vizualizare la: Site-ul editorului | Google Academic
DL Bliwise, „Somn în îmbătrânire normală și demență”, Sleep , voi. 16, nr. 1, p. 40–81, 1993.Vizualizare la: Site-ul editorului | Google Academic
DL Bliwise, „Tulburările de somn în boala Alzheimer și alte demențe”, Clinical Cornerstone , voi. 6, nr. 1, p. S16–S28, 2004.Vizualizare la: Site-ul editorului | Google Academic
CM Walsh, T. Blackwell, GJ Tranah et al., „Ritmurile de activitate circadiană mai slabe sunt asociate cu o funcție executivă mai slabă la femeile în vârstă”, Sleep (Basel) , voi. 37, nr. 12, p. 2009–2016, 2014.Vizualizare la: Site-ul editorului | Google Academic
D. Foley, A. Monjan, K. Masaki și colab., „Somnolența în timpul zilei este asociată cu demența incidentă de 3 ani și declinul cognitiv la bărbații americani-japonezi mai în vârstă”, Journal of the American Geriatrics Society, voi . 49, nr. 12, p. 1628–1632, 2001.Vizualizare la: Site-ul editorului | Google Academic
PC Elwood, AJ Bayer, M. Fish, J. Pickering, C. Mitchell și JEJ Gallacher, „Tulburările de somn și somnolența diurnă prezic demența vasculară”, Journal of Epidemiology and Community Health , voi. 65, nr. 9, p. 820–824, 2011.Vizualizare la: Site-ul editorului | Google Academic
JJ Virta, K. Heikkilä, M. Perola și colab., „Caracteristicile somnului la mijlocul vârstei asociate cu funcția cognitivă târzie a vieții”, Sleep (Basel) , voi. 36, nr. 10, p. 1533–1541, 2013.Vizualizare la: Site-ul editorului | Google Academic
JC Chen, MA Espeland, RL Brunner și colab., „Durata somnului, declinul cognitiv și riscul de demență la femeile în vârstă”, Alzheimer & Dementia , voi. 12, nr. 1, p. 21–33, 2016.Vizualizare la: Site-ul editorului | Google Academic
O. Potvin, D. Lorrain, H. Forget și colab., „Calitatea somnului și deteriorarea cognitivă incidentă de 1 an la adulții în vârstă care locuiesc în comunitate”, Sleep (Basel) , voi. 35, nr. 4, p. 491–499, 2012.Vizualizare la: Site-ul editorului | Google Academic
L. Shi, SJ Chen, MY Ma și colab., „Tulburările de somn cresc riscul de demență: o revizuire sistematică și meta-analiză”, Sleep Medicine Reviews , voi. 40, p. 4–16, 2018.Vizualizare la: Site-ul editorului | Google Academic
M. Cohen-Zion, C. Stepnowsky, T. Marler, T. Shochat, DF Kripke și S. Ancoli-Israel, „Schimbări ale funcției cognitive asociate cu tulburările de respirație în somn la persoanele în vârstă”, Journal of the American Geriatrics Society , vol. 49, nr. 12, p. 1622–1627, 2001.Vizualizare la: Site-ul editorului | Google Academic
J. Benito-León, F. Bermejo-Pareja, S. Vega și ED Louis, „Durata totală a somnului zilnic și riscul de demență: un studiu prospectiv bazat pe populație”, Jurnalul European de Neurologie, voi . 16, nr. 9, p. 990–997, 2009.Vizualizare la: Site-ul editorului | Google Academic
A. Loerbroks, D. Debling, M. Amelang și T. Stürmer, „Durata somnului nocturn și afectarea cognitivă într-un studiu bazat pe populație asupra adulților în vârstă”, Jurnalul Internațional de Psihiatrie Geriatrică , voi. 25, nr. 1, p. 100–109, 2010.Vizualizare la: Site-ul editorului | Google Academic
GJ Tranah, T. Blackwell, KL Stone și colab., „Circadian activity rhythms and risk of incident dementa and mild cognitive impairment la femeile în vârstă”, Annals of Neurology , voi. 70, nr. 5, p. 722–732, 2011.Vizualizare la: Site-ul editorului | Google Academic
pentru Grupul de studiu privind fracturile osteoporotice la bărbați (MrOS), T. Blackwell, K. Yaffe și colab., „Asociații de calitate a somnului măsurată în mod obiectiv și subiectiv cu declin cognitiv ulterior la bărbații în vârstă care locuiesc în comunitate: studiul despre somn MrOS,” Sleep (Basel) , voi. 37, nr. 4, p. 655–663, 2014.Vizualizare la: Site-ul editorului | Google Academic
K. Yaffe, AM Laffan, SL Harrison și colab., „Turburări de respirație în somn, hipoxie și risc de afectare cognitivă ușoară și demență la femeile în vârstă”, Jurnalul Asociației Medicale Americane , voi. 306, nr. 6, p. 613–619, 2011.Vizualizare la: Site-ul editorului | Google Academic
MP Pase, JJ Himali, NA Grima et al., „Arhitectura somnului și riscul de demență incidentă în comunitate”, Neurologie , voi. 89, nr. 12, p. 1244–1250, 2017.Vizualizare la: Site-ul editorului | Google Academic
KM de Almondes, MV Costa, LF Malloy-Diniz și BS Diniz, „Insomnie și risc de demență la adulții în vârstă: revizuire sistematică și meta-analiza”, Journal of Psychiatric Research , voi. 77, p. 109–115, 2016.Vizualizare la: Site-ul editorului | Google Academic
OM Bubu, M. Brannick, J. Mortimer și colab., „Somnul, tulburările cognitive și boala Alzheimer: o revizuire sistematică și meta-analiză”, Sleep (Basel) , voi. 40, nr. 1, 2017.Vizualizare la: Site-ul editorului | Google Academic
Jack CR Jr, MS Albert, DS Knopman et al., „Introducere la recomandările de la Institutul Național pentru Îmbătrânire-Grupurile de lucru Asociația Alzheimer privind ghidurile de diagnostic pentru boala Alzheimer”, Alzheimer și demență: jurnalul Asociației Alzheimer, voi . 7, nr. 3, p. 257–262, 2011.Vizualizare la: Site-ul editorului | Google Academic
RS Osorio, I. Ayappa, J. Mantua și colab., „Interacțiunea dintre respirația tulburată de somn și genotipul apolipoproteinei E pe biomarkerii lichidului cefalorahidian pentru boala Alzheimer la persoanele în vârstă din punct de vedere cognitiv normal”, Neurobiology of Aging, voi . 35, nr. 6, p. 1318–1324, 2014.Vizualizare la: Site-ul editorului | Google Academic
YE Ju, BP Lucey și DM Holtzman, „Somnul și patologia bolii Alzheimer – o relație bidirecțională”, Nature Reviews Neurology , voi. 10, nr. 2, p. 115–119, 2014.Vizualizare la: Site-ul editorului | Google Academic
TT Dang-Vu, M. Schabus, M. Desseilles, V. Sterpenich, M. Bonjean și P. Maquet, „Insights functional neuroimaging in the physiology of human sleep”, Sleep , voi. 33, nr. 12, p. 1589–1603, 2010.Vizualizare la: Site-ul editorului | Google Academic
JE Kang, MM Lim, RJ Bateman și colab., „Dinamica beta-amiloid este reglată de orexină și de ciclul somn-veghe”, Science , voi. 326, nr. 5955, p. 1005–1007, 2009.Vizualizare la: Site-ul editorului | Google Academic
Y. Huang, „Efectele vârstei și depunerea de amiloid asupra dinamicii β în sistemul nervos central uman”, Arhivele Neurologiei , voi. 69, nr. 1, p. 51–58, 2012.Vizualizare la: Site-ul editorului | Google Academic
MK Rasmussen, H. Mestre și M. Nedergaard, „Calea glimfatică în tulburările neurologice”, Lancet Neurology , voi. 17, nr. 11, p. 1016–1024, 2018.Vizualizare la: Site-ul editorului | Google Academic
NA Jessen, AS Munk, I. Lundgaard și M. Nedergaard, „Sistemul glimfatic: un ghid pentru începători”, Cercetare neurochimică , voi. 40, nr. 12, p. 2583–2599, 2015.Vizualizare la: Site-ul editorului | Google Academic
L. Xie, H. Kang, Q. Xu și colab., „Somnul conduce la eliminarea metaboliților din creierul adult”, Science , voi. 342, nr. 6156, p. 373–377, 2013.Vizualizare la: Site-ul editorului | Google Academic
AW Bero, P. Yan, JH Roh și colab., „Activitatea neuronală reglează vulnerabilitatea regională la depunerea de amiloid- β ”, Nature Neuroscience , voi. 14, nr. 6, p. 750–756, 2011.Vizualizare la: Site-ul editorului | Google Academic
YE Ju, JS McLeland, CD Toedebusch și colab., „Calitatea somnului și boala Alzheimer preclinic”, JAMA Neurology , voi. 70, nr. 5, p. 587–593, 2013.Vizualizare la: Site-ul editorului | Google Academic
ES Musiek și DM Holtzman, „Mecanisme care leagă ceasurile circadiene, somnul și neurodegenerarea”, Science , voi. 354, nr. 6315, p. 1004–1008, 2016.Vizualizare la: Site-ul editorului | Google Academic
L. Zhou, Q. Gao, M. Nie și colab., „Degenerarea și deficitul de energie în nucleul suprachiasmatic stau la baza perturbării ritmului circadian la șoareci ApoE -/- : implicații pentru boala Alzheimer,” Rapoarte științifice , voi. 6, nr. 1, p. 36335, 2016.Vizualizare la: Site-ul editorului | Google Academic
BA Kent și RE Mistlberger, „Somnul și neurogeneza hipocampului: Implicații pentru boala Alzheimer”, Frontiers in Neuroendocrinology , voi. 45, p. 35–52, 2017.Vizualizare la: Site-ul editorului | Google Academic
SK Inouye, ST Bogardus Jr., PA Charpentier și colab., „O intervenție multicomponentă pentru prevenirea delirului la pacienții în vârstă spitalizați”, The New England Journal of Medicine , voi. 340, nr. 9, p. 669–676, 1999.Vizualizare la: Site-ul editorului | Google Academic
K. Meguro, S. Yamaguchi, M. Shimada, M. Itoh și A. Yamadori, „Transmiterea dopaminergică striată și utilizarea neocorticală a glucozei în boala Alzheimer: un studiu de tomografie cu emisie de pozitroni cu trei trasare”, Arhivele de Gerontologie și Geriatrie, vol . . 31, nr. 2, p. 147–158, 2000.Vizualizare la: Site-ul editorului | Google Academic
ST O’Keeffe, K. Gavin și JN Lavan, „Statutul fierului și sindromul picioarelor neliniștite la vârstnici”, Age and Aging , voi. 23, nr. 3, p. 200–203, 1994.Vizualizare la: Site-ul editorului | Google Academic
JR McCarten, C. Kovera, MK Maddox și JP Cleary, „Triazolam în boala Alzheimer: studiu pilot asupra efectelor somnului și memoriei”, Farmacologie, biochimie și comportament , voi. 52, nr. 2, p. 447–452, 1995.Vizualizare la: Site-ul editorului | Google Academic
GK Lloyd, A. Lowenthal, F. Javoy-Agid și J. Constantidinis, „GAEA A receptor complex function in frontal cortex membranes from control and neuroological patients”, European Journal of Pharmacology , voi. 197, nr. 1-2, p. 33–39, 1991.Vizualizare la: Site-ul editorului | Google Academic
SH Shaw, H. Curson și JE Coquelin, „Un studiu dublu-orb, comparativ al zolpidemului și placebo în tratamentul insomniei la pacienții vârstnici de psihiatrie internați”, The Journal of International Medical Research, voi . 20, nr. 2, p. 150–161, 1992.Vizualizare la: Site-ul editorului | Google Academic
PS Shelton și LB Hocking, „Zolpidem pentru insomnia legată de demență și rătăcirea nocturnă”, The Annals of Pharmacotherapy , voi. 31, nr. 3, p. 319–322, 1997.Vizualizare la: Site-ul editorului | Google Academic
A. Daniele, A. Albanese, G. Gainotti, B. Gregori și P. Bartolomeo, „Zolpidem in Parkinson’s disease”, Lancet , voi. 349, nr. 9060, p. 1222-1223, 1997.Vizualizare la: Site-ul editorului | Google Academic
M. Ansseau, W. Pitchot, M. Hansenne și AG Moreno, „Reacții psihotice la zolpidem”, Lancet , voi. 339, nr. 8796, p. 809, 1992.Vizualizare la: Site-ul editorului | Google Academic
J. Avorn, SB Soumerai, DE Everitt et al., „Un studiu randomizat al unui program de reducere a consumului de medicamente psihoactive în casele de bătrâni”, The New England Journal of Medicine , voi. 327, nr. 3, p. 168–173, 1992.Vizualizare la: Site-ul editorului | Google Academic
RI Shorr, RL Fought și WA Ray, „Schimbări în utilizarea medicamentelor antipsihotice în casele de bătrâni în timpul implementării reglementărilor OBRA-87”, Jurnalul Asociației Medicale Americane , vol. 271, nr. 5, p. 358–362, 1994.Vizualizare la: Site-ul editorului | Google Academic
ME Exum, BJ Phelps, KE Nabers și JG Osborne, „Sindromul Sundown: se reflectă în utilizarea medicamentelor PRN pentru rezidenții caselor de bătrâni?” Gerontologul , voi. 33, nr. 6, p. 756–761, 1993.Vizualizare la: Site-ul editorului | Google Academic
ME Tinetti, M. Speechley și SE Ginter, „Factori de risc pentru căderi în rândul persoanelor în vârstă care trăiesc în comunitate”, The New England Journal of Medicine , voi. 319, nr. 26, p. 1701–1707, 1988.Vizualizare la: Site-ul editorului | Google Academic
WA Ray, MR Griffin, W. Schaffner, DK Baugh și LJ Melton III, „Utilizarea de droguri psihotrope și riscul de fractură de șold”, The New England Journal of Medicine , voi. 316, nr. 7, p. 363–369, 1987.Vizualizare la: Site-ul editorului | Google Academic
PS Wang, RL Bohn, RJ Glynn, H. Mogun și J. Avorn, „Utilizarea zolpidemului și fracturile de șold la persoanele în vârstă”, Journal of the American Geriatrics Society , voi. 49, nr. 12, p. 1685–1690, 2001.Vizualizare la: Site-ul editorului | Google Academic
MB King și ME Tinetti, „Căderi ale persoanelor în vârstă care locuiesc în comunitate”, Jurnalul Societății Americane de Geriatrie , voi. 43, nr. 10, p. 1146–1154, 1995.Vizualizare la: Site-ul editorului | Google Academic
WA Ray, PB Thapa și P. Gideon, „Benzodiazepine și riscul de cădere la rezidenții caselor de bătrâni”, Jurnalul Societății Americane de Geriatrie , voi. 48, nr. 6, p. 682–685, 2000.Vizualizare la: Site-ul editorului | Google Academic
GS Brassington, AC King și DL Bliwise, „Problemele de somn ca factor de risc pentru căderi la un eșantion de adulți care locuiesc în comunitate cu vârsta cuprinsă între 64 și 99 de ani”, Jurnalul Societății Americane de Geriatrie , voi. 48, nr. 10, p. 1234–1240, 2000.Vizualizare la: Site-ul editorului | Google Academic
D. Riemann, H. Gann, H. Dressing, WE Müller și JB Aldenhoff, „Influența inhibitorului de colinesterază bromhidrat de galantamina asupra somnului normal”, Psychiatry Research , voi. 51, nr. 3, p. 253–267, 1994.Vizualizare la: Site-ul editorului | Google Academic
M. Schredl, B. Weber, ML Leins și I. Heuser, „Mărirea somnului REM indusă de donepezil îmbunătățește performanța memoriei la persoanele în vârstă, sănătoase”, Experimental Gerontology , voi. 36, nr. 2, p. 353–361, 2001.Vizualizare la: Site-ul editorului | Google Academic
E. Holsboer-Trachsler, M. Hatzinger, R. Stohler și colab., „Efectele noului inhibitor de acetilcolinesterază SDZ ENA 713 asupra somnului la om”, Neuropsychopharmacology , voi. 8, nr. 1, p. 87–92, 1993.Vizualizare la: Site-ul editorului | Google Academic
GA Yorston și R. Gray, „Halucinații hipnopompice cu donepezil”, Journal of Psychopharmacology , voi. 14, nr. 3, p. 303-304, 2000.Vizualizare la: Site-ul editorului | Google Academic
JS Ross și JR Shua-Haim, „Coșmarurile induse de Aricept în boala Alzheimer: 2 rapoarte de caz”, Journal of the American Geriatrics Society , voi. 46, nr. 1, p. 119-120, 1998.Vizualizare la: Site-ul editorului | Google Academic
SL Rogers, RS Doody, RC Mohs și LT Friedhoff, „Donepezilul îmbunătățește cogniția și funcția globală în boala Alzheimer: un studiu de 15 săptămâni, dublu-orb, controlat cu placebo. Grupul de studiu Donepezil”, Arhivele de Medicină Internă , voi. 158, nr. 9, p. 1021–1031, 1998.Vizualizare la: Site-ul editorului | Google Academic
A. Burns, M. Rossor, J. Hecker și colab., „Efectele donepezilului în boala Alzheimer – rezultate dintr-un studiu multinațional”, Dementa and Geriatric Cognitive Disorders , voi. 10, nr. 3, p. 237–244, 1999.Vizualizare la: Site-ul editorului | Google Academic
I. McKeith, T. del Ser, E. Spano et al., „Eficacitatea rivastigminei în demența cu corpi Lewy: un studiu internațional randomizat, dublu-orb, controlat cu placebo”, Lancet, voi . 356, nr. 9247, p. 2031–2036, 2000.Vizualizare la: Site-ul editorului | Google Academic
JS Markowitz, EM Gutterman, S. Lilienfeld și G. Papadopoulos, „Rezultatele legate de somn la persoanele cu boală Alzheimer uşoară până la moderată într-un studiu controlat cu placebo de galantamina”, Sleep, voi . 26, nr. 5, p. 602–606, 2003.Vizualizare la: Site-ul editorului | Google Academic
SM Stahl, JS Markowitz, EM Gutterman și G. Papadopoulos, „Utilizarea concomitentă de donepezil și hipnotice în rândul pacienților cu boala Alzheimer care trăiesc în comunitate”, Journal of Clinical Psychiatry , voi. 64, nr. 4, p. 466–472, 2003.Vizualizare la: Site-ul editorului | Google Academic
LE Maclean, CC Collins și EJ Byrne, „Dementa cu corpi Lewy tratați cu rivastigmină: efecte asupra cogniției, simptomelor neuropsihiatrice și somnului”, International Psychogeriatrics , voi. 13, nr. 3, p. 277–288, 2001.Vizualizare la: Site-ul editorului | Google Academic
PJ Reading, AK Luce și IG McKeith, „Rivastigmina în tratamentul psihozei parkinsoniene și a tulburărilor cognitive: constatări preliminare dintr-un studiu deschis”, Movement Disorders , voi. 16, nr. 6, p. 1171–1174, 2001.Vizualizare la: Site-ul editorului | Google Academic
KM Nelson, JL Dahlin, J. Bisson, J. Graham, GF Pauli și MA Walters, „Chimia medicinală esențială a curcuminei”, Journal of Medicinal Chemistry , voi. 60, nr. 5, p. 1620–1637, 2017.Vizualizare la: Site-ul editorului | Google Academic
KI Priyadarsini, „Chimia curcuminei: de la extracție la agent terapeutic”, Molecules , voi. 19, nr. 12, p. 20091–20112, 2014.Vizualizare la: Site-ul editorului | Google Academic
WH Lee, CY Loo, M. Bebawy, F. Luk, R. Mason și R. Rohanizadeh, „Curcuminul și derivații săi: aplicarea lor în neurofarmacologie și neuroștiință în secolul 21”, Current Neuropharmacology, voi . 11, nr. 4, p. 338–378, 2013.Vizualizare la: Site-ul editorului | Google Academic
H. Hatcher, R. Planalp, J. Cho, FM Torti și SV Torti, „Curcumin: from ancient medicine to current clinical trials”, Cellular and Molecular Life Sciences , voi. 65, nr. 11, p. 1631–1652, 2008.Vizualizare la: Site-ul editorului | Google Academic
Y. Katsuyama, T. Kita, N. Funa și S. Horinouchi, „Curcuminoid biosynthesis by two type III polyketide synthases in the herb Curcuma longa ”, Biological Chemistry , voi. 284, nr. 17, p. 11160–11170, 2009.Vizualizare la: Site-ul editorului | Google Academic
MM Wen, NS el-Salamouni, WM el-Refaie și colab., „Sisteme de administrare a medicamentelor bazate pe nanotehnologie pentru managementul bolii Alzheimer: provocări tehnice, industriale și clinice”, Journal of Controlled Release, voi . 245, p. 95–107, 2017.Vizualizare la: Site-ul editorului | Google Academic
TT Nguyen, TT Dung Nguyen, TK Vo și colab., „Livrarea medicamentelor pe bază de nanotehnologie pentru tulburările sistemului nervos central”, Biomedicine & Pharmacotherapy , voi. 143, p. 112117, 2021.Vizualizare la: Site-ul editorului | Google Academic
AK Singh, SN Rai, A. Maurya și colab., „Potențialul terapeutic al fitoconstituenților în managementul bolii Alzheimer”, Medicina complementară și alternativă bazată pe dovezi, , voi. 2021, articol 5578574, p. 1–19, 2021.Vizualizare la: Site-ul editorului | Google Academic
KK Cheng, CF Yeung, SW Ho, SF Chow, AHL Chow și L. Baum, „Nanoparticule de curcumină foarte stabilizate testate într-un model in vitro de barieră hematoencefalică și în șoarecii Tg2576 cu boala Alzheimer”, The AAPS Journal, voi . 15, nr. 2, p. 324–336, 2013.Vizualizare la: Site-ul editorului | Google Academic
O. Naksuriya, S. Okonogi, RM Schiffelers și WE Hennink, „Nanoformulații de curcumină: o revizuire a proprietăților farmaceutice și a studiilor preclinice și a datelor clinice legate de tratamentul cancerului”, Biomaterials , voi. 35, nr. 10, p. 3365–3383, 2014.Vizualizare la: Site-ul editorului | Google Academic
MM Yallapu, PKB Nagesh, M. Jaggi și SC Chauhan, „Aplicații terapeutice ale nanoformulărilor de curcumină”, The AAPS Journal , voi. 17, nr. 6, p. 1341–1356, 2015.Vizualizare la: Site-ul editorului | Google Academic
D. Ege, „Mecanismele de acțiune ale curcuminei în boala Alzheimer și livrarea țintită a creierului”, Materials , voi. 14, nr. 12, p. 3332, 2021.Vizualizare la: Site-ul editorului | Google Academic
U. Shabbir, M. Rubab, A. Tyagi și DH Oh, „Curcuminul și derivații săi ca agenți teranostici în boala Alzheimer: implicația nanotehnologiei”, Jurnalul Internațional de Științe Moleculare, voi . 22, nr. 1, p. 196–223, 2021.Vizualizare la: Site-ul editorului | Google Academic
YM Tsai, CF Chien, LC Lin și TH Tsai, „Curcuminul și nano-formularea sa: cinetica distribuției tisulare și penetrarea barierei hematoencefalice”, Jurnalul Internațional de Farmaceutică, voi . 416, nr. 1, p. 331–338, 2011.Vizualizare la: Site-ul editorului | Google Academic
B. Fonseca-Santos, MP Gremião și M. Chorilli, „Sisteme de livrare a medicamentelor bazate pe nanotehnologie pentru tratamentul bolii Alzheimer”, Jurnalul Internațional de Nanomedicină , voi. 10, p. 4981–5003, 2015.Vizualizare la: Site-ul editorului | Google Academic
Ç. Yücel, G. Şeker Karatoprak şi İ. T. Değim, „Formularea anti-îmbătrânire a etosomilor și lipozomilor încărcați cu acid rosmarinic”, Journal of Microencapsulation , voi. 36, nr. 2, p. 180–191, 2019.Vizualizare la: Site-ul editorului | Google Academic
AN Lazar, S. Mourtas, I. Youssef și colab., „Curcumin-conjugated nanoliposomes with high afinity for A β deposits: Possible applications to Alzheimer disease”, Nanomedicine: Nanotechnology, Biology and Medicine , voi. 9, nr. 5, p. 712–721, 2013.Vizualizare la: Site-ul editorului | Google Academic
M. Taylor, S. Moore, S. Mourtas și colab., „Efectul nanolipozomilor asociați cu curcumină și al ligandului lipidic funcționalizat asupra agregării peptidei Alzheimer A β ” , Nanomedicine: Nanotechnology, Biology and Medicine , voi. 7, nr. 5, p. 541–550, 2011.Vizualizare la: Site-ul editorului | Google Academic
S. Mourtas, AN Lazar, E. Markoutsa, C. Duyckaerts și SG Antimisiaris, „Nanoliposomi multifuncționali cu derivat de curcumină-lipid și funcționalitate de direcționare a creierului cu potențiale aplicații pentru boala Alzheimer”, Jurnalul European de Chimie Medicinală, voi . 80, p. 175–183, 2014.Vizualizare la: Site-ul editorului | Google Academic
S. Mourtas, M. Canovi, C. Zona și colab., „Curcumin-decorated nanoliposomes with very high afinity for amiloid- β 1-42 peptide,” Biomaterials , voi. 32, nr. 6, p. 1635–1645, 2011.Vizualizare la: Site-ul editorului | Google Academic
A. Kalani și P. Chaturvedi, „Curcumin-amorsed and curcumin-loaded exosomes: potential neural therapy”, Neural Regeneration Research , voi. 12, nr. 2, p. 205-206, 2017.Vizualizare la: Site-ul editorului | Google Academic
H. Wang, H. Sui, Y. Zheng și colab., „Exozomii amorsați cu curcumină ameliorează puternic funcția cognitivă la șoarecii AD prin inhibarea hiperfosforilării proteinei tau prin calea AKT/GSK-3 β”, Nanoscale , voi . 11, nr. 15, p. 7481–7496, 2019.Vizualizare la: Site-ul editorului | Google Academic
E. Blanco, H. Shen și M. Ferrari, „Principii de proiectare a nanoparticulelor pentru depășirea barierelor biologice în calea administrării de medicamente”, Nature Biotechnology , voi. 33, nr. 9, p. 941–951, 2015.Vizualizare la: Site-ul editorului | Google Academic
P. Henrich-Noack, D. Nikitovic, M. Neagu și colab., „Bariera hemato-encefalică și dincolo: neurofarmacologie bazată pe nano și rolul matricei extracelulare”, Nanomedicine, voi . 17, p. 359–379, 2019.Vizualizare la: Site-ul editorului | Google Academic
G. Sancini, M. Gregori, E. Salvati, I. Cambianica și F. Re, „Funcționalizarea cu peptida TAT îmbunătățește traversarea barierei hemato-encefalice in vitro a nanolipozomilor care poartă un derivat de curcumină pentru a lega peptida amiloid-β , ” Journal of Nanomedicine and Nanotechnology , voi. 4, nr. 3, p. 1–8, 2013.Vizualizare la: Site-ul editorului | Google Academic
M. Fernandes, I. Lopes, L. Magalhães et al., „Novel concept of exosome-like liposomes for the treatment of Alzheimer’s disease”, Journal of Controlled Release , voi. 336, p. 130–143, 2021.Vizualizare la: Site-ul editorului | Google Academic
SK Tiwari, S. Agarwal, B. Seth și colab., „Nanoparticulele încărcate cu curcumină induc puternic neurogeneza adultului și inversează deficitele cognitive în modelul bolii Alzheimer prin calea canonică wnt/β-catenină”, ACS Nano , voi. 8, nr. 1, p. 76–103, 2014.Vizualizare la: Site-ul editorului | Google Academic
A. Mathew, T. Fukuda, Y. Nagaoka și colab., „Curcumin loaded-PLGA nanoparticules conjugated with Tet-1 peptide for potential use in Alzheimer’s Disease”, PLoS One , voi. 7, nr. 3, articolul e32616, p. 1–10, 2012.Vizualizare la: Site-ul editorului | Google Academic
KK Cheng, PS Chan, S. Fan și colab., „Nanoparticule magnetice conjugate cu curcumină pentru detectarea plăcilor de amiloid la șoarecii cu boala Alzheimer folosind imagistica prin rezonanță magnetică (IRM)” Biomaterials, voi . 44, p. 155–172, 2015.Vizualizare la: Site-ul editorului | Google Academic
S. Fan, Y. Zheng, X. Liu și colab., „Nanoparticule PLGA-PEG încărcate cu curcumină conjugate cu peptida B6 pentru utilizare potențială în boala Alzheimer”, Drug Delivery , voi. 25, nr. 1, p. 1091–1102, 2018.Vizualizare la: Site-ul editorului | Google Academic
N. Huang, S. Lu, XG Liu, J. Zhu, YJ Wang și RT Liu, „Nanoparticulele PLGA modificate cu o peptidă care pătrunde BBB care co-livrează un inhibitor al generației β și curcumina atenuează deficitele de memorie și neuropatologia la șoarecii cu boala Alzheimer. ,” Oncotarget , vol. 8, nr. 46, p. 81001–81013, 2017.Vizualizare la: Site-ul editorului | Google Academic
F. Meng, S. Asghar, S. Gao și colab., „Un nou nanopurtător care imita LDL pentru livrarea țintită a curcuminei în creier pentru a trata boala Alzheimer”, Coloids and Surfaces, B: Biointerfaces , voi. 134, p. 88–97, 2015.Vizualizare la: Site-ul editorului | Google Academic
N. Huo, Y. Zhang, X. Jin, Y. Li și L. Zhang, „O nouă sinteză a nanoparticulelor de seleniu încapsulate nanosfere PLGA cu molecule de curcumină pentru inhibarea agregării amiloid β în boala Alzheimer”, Journal of Photochemistry and Fotobiologie. B , voi. 190, p. 98–102, 2019.Vizualizare la: Site-ul editorului | Google Academic
E. Wu, „Nanoparticule de lipide și polimeri încărcate de curcumină pentru tratamentul bolii Alzheimer”, Jurnalul Internațional de Științe Biomedicale , voi. 17, nr. 3, pp. 34–39, 2021.Vizualizați la: Google Scholar
C. Gao, Y. Wang, J. Sun și colab., „Livrarea de curcumină direcționată pe mitocondriile neuronale prin nanosisteme de inginerie biomimetică la șoarecii cu boala Alzheimer”, Acta Biomaterialia , voi. 108, p. 285–299, 2020.Vizualizare la: Site-ul editorului | Google Academic
V. Kakkar, SK Muppu, K. Chopra și IP Kaur, „Nanoparticule lipidice solide încărcate cu curcumin: o abordare eficientă de formulare pentru leziunea de reperfuzie ischemică cerebrală la șobolani”, Jurnalul European de Farmaceutică și Biofarmaceutică , voi. 85, nr. 3, p. 339–345, 2013.Vizualizare la: Site-ul editorului | Google Academic
S. Tripathi, U. Gupta, RR Ujjwal și AK Yadav, „Formularea nano-lipidice și strategiile terapeutice pentru boala Alzheimer prin cale intranazală”, Journal of Microencapsulation , voi. 38, nr. 7-8, p. 572–593, 2021.Vizualizare la: Site-ul editorului | Google Academic
AC Sintov, „Amylo Lipid nanovesicule: un sistem hibrid de amidon modificat cu lipide auto-asamblat, construit pentru livrarea directă a curcuminei de la nas la creier”, Jurnalul Internațional de Farmaceutică , voi. 588, articolul 119725, 2020.Vizualizați la: Google Scholar
N. Dhas și T. Mehta, „Livrarea intranazală a nanoparticulelor de miez PLGA / coajă decorate cu chitosan care conțin flavonoide pentru a reduce stresul oxidativ în tratamentul bolii Alzheimer”, Journal of Drug Delivery Science and Technology , voi. 61, articolul 102242, 2021.Vizualizare la: Site-ul editorului | Google Academic
L. Zhang, S. Yang , LR Wong, H. Xie și PCL Ho, „ Compararea in vitro și in vivo a nanoparticulelor poli(lactic-co-glicolic) acoperite cu chitosan încapsulat în curcumină și curcumină/hidroxipropil- β- Complexe de includere a ciclodextrinei administrate intranazal ca strategii terapeutice pentru boala Alzheimer”, Molecular Pharmaceutics , voi. 17, nr. 11, p. 4256–4269, 2020.Vizualizare la: Site-ul editorului | Google Academic
S. Sood, K. Jain și K. Gowthamarajan, „Optimizarea nanoemulsiei de curcumină pentru livrarea intranazală folosind proiectarea experimentului și evaluarea toxicității sale”, Coloids and Surfaces, B: Biointerfaces , voi. 113, p. 330–337, 2014.Vizualizare la: Site-ul editorului | Google Academic
SD Mandal, S. Mandal și J. Patel, „Eficiența de direcționare a creierului a microemulsiei mucoadezive încărcate de curcumină prin cale intranazală”, Journal of Pharmaceutical Investigation , voi. 46, nr. 2, p. 179–188, 2016.Vizualizare la: Site-ul editorului | Google Academic
KR Sharma, „Sistemul de livrare a medicamentelor Nanorobot pentru curcumină pentru o biodisponibilitate îmbunătățită în timpul tratamentului bolii Alzheimer”, Journal of Encapsulation and Adsorbtion Sciences , voi. 3, nr. 1, p. 24–34, 2013.Vizualizare la: Site-ul editorului | Google Academic
R. Giacomeli, JC Izoton, RB dos Santos, SP Boeira, CR Jesse și SE Haas, „Efectele neuroprotective ale nanocapsulelor curcuminei lipidice într-un model de boală Alzheimer indusă de peptida β-amiloid 1-42 la șoarecii femele în vârstă ” Cercetarea creierului , voi. 1721, p. 146325–146410, 2019.Vizualizare la: Site-ul editorului | Google Academic
A. Jagota și MY Reddy, „Efectul curcuminei asupra modificărilor induse de etanol în nucleul suprachiasmatic (SCN) și pineal”, Cellular and Molecular Neurobiology , voi. 27, nr. 8, p. 997–1006, 2007.Vizualizare la: Site-ul editorului | Google Academic
Y. Li, M.-M. Zhou, X.-J. Gou, L. Zhao, X.-W. Shi și W. Jia, „Efectele curcuminei asupra microbiotei intestinale a șobolanilor cu privarea de somn interval”, Chinese Traditional and Herbal Drugs , voi. 47, p. 794–798, 2016.Vizualizați la: Google Scholar
CK Saliha și ST Avadhany, „Efectul hepatoprotector și antioxidant al curcuminei asupra șobolanilor albinoși lipsiți de somn”, Journal of Pharmacy & Bioallied Sciences , voi. 9, nr. 2, p. 384–390, 2018.Vizualizare la: Site-ul editorului | Google Academic
M. Erfanizadeh, A. Noorafshan, MR Namavar, S. Karbalay-Doust și T. Talaei-Khozani, „Curcuminul previne pierderea neuronală și modificările structurale în ganglionul cervical superior (simpatic) induse de privarea cronică de somn, în modelul șobolanului ,” Cercetare biologică , voi. 53, nr. 1, p. 31, 2020.Vizualizare la: Site-ul editorului | Google Academic
Î. El, X. Chen, T. Wu, L. Li și X. Fei, „Riscul de demență la utilizatorii de benzodiazepine pe termen lung: dovezi dintr-o meta-analiză a studiilor observaționale”, Journal of Clinical Neurology, voi . 15, nr. 1, pp. 9–19, 2019.Vizualizare la: Site-ul editorului | Google Academic
V. Srinivasan, C. Kaur, S. Pandi-Perumal, GM Brown și DP Cardinali, „Melatonina și agonistul său Ramelteon în boala Alzheimer: Valoare terapeutică posibilă”, Jurnalul internațional al bolii Alzheimer, voi . 2011, articolul 741974, 2010.Vizualizare la: Site-ul editorului | Google Academic
V. Pilipenko, K. Narbute, J. Pupure, J. Rumaks, B. Jansone și V. Klusa, „Acțiunea neuroprotectoare a diazepamului la doze foarte mici și moderate la șobolani model de boala Alzheimer”, Neuropharmacology, voi . 144, p. 319–326, 2019.Vizualizare la: Site-ul editorului | Google Academic
MR Schetinger, NM Porto, MB Moretto și colab., „Noile benzodiazepine modifică activitățile acetilcolinesterazei și ATPDază”, Cercetare neurochimică , voi. 25, nr. 7, p. 949–955, 2000.Vizualizare la: Site-ul editorului | Google Academic
SA Frautschy, W. Hu, P. Kim și colab., „Inversarea antioxidantă antiinflamatoare fenolica a deficitelor cognitive induse de A β și a neuropatologiei”, Neurobiology of Aging , voi. 22, nr. 6, p. 993–1005, 2001.Vizualizare la: Site-ul editorului | Google Academic
L. Baum și A. Ng, „Interacțiunea curcuminei cu cuprul și fierul sugerează un posibil mecanism de acțiune în modelele animale ale bolii Alzheimer”, Jurnalul bolii Alzheimer , voi. 6, nr. 4, p. 367–377, 2004.Vizualizare la: Site-ul editorului | Google Academic
F. Yang, GP Lim, AN Begum și colab., „Curcumina inhibă formarea oligomerilor și fibrilelor β de amiloid , leagă plăcile și reduce amiloidul in vivo ”, The Journal of Biological Chemistry , voi. 280, nr. 7, p. 5892–5901, 2005.Vizualizare la: Site-ul editorului | Google Academic
K. Balasubramanian, „Baza orbitală moleculară pentru prevenirea bolii Alzheimer a curcuminei de curry galben”, Journal of Agricultural and Food Chemistry , voi. 54, nr. 10, p. 3512–3520, 2006.Vizualizare la: Site-ul editorului | Google Academic
M. Garcia-Alloza, LA Borrelli, A. Rozkalne, BT Hyman și BJ Bacskai, „Curcuminul etichetează patologia amiloid in vivo, perturbă plăcile existente și restaurează parțial neuritele distorsionate într-un model de șoarece Alzheimer”, Journal of Neurochemistry, voi . 102, nr. 4, p. 1095–1104, 2007.Vizualizare la: Site-ul editorului | Google Academic
SY Park, HS Kim, EK Cho și colab., „Curcuminul a protejat celulele PC12 împotriva toxicității induse de beta-amiloid prin inhibarea daunelor oxidative și a hiperfosforilării tau”, Food and Chemical Toxicology , voi. 46, nr. 8, p. 2881–2887, 2008.Vizualizare la: Site-ul editorului | Google Academic
T. Ishrat, N. Hoda, MB Khan și colab., „Ameliorarea deficitelor cognitive și neurodegenerarea prin curcumină în modelul șobolanului de demență sporadică de tip Alzheimer (SDAT),” European Neuropsychopharmacology , voi. 19, nr. 9, p. 636–647, 2009.Vizualizare la: Site-ul editorului | Google Academic
H. Awasthi, S. Tota, K. Hanif, C. Nath și R. Shukla, „Efectul protector al curcuminei împotriva streptozotocinei intracerebrale induse de afectarea memoriei și a fluxului sanguin cerebral”, Life Sciences , voi. 86, nr. 3–4, p. 87–94, 2010.Vizualizare la: Site-ul editorului | Google Academic
Y. Wang, H. Yin, J. Lou et al., „Efectele curcuminei asupra expresiei hipocampale Bax și Bcl-2 și funcției cognitive a unui model de șobolan al bolii Alzheimer”, Neural Regeneration Research , voi. 6, nr. 24, p. 1845–1849, 2011.Vizualizați la: Google Scholar
S. Dong, Q. Zeng, ES Mitchell și colab., „Curcuminul îmbunătățește neurogeneza și cogniția la șobolanii în vârstă: Implicații pentru interacțiunile transcripționale legate de creștere și plasticitate sinaptică”, PLoS One , voi. 7, nr. 2, articolul e31211, 2012.Vizualizare la: Site-ul editorului | Google Academic
CY Sun, SS Qi, P. Zhou și colab., „Validitatea neurobiologică și farmacologică a curcuminei în ameliorarea performanței de memorie a șoarecilor accelerați de senescență”, Farmacologie, biochimie și comportament , voi. 105, p. 76–82, 2013.Vizualizare la: Site-ul editorului | Google Academic
OJ Banji, D. Banji și K. Ch, „Curcumina și hesperidina îmbunătățesc cunoașterea prin suprimarea disfuncției mitocondriale și a apoptozei induse de D-galactoză în creierul șobolanului”, Food and Chemical Toxicology , voi. 74, p. 51–59, 2014.Vizualizare la: Site-ul editorului | Google Academic
L. Zhang, Y. Fang, Y. Xu și colab., „Curcuminul îmbunătățește deficitele de memorie spațială induse de β -peptida amiloid (1–42) prin calea de semnalizare BDNF-ERK”, PLoS One , voi. 10, nr. 6, articolul e0131525, 2015.Vizualizare la: Site-ul editorului | Google Academic
D. Yanagisawa, NF Ibrahim, H. Taguchi și colab., „Derivatul de curcumină cu substituție la poziția C-4, dar nu curcumina, este eficient împotriva patologiei amiloide la șoarecii APP/PS1”, Neurobiology of Aging, voi . 36, nr. 1, p. 201–210, 2015.Vizualizare la: Site-ul editorului | Google Academic
S. Doaa, C. Ismail, R. Nassra, T. Zeitoun și A. Nomair, „Modularea în aval a căii apoptotice extrinseci în demența Alzheimer indusă de streptozotocină la șobolani: eritropoietina versus curcumină”, Jurnalul European de Farmacologie, voi . 770, p. 52–60, 2016.Vizualizați la: Google Scholar
B. Vidal, RA Vázquez-Roque, D. Gnecco et al., „Tratamentul cu curcuma previne deficitul cognitiv și alterarea morfologiei neuronale în sistemul limbic al șobolanilor în vârstă”, Synapse, voi . 71, nr. 3, articolul e21952, 2017.Vizualizare la: Site-ul editorului | Google Academic
TL Moore, B. Bowley, P. Shultz et al., „Tratamentul cronic cu curcumină îmbunătățește memoria de lucru spațială, dar nu și memoria de recunoaștere la maimuțele rhesus de vârstă mijlocie”, extras de la. Gero Science , voi. 39, nr. 5–6, p. 571–584, 2017.Vizualizare la: Site-ul editorului | Google Academic
S. Singh și P. Kumar, „Potențialul neuroprotector al curcuminei în combinație cu piperina împotriva deficitului motor indus de 6-hidroxi dopamină și al modificărilor neurochimice la șobolani”, Inflammopharmacology , voi. 25, nr. 1, p. 69–79, 2017.Vizualizare la: Site-ul editorului | Google Academic
JR Sundaram, CP Poore, NHB Sulaimee et al., „Curcuminul ameliorează neuroinflamația, neurodegenerarea și deficitele de memorie în modelul de șoarece transgenic p 25 care poartă semnele distinctive ale bolii Alzheimer”, Journal of Alzheimer’s Disease, voi . 60, nr. 4, p. 1429–1442, 2017.Vizualizare la: Site-ul editorului | Google Academic
LY Shi, L. Zhang, H. Li și colab., „Efectele protectoare ale curcuminei asupra neurotoxicității induse de acroleină în celulele hipocampale de șoarece HT22”, Pharmacological Reports , voi. 70, nr. 5, p. 1040–1046, 2018.Vizualizare la: Site-ul editorului | Google Academic
B. Teter, T. Morihara, GP Lim și colab., „Curcuminul restabilește expresia imună înnăscută a genei de risc pentru boala Alzheimer pentru a ameliora patogeneza Alzheimer”, Neurobiology of Disease , voi. 127, p. 432–448, 2019.Vizualizare la: Site-ul editorului | Google Academic
L. Lin, C. Li, D. Zhang, M. Yuan, CH Chen și M. Li, „Efectele sinergice ale berberinei și curcuminei asupra îmbunătățirii funcției cognitive într-un model de șoarece cu boala Alzheimer”, Neurochemical Research, voi . 45, nr. 5, p. 1130–1141, 2020.Vizualizare la: Site-ul editorului | Google Academic
L. Baum, CWK Lam, SK-K. Cheung și colab., „Trial clinic pilot de șase luni, controlat cu placebo, dublu-orb, al curcuminei la pacienții cu boala Alzheimer”, Journal of Clinical Psychopharmacology , voi. 28, nr. 1, p. 110–113, 2008.Vizualizare la: Site-ul editorului | Google Academic
JM Ringman, SA Frautschy, E. Teng și colab., „Curcumină orală pentru boala Alzheimer: tolerabilitate și eficacitate într-un studiu randomizat, dublu orb, controlat cu placebo de 24 de săptămâni”, Alzheimer’s Research & Therapy, voi . 4, nr. 5, p. 43, 2012.Vizualizare la: Site-ul editorului | Google Academic
KH Cox, A. Pipingas și AB Scholey, „Investigarea efectelor curcuminei lipide solide asupra cogniției și dispoziției într-o populație în vârstă sănătoasă”, Journal of Psychopharmacology , voi. 29, nr. 5, p. 642–651, 2015.Vizualizare la: Site-ul editorului | Google Academic
SR Rainey-Smith, BM Brown, HR Sohrabi et al., „Curcumină și cogniție: un studiu randomizat, controlat cu placebo, dublu-orb al adulților în vârstă care locuiesc în comunitate”, The British Journal of Nutrition, voi . 115, nr. 12, p. 2106–2113, 2016.Vizualizare la: Site-ul editorului | Google Academic
GW Small, P. Siddarth, ZP Li și colab., „Efectele memoriei și creierului amiloid și Tau ale unei forme biodisponibile de curcumină la adulții non-dementați: un studiu dublu-orb, controlat cu placebo de 18 luni”, The American Journal de Psihiatrie Geriatrică , voi. 26, nr. 3, p. 266–277, 2018.Vizualizare la: Site-ul editorului | Google Academic
SC Gupta, S. Patchva și BB Aggarwal, „Rolurile terapeutice ale curcuminei: lecții învățate din studiile clinice”, The AAPS Journal , voi. 15, nr. 1, p. 195–218, 2013.Vizualizare la: Site-ul editorului | Google Academic
C. Mancuso și E. Barone, „Curcuminul în practica clinică: mit sau realitate?” Trends in Pharmacological Sciences , voi. 30, nr. 7, p. 333-334, 2009.Vizualizare la: Site-ul editorului | Google Academic
CE Goodpasture și FE Arrighi, „Efectele condimentelor alimentare asupra ciclului celular și morfologiei cromozomilor celulelor de mamifere _in vitro_ cu referire specială la turmeric”, Food and Cosmetics Toxicology , voi. 14, nr. 1, p. 9–14, 1976.Vizualizare la: Site-ul editorului | Google Academic
P. Urbina-Cano, L. Bobadilla-Morales, MA Ramírez-Herrera și colab., „Daune ADN în limfocitele de șoarece expuse la curcumină și cupru”, Journal of Applied Genetics , voi. 47, nr. 4, p. 377–382, 2006.Vizualizare la: Site-ul editorului | Google Academic
RA Sharma, SA Euden, SL Platton și colab., „Testul clinic de fază I cu curcumină orală: biomarkeri ai activității sistemice și conformității”, Clinical Cancer Research , voi. 10, nr. 20, p. 6847–6854, 2004.Vizualizare la: Site-ul editorului | Google Academic
H. Hanai, T. Iida, K. Takeuchi și colab., „Terapia de întreținere a curcuminei pentru colita ulceroasă: studiu randomizat, multicentric, dublu-orb, controlat cu placebo”, Clinical Gastroenterology and Hepatology, voi . 4, nr. 12, p. 1502–1506, 2006.Vizualizare la: Site-ul editorului | Google Academic
B. Lal, A. Kapoor, P. Agrawal, OP Asthana și RC Srimal, „Rolul curcuminei în pseudotumorile orbitale inflamatorii idiopatice”, Phytotherapy Research , voi. 14, nr. 6, p. 443–447, 2000.Vizualizare la: Site-ul editorului | Google Academic
P. Pinsornsak și S. Niempoog, „Eficacitatea extractului de Curcuma longa L. ca terapie adjuvantă în osteoartrita primară a genunchiului: un studiu de control randomizat”, Jurnalul Asociației Medicale din Thailanda , voi. 95, p. 51–58, 2012.Vizualizați la: Google Scholar
JL Ryan, CE Heckler, M. Ling și colab., „Curcumină pentru dermatita radiațiilor: un studiu clinic randomizat, dublu-orb, controlat cu placebo, pe treizeci de pacienți cu cancer de sân”, Radiation Research , voi. 180, nr. 1, p. 34–43, 2013.Vizualizare la: Site-ul editorului | Google Academic
AR Rahimnia, Y. Panahi, G. Alishiri, M. Sharafi și A. Sahebkar, „Impact al suplimentării cu curcuminoizi asupra inflamației sistemice la pacienții cu osteoartrită a genunchiului: constatări dintr-un studiu randomizat, dublu-orb, controlat cu placebo,” Drug research , vol. 65, nr. 10, p. 521–525, 2015.Vizualizare la: Site-ul editorului | Google Academic
MA Kuriakose, K. Ramdas, B. Dey și colab., „Un studiu randomizat de fază iib, dublu-orb, controlat cu placebo, al curcuminei în leucoplazia orală”, Cancer Prevention Research , voi. 9, nr. 8, p. 683–691, 2016.Vizualizare la: Site-ul editorului | Google Academic
Licența de atribuire Creative Commons , care permite utilizarea, distribuirea și reproducerea fără restricții pe orice mediu, cu condiția ca lucrarea originală să fie citată corespunzător.
Peste 80% dintre pacienții afectați de cancer dezvoltă dureri legate de cancer, una dintre cele mai de temut consecințe din cauza naturii sale insolubile, în special în stadiul terminal al bolii. Recomandări recente bazate pe dovezi privind medicina integrativă pentru gestionarea durerii cauzate de cancer subliniază rolul produselor naturale. Prezenta revizuire sistematică și meta-analiză își propune să evalueze pentru prima dată eficacitatea aromoterapiei în durerea canceroasă în studii clinice cu design diferit, conform celor mai actualizate articole de raportare preferate pentru recenzii sistematice și meta-analize (PRISMA) 2020 – recomandări. Căutarea preia 1002 înregistrări în total. Douăsprezece studii sunt incluse și șase sunt eligibile pentru meta-analiză.Prezentul studiu demonstrează eficacitatea semnificativă a utilizării uleiurilor esențiale în reducerea intensității durerii asociate cu cancerul (p < 0,00001),, evidențiind necesitatea unor studii clinice mai devreme, mai omogene și concepute corespunzător. Este nevoie de un corp de dovezi cu certitudine bună pentru gestionarea eficientă și sigură a durerii legate de cancer, folosind uleiuri esențiale, prin stabilirea unei căi pas cu pas de la preclinic la clinic pentru a oferi o bază rațională pentru utilizarea clinică în oncologia integrativă. Înregistrare PROSPERO: CRD42023393182.
Termenul „durere de cancer” caracterizează cel mai bine natura multidimensională și cu mai multe fațete a durerii, deoarece include domeniile fizice, psihosociale și ale calității vieții, adesea concomitent cu așa-numitul grup de simptome de cancer, inclusiv anxietatea, depresia și tulburările de somn. 1 , 2 ]. Problema prezentată de durerea canceroasă, din cauza tratamentului chirurgical și chimioterapic, alături de caracteristicile specifice tumorii, este o chestiune de urgență datorită legăturii strânse cu reducerea calității generale a vieții și numărul tot mai mare de pacienți afectați de cancer. De fapt, îmbunătățirea sistemelor de detecție precoce și îmbătrânirea globală provoacă o creștere continuă a supraviețuitorilor de cancer, cu aproximativ 18 milioane de persoane cu antecedente de cancer până la începutul anului 2022 în Statele Unite [ 3 ]]. Toate grupurile de simptome de cancer pot împărtăși baze biologice comune, probabil datorită implicării unor răspunsuri inflamatorii și imune intense și mocnite provocate de mediul de patogeneză a cancerului, dar și de tratament [4 ] .
Peste 70–80% dintre pacienții afectați de tumori sunt raportați că suferă de durere canceroasă în meta-analizele referitoare la prevalența durerii canceroase într-o perioadă de 40 de ani [ 5 , 6 ]; această durere este insuportabilă până la 33% dintre pacienți [ 7 ] și atinge rata de 95% la pacienții afectați de boală avansată (vezi [ 8 ]), făcând durerea canceroasă o problemă majoră în managementul pacienților oncologici. Mai mult, cea mai recentă revizuire sistematică care se ocupă de prevalența durerii la supraviețuitorii cancerului datează din 2022 [ 9]]: subliniază că prevalența durerii la supraviețuitorii unei tumori solide care au terminat tratamentul cu cel puțin 3 luni mai devreme este de 47%, cu o eterogenitate de 98,99% între studii. Aceste date susțin importanța tratamentului durerii canceroase pentru managementul și îmbunătățirea calității vieții pacienților cu cancer.
În plus, durerea canceroasă este adesea persistentă și cronică [ 10 ], cu caracteristici revoluționare. Reprezintă una dintre cele mai de temut consecințe ale cancerului datorită naturii sale insolubile în stadiul terminal al bolii, determinând ca aproximativ 25% dintre pacienți să moară cu dureri semnificative (vezi [8] ) .
Durerea cronică, care include adesea caracteristici inflamatorii și neuropatice, așa cum apar în durerea canceroasă, este unul dintre cele mai comune motive pentru a ajunge la observația clinică [ 11 ]. Durerea de cancer include sindroame legate de tratamente chirurgicale sau de chimioterapie/radioterapie/hormonoterapia [ 12 ]. De fapt, durerea neuropatică, din cauza unei leziuni sau boli a sistemului somatosenzorial [ 13 ], poate apărea după proceduri chirurgicale [ 14 ] sau leziuni [ 15 ] și, de asemenea, accident vascular cerebral [ 16 ], dar și neuropatii și cefalee [ 17 ] rezultând de la tratamente chimioterapeutice care provoacă neuropatie periferică indusă de chimioterapie (CIPN), care este adesea subrecunoscută și subtratată [ 18], 19 ].
În plus, tratamentul durerii cronice la pacienții în vârstă este complicat de: lipsa de informații despre utilizarea adecvată și doza de analgezice din cauza excluderii acestor pacienți din studiile clinice [20], în special pentru tratamentele anti-migrenă [ 21 , 22 , 23] ]; polifarmacie [ 24 ]; diferențe fiziologice [ 25 , 26 ]. Această lipsă de informare se datorează practicii de excludere a pacienților în vârstă. Utilizarea uleiurilor esențiale dotate cu proprietăți analgezice dovedite poate fi cea mai sigură opțiune în cadrul medicinei integrative în oncologie.
Un ghid recent (2022), elaborat pentru a oferi clinicienilor recomandări bazate pe dovezi despre abordări integrative pentru gestionarea durerii la pacienții cu cancer, raportează rolul produselor naturale [27 ] . În special, aromaterapia s-a dovedit a avea niveluri scăzute de dovezi pentru tratamentul CIPN, raportând doar cinci studii randomizate, controlate [ 27 ]. O revizuire sistematică anterioară care a evaluat eficacitatea terapiilor complementare și alternative în durerea canceroasă a evidențiat deja calitatea scăzută a cercetării asupra suplimentelor pe bază de plante și deficitul de studii randomizate, controlate în acest domeniu [28 ] .
Cercetările preclinice din grupul nostru au construit rațiunea pentru traducerea clinică a uleiului esențial de bergamotă (BEO) pentru administrarea continuă ca aromoterapie și pentru aplicarea transdermică a unei forme farmaceutice, nanotehnologice, concepute, numită NanoBEO eliberată de un dozator fără aer [ 29 , 30 , 31 , 32 , 33 , 34 , 35 , 36 , 37 ]. Deoarece BEO și-a dovedit eficiența analgezică atât în modelele de durere inflamatorie, cât și în cele neuropatice, de încredere clinică, ar putea reprezenta un candidat pentru tratamentul durerii canceroase.
Scopul prezentei revizuiri sistematice și meta-analizei este de a evalua pentru prima dată eficacitatea aromoterapiei în reducerea durerii canceroase prin studii clinice de orice tip, urmând cele mai actualizate articole de raportare preferate pentru revizuiri sistematice și metaanalize ( PRISMA) Recomandări 2020 [ 38 ]. Mai mult, informațiile referitoare la tipul de uleiuri esențiale utilizate și eficacitatea pot oferi puncte de reflecție despre posibilele mecanisme implicate în controlul durerii canceroase. Prezenta revizuire sistematică este înregistrată în Registrul internațional prospectiv al recenziilor sistematice (PROSPERO) de Institutul Național de Cercetare în Sănătate (NIHR) cu numărul CRD42023393182.
Căutarea, extragerea și selecția rezultatelor se efectuează în acord cu cele mai recent actualizate articole de raportare preferate pentru revizuiri sistematice și meta-analize (PRISMA) 2020 recomandări [ 38 , 38 , 39 , 40 .]. Întrebarea de cercetare este formulată ca o întrebare PICOS (participanți/populație, intervenții, comparații, rezultate și design de studiu). Participanții sunt pacienți afectați de dureri de cancer. Intervenția constă în uleiuri esențiale administrate în orice doză și cale. Studiile sunt considerate a fi eligibile dacă evaluează eficacitatea intervenției față de comparatorul constând în placebo/fără tratament sau control activ, adică orice medicament aprobat pentru tratamentul durerii. Modelele de studiu care trebuie incluse sunt studii clinice prospective și retrospective. Rezultatul principal constă în reducerea durerii. Sunt excluse studiile preclinice in vivo și in vitro, recenziile, capitolele de carte și comunicările și lucrările congresului. Protocolul a fost stabilit anterior căutării literaturii și înregistrat în PROSPERO (CRD42023393182). Titlurile și rezumatele și, ulterior, textul integral al studiilor preluate, sunt verificate de doi membri independenți ai comisiei de revizuire pe baza criteriilor de includere și excludere stabilite a priori. Referințele celor mai semnificative lucrări sunt inspectate pentru a evita lipsa unor studii suplimentare. Orice dezacord se rezolvă prin obținerea consensului prin metoda Delphi [41 ] sau prin consultarea unui al treilea membru al echipei.
2.2. Surse de informare
PubMed/MEDLINE, Scopus și WOS sunt inspectate pentru studiile revizuite de colegi publicate de la începutul bazei de date și până în prezent, fără restricții de dată, până la data ultimei căutări din 3 februarie 2023. Sunt incluse numai articolele publicate în limba engleză. După examinarea titlului și a rezumatelor, articolele care nu sunt disponibile în text complet sunt excluse. Doi membri independenți ai comitetului de revizuire caută înregistrări care se potrivesc cu șirurile strategiei.
2.3. Strategia de căutare
Următorii termeni medicali și titluri de subiecte (MeSH) sunt utilizați în combinație în cadrul șirurilor de căutare: „Cancer”; „Oncologic”; „Tumo(u)r”; „Durere”; „Uleiuri esentiale”; „Aromaterapie”; „Medicina integrativă”. PubMed/MEDLINE sunt căutați pentru termeni MeSH, Scopus pentru Titlul articolului, Rezumat, Cuvinte cheie și WOS pentru toate câmpurile. Căutarea are caracteristicile unei strategii de căutare cu sensibilitate mare/rechemare, păstrând precizia [ 42]. Rândurile și ortografia șirurilor de căutare, capacitatea căutării de a acoperi toate aspectele cele mai relevante și acuratețea de a răspunde la întrebarea PICOS sunt verificate de un autor diferit (revizor) de cei doi care caută în bazele de date în mod independent (solicitanți) în acord. cu ghidul bazat pe dovezi pentru Peer Review of Electronic Search Strategies (PRESS) pentru revizuiri sistematice (SRs) [ 42 , 43 ].
2.4. Selectarea studiilor
Doi autori evaluează în mod independent eligibilitatea rezultatelor obținute, pentru a minimiza riscul de a pierde înregistrările relevante. Duplicatele sunt eliminate de software-ul de gestionare a referințelor (EndNote X7, Clarivate, Londra, Marea Britanie). Titlul și rezumatul și ulterior textul complet sunt ecranizate.
2.5. Sinteza datelor, evaluarea riscului de părtinire și evaluare critică
Sinteza rezultatelor este realizată conform ghidurilor Cochrane Consumers and Communication Review Group [ 44 ]. Riscul de părtinire (RoB) și calitatea/certitudinea [ 45 ] a corpului de dovezi sunt evaluate independent de doi membri ai comitetului de revizuire conform declarației PRISMA 2020 [ 46 ] folosind: instrumentul Cochrane de risc de părtinire revizuit RoB2 pentru randomizați. studii clinice [ 47 ]; instrumentul ROBINS-I [ 48 ] pentru studii nerandomizate. Vizualizarea riscului de evaluare a părtinirii este produsă cu instrumentul de vizualizare Cochrane robvis [ 49 ].
2.6. Analiză statistică și măsuri de efect
Cochrane Review Manager 5.4.1 (RevMan5.4.1; Copenhaga: Nordic Cochrane Center, The Cochrane Collaboration) este utilizat pentru a măsura intervalele de încredere de 95% (CI) sau diferențele medii standardizate (SMD) și varianța inversă pentru variabile dihotomice și continue , respectiv. Lipsa de studii eligibile pentru analiză nu permite planificarea unei analize de sensibilitate, limitând analiza primară la studii cu risc scăzut de părtinire sau în urma analizei subgrupurilor sau a meta-regresiei bazate pe stratificarea studiilor. Eterogenitatea rezultatelor obținute este calculată prin modelul cu efect aleatoriu [ 50 ] și valoarea Higgins I 2 [ 51 ], în timp ce testul de regresie liniară al lui Egger [ 52 ] pentru asimetria diagramei pâlnie [ 53 ]], ajustată prin metoda „trim and fill” [ 54 ], este utilizată pentru a evalua distorsiunea publicării.
Bazele de date căutate sunt PubMed/MEDLINE, Scopus și Web of Science (WOS) de la începutul lor până la data ultimei căutări, adică 3 februarie 2023. Căutarea preia 1002 înregistrări totale. În special, 131 de înregistrări sunt preluate de la PubMed/MEDLINE, 563 de la Scopus și 308 de la WOS. Nu sunt identificate studii suplimentare prin screening-ul listei de referință. Eliminarea dublelor duce la 563 de studii rămase. Screeningul titlului și al rezumatelor conduce la excluderea înregistrărilor care nu îndeplinesc criteriile de includere (pentru diferite rezultate investigate sau design de studiu, de exemplu, studii preclinice in vitro și in vivo, rapoarte de caz, studii clinice calitative, anchete, recenzii, capitole de carte, congres rezumate, lucrări etc.), precum și pentru intervenția utilizată. De fapt, unele înregistrări pot părea că îndeplinesc criteriile de includere, dar intervenția constă în extracte sau componente ale uleiurilor esențiale, și nu uleiuri esențiale sau aromoterapie. De exemplu, studiul lui Hasheminasab et al., 2020 [55 ] este exclus deoarece tratamentul pe bază de plante utilizat nu este un ulei esențial. În plus, în evidențele lui Aghamohammadi și colaboratori (2018) [ 56 ] și ale lui Arantes et al., 2021 [ 57 ], intervenția conține un extract, nu un ulei esențial. Studiile lui Czakert [ 58 ] și ale lui Ho [ 59 ] și colegii de muncă sunt excluse deoarece efectuează o analiză calitativă. După ecranizarea titlului și a rezumatului, au rămas 22 de înregistrări. În special, lucrările lui Oyston și McGee, 2012 [ 60 ], ale lui Lee și Park, 2018 [ 61 ] și ale lui Cheong și colab., 2022 [ 62 ] nu sunt disponibile în textul integral; prin urmare, acestea sunt excluse. Articolele lui Xiao și colaboratori (2018) [63 ] și din Chang, 2008 [ 64 ] sunt excluse din cauza faptului că sunt în limba chineză. Înregistrările lui Nekuzad și colab., 2012 [ 65 ] și studiul pilot cvasi-randomizat, controlat, realizat de Yayla și Ozdemir în 2019 [ 66 ] nu sunt eligibile deoarece articolele nu sunt disponibile în text integral. Studiul lui Corbin și colab., 2009 [ 67 ] nu este eligibil pentru includerea în analiză, deoarece este o scrisoare către editor care raportează despre utilizarea medicinei complementare și alternative, dar nu aromoterapiei sau uleiurilor esențiale în mod specific. Lucrarea lui Ovayolu et al., 2014 [ 68 ] nu poate fi inclusă în analiză, deoarece durerea nu este un punct final al studiului. Studiul lui Elhadad și al colegilor (2022) [ 69] nu poate fi inclus deoarece intervenția se bazează pe un gel de extract alcoolic de mușețel. Screeningul textului complet lasă 12 rezultate care urmează să fie incluse în analiză. Procesul de extragere a studiilor este ilustrat înfigura 1.
Procesul de extragere a rezultatelor conform Diagramei de flux Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) 2020 produs cu aplicația Shiny bazată pe web [ 70 ].
3.2. Sinteza datelor, evaluarea critică și meta-analiză
Studiul Blackburn et al., 2017 [ 71] este un studiu randomizat, încrucișat, wash-out (design ales pentru a ține cont de variabilitatea inter-individuală față de simptomele induse de terapie) în care 50 de pacienți cu leucemie mielogenă acută supuși chimioterapiei primesc aromaterapie. Lavanda este parfumul selectat cel mai des în acest studiu, urmat de mentă și apoi de mușețel. Randomizarea folosește un tabel generat de computer, iar caracteristicile de referință nu sugerează îngrijorare; prin urmare, nu există niciun risc de părtinire în ceea ce privește domeniul 1, adică randomizarea, folosind instrumentul RoB 2 pentru studiile încrucișate în funcție de situația în care datele din ambele perioade sunt analizate în mod corespunzător, luând în considerare împerecherea observațiilor în cele două. perioade pentru fiecare individ. În prima săptămână, pacienții sunt randomizați la placebo sau aromoterapie, urmate de eliminarea în a doua săptămână, iar apoi placebo sau aromaterapie în a treia săptămână. Douăzeci și cinci de pacienți sunt supuși mai întâi aromoterapie și 28 de pacienți primesc placebo. Cu toate acestea, o anumită părtinire care decurge din efectele de transfer (domeniul S) este înregistrată deoarece ar trebui incluse interacțiunile intervenției în funcție de perioadă de timp. Doar trei pacienți nu primesc intervenții conform planului; prin urmare, un număr sub 10% din eșantionul total, adică domeniul 2 de abateri de la intervenția intenționată poate fi considerat imparțial. Din cei cincizeci și trei de pacienți înrolați, 50 au finalizat studiul și 53 de pacienți au demonstrat o putere de 91,8% conform calculului puterii eșantionului; prin urmare, nu există nicio îngrijorare în ceea ce privește părtinirea din cauza lipsei datelor privind rezultatele (domeniul 3). În plus, nu este detectată nicio părtinire în măsurarea rezultatului (domeniul 4), deoarece sistemele de măsurare a rezultatului sunt adecvate utilizând Scala de Evaluare a Simptomelor Edmonton – Revizuită cu Scala Analogică Vizuală (VAS). Nu apare niciun risc de părtinire în selectarea rezultatelor raportate (domeniul 5). Evaluarea părtinirii este raportată înFigura 2. În acest studiu, aromaterapia oferă ameliorarea oboselii, somnolenței, lipsei poftei de mâncare, depresiei, anxietății și bunăstării, dar nu o reducere a durerii.
Evaluarea riscului de părtinire ca parcelă de semafor ( a ) și diagrame cu bare ponderate ( b ). Vizualizarea riscului de evaluare a părtinirii este produsă cu instrumentul de vizualizare Cochrane robvis [ 49 , 71 , 72 , 73 , 74 , 75 , 76 , 77 , 78 , 79 , 80 , 81 , 82 ] și evaluată în conformitate cu revizuirile. Instrumentul Cochrane pentru riscul de părtinire RoB2 pentru studiile clinice randomizate și instrumentul Riscul de părtinire în studiile nerandomizate — de intervenții (ROBINS-I) pentru studiile nerandomizate.
Studiul Blackburn și colaboratorii efectuat în 2021 [ 72] implică pacienți afectați de cancer de col uterin avansat, supuși brahiterapiei pentru administrarea de doze mari de radiații la tumora primară și doze de economisire la organele cu risc. Această procedură este invazivă, provocând durere și anxietate. Prin urmare, acest studiu randomizat, controlat își propune să evalueze eficacitatea aromoterapiei cu levănțică, lămâie sau mentă și reflexologie la picioare pentru a reduce durerea și anxietatea la pacienții care primesc brahiterapie pentru cancer de col uterin. Chiar și fără analiză statistică, ca și în studiul lui Blackburn și colegii de muncă din 2017, caracteristicile de bază nu sugerează o părtinire pentru evaluarea durerii, în afară de ceea ce se întâmplă pentru elementul care contabilizează consumul de opioide înainte de studiu, utilizat de o medie de nouă pacienții din grupul de intervenție și cu o mediană de patru în grupul de control (diferență de 20%, după cum au raportat autorii). Această problemă este gestionată prin măsurile repetate în cadrul subiectului, pentru a ține seama de posibila corelație a scorurilor durerii de la același subiect. Prin urmare, deși pacienții sunt randomizați folosind software-ul GraphPad, este raportată îngrijorarea în ceea ce privește părtinirea randomizării (domeniul 1). Eșantionul de 41 de pacienți îndeplinește calculul de dimensiune a eșantionului efectuat, conform căruia dimensiunea minimă a eșantionului a fost stabilită la 40 de pacienți pentru a atinge puterea de cel puțin 80%. Prin urmare, în ciuda celor șase participanți care nu au reușit să completeze datele pentru cele cinci fracțiuni complete de tratament, există un anumit risc de părtinire în domeniul 2, dar nicio îngrijorare în ceea ce privește părtinirea din cauza lipsei datelor privind rezultatul (domeniul 3). Durerea este măsurată folosind o scală de evaluare numerică (NRS). Deoarece sistemul de măsurare constând în NRS este adecvat, nu este raportat niciun risc de părtinire în măsurarea rezultatului (domeniul 4). Nu este semnalat niciun risc de părtinire în selectarea rezultatelor raportate (domeniul 5). Efectul intervenției, după reflexoterapie, este semnificativ statistic (p < 0,0001).
Studiul realizat de Deng et al., 2021 [ 73 ] se ocupă de efectul aromaterapiei perioperatorii, cu sau fără terapie prin muzică, asupra nivelului de durere și anxietate la femei după operația de cancer la sân. Un total de 160 de pacienți cu cancer de sân sunt repartizați aleatoriu într-un raport de 1:1:1:1 pentru a primi îngrijire obișnuită, aromoterapie, terapie muzicală sau terapie combinată în perioadele perioperatorii. Măsurarea durerii constă în VAS; astfel, nu este raportat niciun risc de părtinire în măsurarea rezultatului (domeniul 4). Interesant, nivelurile de interleukină (IL) 6 sunt evaluate ca biomarker. Natura deschisă a studiului, fără orbire, ridică o anumită îngrijorare în ceea ce privește riscul de părtinire pentru alocare (domeniul 1). Pe de altă parte, diferențele dintre caracteristicile de bază sunt comparate prin χ 2teste pentru variabile categorice, care nu arată nicio diferență semnificativă în caracteristicile demografice și clinice la momentul inițial. Un număr de 196 de pacienți este examinat pentru eligibilitate, iar 160 de participanți sunt alocați aleatoriu protocolului: deși numărul de pacienți este mare pentru studiile de aromoterapie, nu este raportat niciun calcul al puterii eșantionului. Datele nu sugerează părtinire în domeniul 2, 3, 4 și 5. Conform rezultatelor, la 4 ore după extubarea traheală după intervenție chirurgicală, terapia combinată este cea mai eficientă opțiune și este mai eficientă decât aromaterapia singură; aceste rezultate sunt paralele cu reducerea nivelului de IL-6. Din păcate, alegerea uleiului esențial este un factor important care nu este explorat în studiul de față.
Cercetarea efectuată de Ha et al., 2022 [ 74] constă într-un studiu de fază II, randomizat, încrucișat, care își propune să evalueze efectele treajului limfatic aromă asupra durerii la pacienții cu cancer de sân care urmează să primească chimioterapie pe bază de taxani. Tămâia este folosită ca ulei esențial și uleiul de migdale dulci ca ulei purtător. Tresajul limfatic cu aromă se aplică în plus față de îngrijirea standard pentru durere, în comparație cu îngrijirea standard numai, în 4 zile consecutive din ziua următoare administrării taxanului. Diferențele de bază de vârstă ridică unele preocupări, în ciuda randomizării și alocării automate. Există o perioadă de două săptămâni de eliminare, importantă în studiile clinice încrucișate și, în mod adecvat, evaluările perioadei/ciclului sunt raportate, de asemenea, separat pentru ciclul 1, presupunând absența efectului de transfer, așa cum se precizează de autorii (prejudecată scăzută în domeniul S). În plus, se calculează efectul intervenției tratamentului și efectul perioadei dintre ciclurile 1 și 2. Pentru a obține o putere de 80% cu un nivel de semnificație de 5%, 37 de pacienți din fiecare grup, deci un total de 74 de pacienți, sunt înrolați în studiu. Pe parcursul studiului, nouă pacienți refuză să continue intervenția; astfel, 65 de pacienți finalizează studiul pe protocol. Măsura rezultatului este înregistrată prin VAS. În consecință, nu este raportat niciun risc de părtinire în măsurarea rezultatului (domeniul 4). În plus, nu este detectat niciun risc de părtinire în selectarea rezultatelor raportate (domeniul 5). Scorurile de vârf ale durerii între opțiunile de tratament în ambele cicluri arată 5,05 ± 2,56 pentru intervenție și 5,28 ± 2,45 pentru îngrijirea standard. Prin urmare, nu există nicio diferență semnificativă în scorul durerii ( Pentru a obține o putere de 80% cu un nivel de semnificație de 5%, 37 de pacienți din fiecare grup, deci un total de 74 de pacienți, sunt înrolați în studiu. Pe parcursul studiului, nouă pacienți refuză să continue intervenția; astfel, 65 de pacienți finalizează studiul pe protocol. Măsura rezultatului este înregistrată prin VAS. În consecință, nu este raportat niciun risc de părtinire în măsurarea rezultatului (domeniul 4). În plus, nu este detectat niciun risc de părtinire în selectarea rezultatelor raportate (domeniul 5). Scorurile de vârf ale durerii între opțiunile de tratament în ambele cicluri arată 5,05 ± 2,56 pentru intervenție și 5,28 ± 2,45 pentru îngrijirea standard. Prin urmare, nu există nicio diferență semnificativă în scorul durerii ( Pentru a obține o putere de 80% cu un nivel de semnificație de 5%, 37 de pacienți din fiecare grup, deci un total de 74 de pacienți, sunt înrolați în studiu. Pe parcursul studiului, nouă pacienți refuză să continue intervenția; astfel, 65 de pacienți finalizează studiul pe protocol. Măsura rezultatului este înregistrată prin VAS. În consecință, nu este raportat niciun risc de părtinire în măsurarea rezultatului (domeniul 4). În plus, nu este detectat niciun risc de părtinire în selectarea rezultatelor raportate (domeniul 5). Scorurile de vârf ale durerii între opțiunile de tratament în ambele cicluri arată 5,05 ± 2,56 pentru intervenție și 5,28 ± 2,45 pentru îngrijirea standard. Prin urmare, nu există nicio diferență semnificativă în scorul durerii ( 65 de pacienți finalizează studiul pe protocol. Măsura rezultatului este înregistrată prin VAS. În consecință, nu este raportat niciun risc de părtinire în măsurarea rezultatului (domeniul 4). În plus, nu este detectat niciun risc de părtinire în selectarea rezultatelor raportate (domeniul 5). Scorurile de vârf ale durerii între opțiunile de tratament în ambele cicluri arată 5,05 ± 2,56 pentru intervenție și 5,28 ± 2,45 pentru îngrijirea standard. Prin urmare, nu există nicio diferență semnificativă în scorul durerii ( 65 de pacienți finalizează studiul pe protocol. Măsura rezultatului este înregistrată prin VAS. În consecință, nu este raportat niciun risc de părtinire în măsurarea rezultatului (domeniul 4). În plus, nu este detectat niciun risc de părtinire în selectarea rezultatelor raportate (domeniul 5). Scorurile de vârf ale durerii între opțiunile de tratament în ambele cicluri arată 5,05 ± 2,56 pentru intervenție și 5,28 ± 2,45 pentru îngrijirea standard. Prin urmare, nu există nicio diferență semnificativă în scorul durerii (p = 0,368) sau se raportează toxicitate.
Studiul lui Ilter și colab., 2019 [ 75 ] este un studiu controlat, nerandomizat, care evaluează efectul aromaterapiei inhalatoare asupra durerii invazive în timpul inserării cateterului portuar la pacienții cu cancer. Prin urmare, există unele preocupări în ceea ce privește distorsiunea de randomizare, deși caracteristicile inițiale ale pacienților sunt similare ( p > 0,05), cu excepția nivelului educațional ( p.= 0,047). Amestecul aromat se obține prin diluarea uleiului de portocale, mușețel și lavandă în raport de 1:1:1 în 70 ml apă distilată. Conform calculului puterii eșantionului, numărul minim de pacienți necesar este de 17, iar studiul este finalizat de 30 de pacienți pentru grupul de intervenție și 30 de pacienți pentru grupul de control. În plus, niciun pacient nu solicită întreruperea intervenției sau decide să nu continue studiul. Este important să se ia în considerare faptul că evaluările grupului de control sunt efectuate în mod corespunzător înaintea grupului de intervenție pentru a preveni participanții din grupul de control să fie expuși la orice reziduu de aromoterapie. În plus, sunt evaluate semnele vitale înainte, în timpul și după procedură și respectarea procedurii.
Studiul lui Izgu et al., 2019 [ 76] este un studiu pilot deschis, cu grup paralel, cvasi-randomizat (randomizare stratificată), controlat, care ridică un risc ridicat de părtinire în ceea ce privește randomizarea și alocare. Acest studiu își propune să investigheze efectul masajului cu aromoterapie asupra durerii CIPN la pacienții cu cancer tratați cu oxaliplatin. Este de remarcat faptul că Autorii raportează că un control placebo nu este efectuat din cauza mirosului caracteristic al amestecului de uleiuri esențiale (mentă, mușețel și rozmarin amestecate în proporție de 1:1:1 la 1,5% în 50 ml de ulei de cocos), prin urmare, folosind îngrijirea de rutină ca comparator. Se raportează că amestecul de uleiuri esențiale este depozitat în sticle de sticlă de 50 ml rezistente la lumină și ermetice. În mod corespunzător, durerea neuropatică este evaluată folosind Douleur Neuropathique 4 Întrebări (DN4), fără risc de părtinire în domeniul 4. Conform calculului eșantionului de putere efectuat folosind pachetul software G*power, versiunea 3.1.7, N = 22 este suficient pentru fiecare grup din acest studiu. Patruzeci și șase de pacienți sunt repartizați cvasialeatoriu în grupurile de intervenție (N = 22) și de control (N = 24), similare în ceea ce privește caracteristicile inițiale (p = 0,627). În perioada de studiu, în grupul de intervenție doi pacienți abandonează și în grupul de control, patru pacienți abandonează și doi întrerup. În ambele grupuri, un pacient suferă abateri de la protocolul prevăzut din cauza infecției. În săptămâna 6, rata durerii neuropatice este semnificativ mai mică în grupul de masaj cu aromoterapie (N = 4, % = 18,2; N = 11, % = 45,8; p = 0,046). Acest studiu nu poate fi inclus în meta-analiză deoarece evaluarea este prea eterogenă (DN4, cu variabile dihotomice) pentru a fi comparată cu celelalte studii.
Studiul lui Louis și Kowalski din 2022 [ 77] evaluează eficacitatea aromoterapiei cu ulei esențial de lavandă umidificat asupra durerii, anxietății, depresiei și sentimentului de bunăstare la 17 pacienți cu cancer. Designul acestui studiu este un design cvasi-experimental, cu măsuri repetate, cu un singur grup; prin urmare, riscul de părtinire este evaluat conform instrumentului ROBINS-I. Tabelul cu caracteristicile de referință cu analiză statistică nu este raportat, ridicând astfel îngrijorări în ceea ce privește riscul de părtinire din cauza factorilor de confuzie și selecția participanților la studiu, grupați în domeniul 1 pentru a permite compararea cu celelalte studii randomizate. Deoarece scara selectată este VAS, nu există niciun risc de părtinire în măsurarea rezultatului. Nu sunt raportate marimea eșantionului sau menționarea abandonului sau lipsei datelor. Durerea scade de la 1,70–1,66 din grupurile de control la 1,25 din intervenție.
Studiul lui Maddocks-Jennings și al colegilor din 2009 [ 78] este un studiu de fezabilitate randomizat, controlat cu placebo, care investighează efectul gargara care conține două picături dintr-un amestec 1:1 de uleiuri esențiale de manuka și kanuka în apă asupra mucozitei induse de radiații a zonei orofaringiene în timpul tratamentului pentru cap și gât. cancere. După cum au raportat autorii, o limitare semnificativă a acestui studiu este dimensiunea mică a eșantionului (N = 19). Se alege un eșantion de conveniență de douăzeci și șase de pacienți, dar patru pacienți abandonează voluntar, doi din grupul activ întrerup, iar un pacient din grupul de control se abate de la intervenția intenționată. Datorită dimensiunii reduse a eșantionului, aceste probleme ridică o mare îngrijorare cu privire la riscul de părtinire, dar personalul clinic și oncologii radioterapici care efectuează evaluările sunt nevăzători față de brațul de tratament căruia îi aparțin pacienții. Durerea datorată mucozitei este măsurată în mod corespunzător prin scala VAS. În cadrul grupului activ, n = 2 pacienți prezintă scoruri de durere ≥3, n = 5 din grupul de control și n = 4 din grupul placebo. Acest studiu nu poate fi inclus în meta-analiză din cauza eterogenității raportării măsurilor de rezultat.
Studiul randomizat (randomizarea este stratificată în funcție de vârstă), controlat cu placebo, dublu-orb, condus de Ndao și colab. în 2012 [ 79] își propune să evalueze efectul inhalării uleiului esențial de bergamotă asupra durerii, anxietății și greaței la copiii și adolescenții supuși perfuziei cu celule stem. Acesta implică 37 din 40 de pacienți necesari pentru a furniza 80% putere, dar niciun pacient nu renunță la studiu în timpul studiului după alocare. Caracteristicile inițiale nu arată diferențe semnificative statistic, cu excepția durerii inițiale, așa cum au raportat autorii. Alegerea placebo, constând în șampon parfumat pe bază de uleiuri neesențiale, nu este direct comparabilă cu intervenția. O altă limitare a acestui studiu, afirmată de autori, este dimensiunea mică a eșantionului format din pacienți cu diferite diagnostice și istorice de tratament. Schema de tratament este foarte diferită de celelalte studii, prevenind astfel metaanaliza și este după cum urmează: cu o săptămână înainte de transplant (T1), după administrarea de medicamente intravenoase și înainte de perfuzia cu celule stem sau măduvă osoasă (T2), după terminarea perfuziei (T3) și la o oră după terminarea perfuziei (T4). Nu este înregistrat niciun efect semnificativ asupra durerii.
Studiul lui Shamas et al., 2021 [ 80] este un studiu prospectiv, simplu-orb, randomizat, controlat, cu scopul de a evalua efectul uleiului de lavandă (plasat pe încheieturile mâinilor stângi și drepte pentru a freca cu mâinile și asupra tâmplelor intraoperator) asupra anxietății, depresiei, durerii și somnului. la femeile supuse reconstrucției microvasculare a sânilor din cauza cancerului de sân. Uleiul de nucă de cocos fracționat este selectat ca control fiind incolor, inodor și inert în natură. Randomizarea este o randomizare pe bloc și doar coordonatorul studiului cunoaște alocarea. Din cei 29 de pacienți alocați pe grup, 27 în intervenție și 22 în grupul de control completează studiul. O limitare a studiului raportată de autori este că dimensiunea eșantionului nu este calculată a priori, ci aleasă în mod arbitrar. Nu apare niciun efect semnificativ asupra durerii, în absența oricăror evenimente adverse sau complicații semnificative. Deoarece rezultatele durerii sunt raportate ca un model mixt de contabilizare pentru ArmxTime, aceste date nu sunt potrivite pentru meta-analiză.
În studiul randomizat, controlat, realizat de Soden și colegii săi în 2004 [ 81], patruzeci și doi de pacienți sunt alocați aleatoriu masajului cu aromoterapie cu ulei esențial de lavandă și un ulei purtător inert (grupul de aromoterapie) în comparație cu masajul cu un ulei purtător inert singur (grup de masaj) sau fără intervenție (grupul de control). Intensitatea durerii este evaluată în mod corespunzător prin VAS și Verbal Rating Scale (VRS). De fapt, rezultatul principal al prezentului studiu este efectul asupra scorurilor de intensitate a durerii, în timp ce rezultatele secundare includ calitatea somnului, anxietatea, depresia și calitatea generală a vieții. În plus, calculul puterii eșantionului este efectuat pentru a putea detecta o îmbunătățire a durerii de la valoarea inițială de 2,3 puncte VAS în comparație cu grupul de control (cu o putere = 80%), recrutând 15 pacienți pentru fiecare braț al studiului. Pacienții sunt alocați aleatoriu într-unul din trei grupuri, alocarea tratamentului este ascunsă în timpul evaluărilor inițiale, iar pacienții și cercetătorii care înregistrează și analizează datele sunt orbiți față de intervenții, prevenind astfel riscul de părtinire pentru randomizare și ascunderea alocării. Cu toate acestea, unele preocupări sunt ridicate de dovezile raportate de autori că, deși nu este observată nicio diferență semnificativă în evaluările de bază, și pentru intensitatea durerii, există semnificativ mai multe femei în grupul de control decât oricare dintre celelalte grupuri (p = 0,02) iar cazurile de depresie au un prag de cut-off în masaj decât în grupul de masaj cu aromoterapie ( p = 0,03). Șase pacienți nu finalizează studiul. Nu există nicio diferență semnificativă statistic în intensitatea durerii de la evaluarea finală la evaluarea inițială și, deoarece abaterea standard nu este raportată, acest studiu nu este eligibil pentru meta-analiză.
Triana și colaboratorii în 2022 [ 82] efectuează un studiu cvasi-experimental cu o tehnică de eșantionare consecutivă cu scopul de a înțelege eficacitatea aromoterapiei la copiii și adolescenții de vârstă școlară (7–17 ani) cu diagnostic de cancer, care suferă de durere cronică (mai mult de 1 lună) și primirea moderată de analgezice/opioide. Un eșantion de 20 de pacienți sunt repartizați aleatoriu în două grupe: Intervenție (N = 10), constând în aromaterapie cu cel mai favorit parfum (cel mai adesea aloe vera) și Control (N = 10) cu îngrijire standard. Dimensiunea mică a eșantionului și lipsa de menționare a oricăror abandonuri și rezultate lipsă ridică unele preocupări cu privire la riscul de părtinire. Asistentele administrează o intervenție de aromoterapie prin picurarea a patru picături din uleiul esențial selectat pe tifon curat, lipindu-l de pieptul pacientului la 20 cm și evaluând durerea folosind VAS la 10 și 30 min.p = 0,001) comparativ cu îngrijirea standard.
Principalele caracteristici ale studiilor incluse în analiză sunt raportate întabelul 1.
tabelul 1
Principalele caracteristici ale studiilor incluse în analiză conform recomandărilor Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) 2020.
Studiu
Populația
Intervenţie
Comparator
Măsuri finale
Design de studiu
Analiza puterii
Aprobarea și cererea de consimțământ informat de către Comitetul de etică/ Comitetul de examinare instituțională
Pacienți cu vârsta de 18 ani sau peste, care au un nou diagnostic de leucemie acută mielogenă și care inițiază patru săptămâni de chimioterapie intensivă de inducție
Lavandă, mentă sau mușețel
Placebo
Scala de evaluare a simptomelor Edmonton – revizuită cu scala vizuală analogică (VAS)
Studiu randomizat, încrucișat, cu spălare, N = 50; Prima aromaterapie, N = 25; Primul placebo, N = 28
Pacienții diagnosticați cu cancer supuși inserției de cateter portuar cu vârsta de 18 ani sau mai mult
Amestec aromat preparat prin diluarea uleiului de portocale, mușețel și lavandă în proporție de 1:1:1 în 70 ml apă distilată pentru inhalare
Practici de rutină
VAS
Studiu nerandomizat, controlat, N = 60; Intervenție, N = 30; Control, N = 30
+
+
Aromaterapia (5 ± 1,2) reduce durerea în timpul și după procedură într-o manieră semnificativă statistic în comparație cu controlul (adică, fără tratament; 7,4 ± 1,4)
Pacienți cu vârsta peste 18 ani afectați de mucozită indusă de radiații a zonei orofaringiene în timpul tratamentului pentru cancerele capului și gâtului
Gargară care conține 2 picături dintr-un amestec 1:1 de uleiuri esențiale de manuka și kanuka în apă + îngrijire orală obișnuită conform prescripției
Sticlă cu apă sterilă pentru gargară + îngrijire orală obișnuită conform prescripției. În loc de placebo, primește îngrijirea obișnuită
VAS
Studiu de fezabilitate randomizat, controlat cu placebo, N = 19; Grupa activă, N = 9; Grupa placebo, N = 6; Grupul de control, N = 8
–
+
În cadrul grupului activ, n = 2 pacienți prezintă scoruri de durere ≥ 3, n = 5 din grupul de control și n = 4 din grupul placebo
Copii și adolescenți de vârstă școlară (7-17 ani), cu diagnostic de cancer, care suferă de durere cronică (mai mult de 1 lună) și care primesc analgezice/opioide moderate
Inhalarea cu aromoterapie a unui parfum care îi place participantului (preferat, aloe vera)
Îngrijire standard (calmante și o tehnică de relaxare)
VAS
Cvasi-experimental cu eșantionare consecutivă, N = 20; Grup de intervenție, N = 10; Grupul de control, N = 10
–
+
Aromaterapia inhalată reduce semnificativ durerea cronică ( p = 0,001) în comparație cu îngrijirea standard. La minutul 30, Intervenție = 0,4 ± 0,97 și Control = 6,6 ± 1,43
Meta-analiză, care include șase din douăsprezece studii incluse în sinteză (parcela de pădure raportată înFigura 3), demonstrează eficacitatea semnificativă statistic a utilizării uleiurilor esențiale în reducerea intensității durerii asociate cancerului, evaluată prin scale de durere unidimensionale (p = 0,002 ) . În special, studiile contribuie aproape în egală măsură la rezultate, în afară de studiul realizat de Triana și colaboratori [ 82 ], care are o pondere mai mică datorită dimensiunii reduse a eșantionului și deoarece prezintă o abatere standard mai mare. Mai mult, este posibil să se evidențieze o eterogenitate ridicată în rândul studiilor eligibile pentru metaanaliză (I 2 = 96%). Graficul pâlnie sugerează riscul de părtinire a publicării (Figura 4).
Graficul forestier al meta-analizei pentru care șase din 12 studii extrase sunt eligibile. Uleiurile esențiale reprezintă intervenția, singure sau în combinație cu alte tratamente, iar placebo reprezintă controlul sau cel mai inert comparator inclus în studiile cu mai mult de un comparator [ 71 , 72 , 73 , 74 , 76 , 82 ].
Peste 80% dintre pacienții afectați de cancer suferă de durere, ceea ce reprezintă, prin urmare, una dintre cele mai înfricoșătoare consecințe ale cancerului [ 83 ]. Prezenta revizuire sistematică și meta-analiză evidențiază deficitul de studii clinice în domeniul aromoterapiei și al utilizării uleiurilor esențiale pentru a gestiona durerea asociată cu cancerul. Toate studiile eligibile pentru includerea în această analiză includ pacienți cu vârsta peste 18 ani, în afară de studiile efectuate de Ndao și colaboratori [ 79 ] și Triana și colegii [ 82]] concentrându-se pe o populație pediatrică. Cu toate acestea, multe tipuri de cancer permit o supraviețuire mai lungă decât în trecutul apropiat; astfel, trebuie luată în considerare posibilitatea unor comorbidități legate de vârstă. În special, în afară de studiul realizat de Triana și colegii în care durerea este dedusă și evaluată de asistentă [ 82], capacitatea de auto-raportare a durerii și de a răspunde la evaluarea intensității durerii prin intermediul VAS/NRS/VRS este unul dintre cele mai frecvente criterii de includere a studiilor recuperate prin această căutare sistematică. Acest aspect este demn de remarcat, deoarece indică necesitatea unei evaluări mai adecvate a durerii în timpul cancerului. În special, durerea legată de cancer este caracterizată printr-o natură multidimensională constând din diferite fiziopatologie și etiologie și care include componente senzoriale, afective, cognitive și comportamentale importante, iar cercetările încă nu au identificarea acestor trăsături fundamentale [84] .]. Prin urmare, evaluarea durerii prin scale unidimensionale, deși adecvată pentru afecțiunile acute și pentru evaluarea domeniului de intensitate unic, ar trebui să fie flancată și înlocuită cu utilizarea scalelor multidimensionale ca Brief Pain Inventory (BPI) [85 ] . De fapt, BPI permite măsurarea atât a dimensiunii senzoriale a intensității durerii, cât și a dimensiunii reactive a interferenței durerii în viața pacientului [ 85] .]. Acest lucru este din ce în ce mai important pentru evaluarea eficacității și siguranței uleiurilor esențiale asupra durerii în oncologia integrativă. Studiile găsite prin căutarea prezentă a bazelor de date și inspecția referințelor din literatură demonstrează un efect principal al uleiurilor esențiale asupra bunăstării generale și a somnului. Cu toate acestea, aceste simptome și beneficiile acestora ar putea fi legate de procesarea durerii [ 86 ], deși nu sunt întotdeauna detectate de un instrument de măsurare adecvat, în principal la pacienții în vârstă și în cazul depresiei [ 87 ]. Acest lucru este susținut de meta-analiză care demonstrează eficacitatea uleiurilor esențiale utilizate ca aromoterapie în reducerea intensității durerii asociate cancerului, evaluată prin scale de durere unidimensionale (p .= 0,002). Cu toate acestea, în acord cu lipsa de utilizare a dispozitivelor omogene și adecvate, doar șase din douăsprezece studii incluse în sinteză sunt eligibile pentru metaanaliză și acest lucru este dovedit de eterogenitatea ridicată dintre studii (I 2 = 96% ) și prin părtinirea publicării care apar în domeniul uleiurilor esențiale, precum și al suplimentelor orale și al nutraceuticelor [ 88 , 89 ]. În special, studiile incluse subliniază modele de studiu foarte diferite și unele preocupări în ceea ce privește riscul de părtinire, care decurg în principal din evaluarea inadecvată de referință și datele privind rezultatele și, de asemenea, din cauza dimensiunii reduse a eșantionului. De exemplu, studiul realizat de Triana și colaboratori [ 82] prezintă o pondere mai mică în meta-analiză din cauza dimensiunii sale mici ale eșantionului. Problema evaluării durerii este și mai importantă pentru populațiile în vârstă cu declin cognitiv, având nevoie de instrumente de observație adecvate [ 90 , 91 , 92 ], precum și de metode valide și de încredere, cu proprietăți psihomometrice și clinometrice bune în acest cadru și care merită luate în considerare pentru suplimentare. sursele durerii și tratamentul lor [ 93 , 94 , 95 , 96 , 97]. În plus, diferite tipuri de cancer pot influența durerea, îngreunând astfel comparația. Unul dintre cele mai utilizate uleiuri esențiale este uleiul esențial de lavandă, dar întrucât implică în principal sistemul colinergic, acesta nu este înzestrat cu o rațiune preclinică puternică pentru activitatea analgezică [98 , 99 ] ; asta se întâmplă în schimb pentru BEO. Lipsa de eficacitate a acestuia din urmă în studiul realizat de Ndao și colaboratori [ 79] ar putea fi, cel puțin parțial, explicată prin dimensiunea mică a eșantionului format din pacienți cu diagnostice și istorice diferite de tratament. Mai mult, lipsa reducerii intensității durerii folosind uleiuri esențiale în unele studii se poate datora unui început târziu al tratamentului care poate acționa doar ca paliativ atunci când durerea cronică este stabilită, în special pentru CIPN, în timp ce aceasta ar trebui prevenită cu o terapie mai timpurie [ 100 ] . , 101 ]. Prin urmare, pentru a oferi o bună certitudine cu privire la corpul de dovezi pentru gestionarea durerii asociate cancerului folosind uleiuri esențiale, este necesar să se stabilească, pentru produsele naturale, o cale pas cu pas de la preclinic la clinic, care să ofere o justificare pentru eficient și eficient. utilizare sigură.
Această cercetare este coordonată de DS și a primit sprijin financiar parțial de la: (1) Etapa 2 Finanțare RIABEO (Decretul executiv nr.6790 din 22 iunie 2022) Progetto Ingegno POR Calabria FESR 2014/2020—Azione 1 1 5—Sostegno all’Avanzamento tecnologico delle Imprese Attraverso il Finanziamento di Linee Pilota e Azioni di Validazione Precoce di Prodotti e di Dimostrazione su Larga Scala (DDG N. 12814 DEL 17 octombrie 2019); (2) de către Ministerul italian al Sănătății: NET-2016–02361805 (WP 5).
Conceptualizare, DS, MTC, GB, LAM, PT și SM; Metodologia și curatarea datelor, KH, TH, FG și DS Toți autorii au citit și au fost de acord cu versiunea publicată a manuscrisului.
Disclaimer/Nota editorului: Declarațiile, opiniile și datele conținute în toate publicațiile sunt exclusiv ale autorilor și colaboratorilor individuali și nu ale MDPI și/sau ale editorilor. MDPI și/sau editorii își declină responsabilitatea pentru orice vătămare a persoanelor sau a proprietății care rezultă din orice idei, metode, instrucțiuni sau produse la care se face referire în conținut.
Portenoy RK, Thaler HT, Kornblith AB, McCarthy Lepore J., Friedlander-Klar H., Coyle N., Smart-Curley T., Kemeny N., Norton L., Hoskins W., et al. Prevalența simptomelor, caracteristicile și suferința într-o populație cu cancer. Cal. Life Res. 1994; 3 :183–189. doi: 10.1007/BF00435383. [ PubMed ] [ CrossRef ] [ Google Scholar ]2.
Managementul grupului de simptome de cancer Kwekkeboom KL. Semin. Oncol. Asistente medicale. 2016; 32 :373–382. doi: 10.1016/j.soncn.2016.08.004. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]3.
Societatea Americană de Cancer. Fapte și cifre privind tratamentul cancerului și supraviețuirea 2022–2024. Societatea Americană de Cancer; Atlanta, GA, SUA: 2022. [ Google Scholar ]4.
Mantovani A., Allavena P., Sica A., Balkwill F. Cancer-related inflammation. Natură. 2008; 454 :436–444. doi: 10.1038/nature07205. [ PubMed ] [ CrossRef ] [ Google Scholar ]5.
Van den Beuken-van Everdingen MH, de Rijke JM, Kessels AG, Schouten HC, van Kleef M., Patijn J. Prevalența durerii la pacienții cu cancer: o revizuire sistematică a ultimilor 40 de ani. Ann. Oncol. 2007; 18 :1437–1449. doi: 10.1093/annonc/mdm056. [ PubMed ] [ CrossRef ] [ Google Scholar ]6.
Van den Beuken-van Everdingen MH, Hochstenbach LM, Joosten EA, Tjan-Heijnen VC, Janssen DJ Actualizare privind prevalența durerii la pacienții cu cancer: revizuire sistematică și meta-analiză. J. Gestionarea simptomelor durerii. 2016; 51 :1070–1090.e1079. doi: 10.1016/j.jpainsymman.2015.12.340. [ PubMed ] [ CrossRef ] [ Google Scholar ]7.
Breivik H., Cherny N., Collett B., de Conno F., Filbet M., Foubert AJ, Cohen R., Dow L. Cancer-related pain: A pan-european survey of prevalence, treatment, and patient atitudini. Ann. Oncol. 2009; 20 :1420–1433. doi: 10.1093/annonc/mdp001. [ PubMed ] [ CrossRef ] [ Google Scholar ]8.
Thomas EM, Weiss SM Intervenții nonfarmacologice cu durere cronică de cancer la adulți. Controlul cancerului. 2000; 7 :157–164. doi: 10.1177/107327480000700206. [ PubMed ] [ CrossRef ] [ Google Scholar ]9.
Haenen V., Evenepoel M., De Baerdemaecker T., Meeus M., Devoogdt N., Morlion B., Dams L., Van Dijck S., Van der Gucht E., De Vrieze T., et al. Prevalența și caracteristicile durerii la supraviețuitorii cancerelor solide: o revizuire sistematică și meta-analiză. A sustine. Îngrijire Cancer. 2022; 31:85 . doi: 10.1007/s00520-022-07491-8. [ PubMed ] [ CrossRef ] [ Google Scholar ]10.
Tan HS, Plichta JK, Kong A., Tan CW, Hwang S., Sultana R., Wright MC, Sia ATH, Sng BL, Habib AS Factori de risc pentru durerea persistentă după intervenția chirurgicală a cancerului de sân: un studiu de cohortă prospectiv multicentric. Anestezie. 2023 doi: 10.1111/anae.15958. [ PubMed ] [ CrossRef ] [ Google Scholar ]11.
Schappert SM, Burt CW Vizite de îngrijire ambulatorie la cabinetele medicului, secțiile de ambulatoriu ale spitalelor și departamentele de urgență: Statele Unite, 2001–2002. Statul Sănătăţii Vitale. 2006; 159 :1–66. [ PubMed ] [ Google Scholar ]12.
Russo MM, Sundaramurthi T. O privire de ansamblu asupra durerii canceroase: Epidemiologie și fiziopatologie. Semin. Oncol. Asistente medicale. 2019; 35 :223–228. doi: 10.1016/j.soncn.2019.04.002. [ PubMed ] [ CrossRef ] [ Google Scholar ]13.
Treede RD, Jensen TS, Campbell JN, Cruccu G., Dostrovsky JO, Griffin JW, Hansson P., Hughes R., Nurmikko T., Serra J. Neuropathic pain: Redefinition and a grading system for clinical and research purposes. Neurologie. 2008; 70 :1630–1635. doi: 10.1212/01.wnl.0000282763.29778.59. [ PubMed ] [ CrossRef ] [ Google Scholar ]14.
Bajwa SJ, Haldar R. Managementul durerii în urma intervențiilor chirurgicale ale coloanei vertebrale: o evaluare a opțiunilor disponibile. J. Craniovertebral Junction Spine. 2015; 6 :105–110. doi: 10.4103/0974-8237.161589. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]15.
Masri R., Keller A. Dureri cronice în urma leziunii măduvei spinării. Adv. Exp. Med. Biol. 2012; 760 :74–88. doi: 10.1007/978-1-4614-4090-1_5. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]16.
Scuteri D., Mantovani E., Tamburin S., Sandrini G., Corasaniti MT, Bagetta G., Tonin P. Opioids in Post-stroke Pain: A Systematic Review and Meta-Analysis. Față. Pharmacol. 2020; 11 :587050. doi: 10.3389/fphar.2020.587050. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]17.
Portenoy RK, Ahmed E. Cancer Pain Syndromes. Hematol. Oncol. Clin. N. Am. 2018; 32 :371–386. doi: 10.1016/j.hoc.2018.01.002. [ PubMed ] [ CrossRef ] [ Google Scholar ]18.
Farquhar-Smith P. Durerea neuropatică indusă de chimioterapie. Curr. Opinează. A sustine. Palliat. Îngrijire. 2011; 5 :1–7. doi: 10.1097/SPC.0b013e328342f9cc. [ PubMed ] [ CrossRef ] [ Google Scholar ]19.
Quintão NLM, Santin JR, Stoeberl LC, Corrêa TP, Melato J., Costa R. Tratamentul farmacologic al durerii neuropatice induse de chimioterapie: Agoniştii PPARγ ca instrument promiţător. Față. Neurosci. 2019; 13 :907. doi: 10.3389/fnins.2019.00907. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]20.
Bayer A., Tadd W. Excluderea nejustificată a persoanelor în vârstă de la studiile prezentate spre aprobare comitetului de etică a cercetării: Studiu descriptiv. BMJ. 2000; 321 :992–993. doi: 10.1136/bmj.321.7267.992. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]21.
Scuteri D., Adornetto A., Rombolà L., Naturale MD, De Francesco AE, Esposito S., Zito M., Morrone LA, Bagetta G., Tonin P., et al. Modelul de utilizare a triptanilor: un studiu retrospectiv de prescripție în Calabria, Italia. Regenerare neuronală. Res. 2020; 15 :1340–1343. doi: 10.4103/1673-5374.272630. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]22.
Scuteri D., Corasaniti MT, Tonin P., Bagetta G. Eptinezumab pentru tratamentul migrenei. Droguri azi. 2019; 55 :695–703. doi: 10.1358/dot.2019.55.11.3069864. [ PubMed ] [ CrossRef ] [ Google Scholar ]23.
Scuteri D., Corasaniti MT, Tonin P., Nicotera P., Bagetta G. Rolul polimorfismelor căii CGRP în migrenă: o revizuire sistematică și impact asupra terapiei migrenei CGRP mAbs. J Cefalee Durere. 2021; 22:87 . doi: 10.1186/s10194-021-01295-7. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]24.
Nørgaard A., Jensen-Dahm C., Gasse C., Hansen ES, Waldemar G. Psychotropic Polypharmacy in Patients with Dementa: Prevalence and Predictors. J. Alzheimer Dis. 2017; 56 :707–716. doi: 10.3233/JAD-160828. [ PubMed ] [ CrossRef ] [ Google Scholar ]25.
Scuteri D., Berliocchi L., Rombolà L., Morrone LA, Tonin P., Bagetta G., Corasaniti MT Efectele îmbătrânirii asupra comportamentului durerii induse de formalină și activității analgezice a gabapentinei la șoareci C57BL/6. Față. Pharmacol. 2020; 11 :663. doi: 10.3389/fphar.2020.00663. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]26.
McLachlan AJ, Hilmer SN, Le Couteur DG Variabilitatea ca răspuns la medicamente la persoanele în vârstă: factori fenotipici și genotipici. Clin. Pharmacol. Acolo. 2009; 85 :431–433. doi: 10.1038/clpt.2009.1. [ PubMed ] [ CrossRef ] [ Google Scholar ]27.
Mao JJ, Ismaila N., Bao T., Barton D., Ben-Arye E., Garland EL, Greenlee H., Leblanc T., Lee RT, Lopez AM și colab. Medicină integrativă pentru managementul durerii în oncologie: Societatea pentru Oncologie Integrativă – Ghidul ASCO. J. Clin. Oncol. 2022; 40 :3998–4024. doi: 10.1200/JCO.22.01357. [ PubMed ] [ CrossRef ] [ Google Scholar ]28.
Bardia A., Barton DL, Prokop LJ, Bauer BA, Moynihan TJ Eficacitatea terapiilor de medicină complementară și alternativă în ameliorarea durerii canceroase: o revizuire sistematică. J. Clin. Oncol. 2006; 24 :5457–5464. doi: 10.1200/JCO.2006.08.3725. [ PubMed ] [ CrossRef ] [ Google Scholar ]29.
Rombolà L., Scuteri D., Watanabe C., Sakurada S., Hamamura K., Sakurada T., Tonin P., Corasaniti MT, Bagetta G., Morrone LA Role of 5-HT1A Receptor in the Anxiolytic-Relaxant Efectele uleiului esențial de bergamotă la rozătoare. Int. J. Mol. Sci. 2020; 21 :2597. doi: 10.3390/ijms21072597. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]30.
Scuteri D., Cassano R., Trombino S., Russo R., Mizoguchi H., Watanabe C., Hamamura K., Katsuyama S., Komatsu T., Morrone LA, et al. Dezvoltarea și traducerea NanoBEO, un sistem de livrare bazat pe nanotehnologie a uleiului esențial de bergamotă lipsit de furocumarine, în controlul agitației în demența severă. Farmaceutică. 2021; 13 :379. doi: 10.3390/pharmaceutics13030379. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]31.
Scuteri D., Hamamura K., Sakurada T., Watanabe C., Sakurada S., Morrone LA, Rombolà L., Tonin P., Bagetta G., Corasaniti MT Efficacy of Essential Oils in Pain: A Systematic Review and Meta-analiză a dovezilor preclinice. Față. Pharmacol. 2021; 12 :640128. doi: 10.3389/fphar.2021.640128. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]32.
Scuteri D., Sandrini G., Tamburin S., Corasaniti MT, Nicotera P., Tonin P., Bagetta G. Bergamot rehabilitation AgaINst agitation in dementa (BRAINAID): Study protocol for a randomized, double-orb, placebo- studiu controlat pentru a evalua eficacitatea bergamotei fără furocumarină încărcată într-un sistem de livrare bazat pe nanotehnologie a uleiului esențial în tratamentul agitației la vârstnicii afectați de demență severă. Phytother. Res. 2021; 35 :5333–5338. doi: 10.1002/ptr.7223. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]33.
Scuteri D., Rombolà L., Crudo M., Watanabe C., Mizoguchi H., Sakurada S., Hamamura K., Sakurada T., Morrone LA, Tonin P., et al. Valoarea de translație a administrării transdermice a uleiului esențial de bergamotă și a fracțiilor sale. Farmaceutică. 2022; 14 :1006. doi: 10.3390/pharmaceutics14051006. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]34.
Scuteri D., Rombolà L., Crudo M., Watanabe C., Mizoguchi H., Sakurada S., Hamamura K., Sakurada T., Tonin P., Corasaniti MT, et al. Caracterizarea preclinică a efectului antinociceptiv al uleiului esențial de bergamotă și a fracțiilor sale pentru traducerea rațională în terapia complementară. Farmaceutică. 2022; 14 :312. doi: 10.3390/pharmaceutics14020312. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]35.
Hamamura K., Katsuyama S., Komatsu T., Scuteri D., Bagetta G., Aritake K., Sakurada T. Behavioral Effects of Continuously Administered Bergamot Essential Oil on Mice With Partial Sciatic Nerve Ligation. Față. Pharmacol. 2020; 11 :1310. doi: 10.3389/fphar.2020.01310. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]36.
Scuteri D., Crudo M., Rombolà L., Watanabe C., Mizoguchi H., Sakurada S., Sakurada T., Greco R., Corasaniti MT, Morrone LA, et al. Efectul antinociceptiv al inhalării uleiului esențial de bergamotă la șoareci. Fitoterapia. 2018; 129 :20–24. doi: 10.1016/j.fitote.2018.06.007. [ PubMed ] [ CrossRef ] [ Google Scholar ]37.
Scuteri D., Rombolà L., Hayashi T., Watanabe C., Sakurada S., Hamamura K., Sakurada T., Tonin P., Bagetta G., Morrone LA, et al. Caracteristicile analgezice ale NanoBEO eliberat de un distribuitor fără aer pentru controlul agitației în demența severă. Molecule. 2022; 27 :4987. doi: 10.3390/molecules27154987. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]38.
Page MJ, McKenzie JE, Bossuyt PM, Boutron I., Hoffmann TC, Mulrow CD, Shamseer L., Tetzlaff JM, Akl EA, Brennan SE, et al. Declarația PRISMA 2020: un ghid actualizat pentru raportarea evaluărilor sistematice. BMJ Clin. Res. Ed. 2021; 372 :n71. doi: 10.1136/bmj.n71. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]39.
Liberati A., Altman DG, Tetzlaff J., Mulrow C., Gøtzsche PC, Ioannidis JP, Clarke M., Devereaux PJ, Kleijnen J., Moher D. Declarația PRISMA pentru raportarea revizuirilor sistematice și meta-analizelor studiilor care evaluează intervenţiile de îngrijire a sănătăţii: Explicaţie şi elaborare. PLoS Med. 2009; 6 :e1000100. doi: 10.1371/journal.pmed.1000100. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]40.
Moher D., Liberati A., Tetzlaff J., Altman DG, Group P. Elemente de raportare preferate pentru revizuiri sistematice și meta-analize: Declarația PRISMA. PLoS Med. 2009; 6 :e1000097. doi: 10.1371/journal.pmed.1000097. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]41.
Jones J., Hunter D. Cercetare calitativă: Metode de consens pentru cercetarea serviciilor medicale și de sănătate. BMJ. 1995; 311 :376–380. doi: 10.1136/bmj.311.7001.376. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]42.
Lefebvre C., Glanville J., Briscoe S., Littlewood A., Marshall C., Metzendorf M.-I., Noel-Storr A., Rader T., Shokraneh F., Thomas J., et al. Manual Cochrane pentru revizuiri sistematice ale intervențiilor. Wiley; Hoboken, NJ, SUA: 2019. Căutarea și selectarea studiilor. [ Google Scholar ]43.
McGowan J., Sampson M., Salzwedel DM, Cogo E., Foerster V., Lefebvre C. PRESS Peer Review of Electronic Search Strategies: 2015 Guideline Statement. J. Clin. Epidemiol. 2016; 75 :40–46. doi: 10.1016/j.jclinepi.2016.01.021. [ PubMed ] [ CrossRef ] [ Google Scholar ]44.
Ryan R. Cochrane Consumers and Communication Review Group: Sinteza și analiza datelor. [(accesat la 13 martie 2019)]. Disponibil online: http://cccrg.cochrane.org45.
Hultcrantz M., Rind D., Akl EA, Treweek S., Mustafa RA, Iorio A., Alper BS, Meerpohl JJ, Murad MH, Ansari MT, et al. Grupul de lucru GRADE clarifică constructul de certitudine a dovezilor. J. Clin. Epidemiol. 2017; 87 :4–13. doi: 10.1016/j.jclinepi.2017.05.006. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]46.
Page MJ, Moher D., Bossuyt PM, Boutron I., Hoffmann TC, Mulrow CD, Shamseer L., Tetzlaff JM, Akl EA, Brennan SE, et al. Explicația și elaborarea PRISMA 2020: orientări actualizate și exemple pentru raportarea evaluărilor sistematice. BMJ Clin. Res. Ed. 2021; 372 :n160. doi: 10.1136/bmj.n160. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]47.
Sterne JAC, Savović J., Page MJ, Elbers RG, Blencowe NS, Boutron I., Cates CJ, Cheng H.-Y., Corbett MS, Eldridge SM și colab. RoB 2: Un instrument revizuit pentru evaluarea riscului de părtinire în studiile randomizate. BMJ. 2019; 366 :l4898. doi: 10.1136/bmj.l4898. [ PubMed ] [ CrossRef ] [ Google Scholar ]48.
Sterne JA, Hernán MA, Reeves BC, Savović J., Berkman ND, Viswanathan M., Henry D., Altman DG, Ansari MT, Boutron I., et al. ROBINS-I: Un instrument pentru evaluarea riscului de părtinire în studiile nerandomizate ale intervențiilor. BMJ. 2016; 355 :i4919. doi: 10.1136/bmj.i4919. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]49.
McGuinness LA, Higgins JPT Risk-of-bias VISualization (robvis): Un pachet R și aplicație web Shiny pentru vizualizarea evaluărilor riscului de părtinire. Res. Sintetizator. Metode. 2020; 12 :55–61. doi: 10.1002/jrsm.1411. [ PubMed ] [ CrossRef ] [ Google Scholar ]50.
DerSimonian R., Kacker R. Modelul cu efecte aleatoare pentru meta-analiză a studiilor clinice: o actualizare. Contemp. Clin. Încercări. 2007; 28 :105–114. doi: 10.1016/j.cct.2006.04.004. [ PubMed ] [ CrossRef ] [ Google Scholar ]51.
Egger M., Smith GD, Schneider M., Minder C. Bias in meta-analysis detected by a simple, graphical test. BMJ. 1997; 315 :629. doi: 10.1136/bmj.315.7109.629. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]53.
Sterne JA, Egger M. Funnel plots for detecting bias in meta-analisis: Guidelines on choice of axis. J. Clin. Epidemiol. 2001; 54 :1046–1055. doi: 10.1016/S0895-4356(01)00377-8. [ PubMed ] [ CrossRef ] [ Google Scholar ]54.
Duval S., Tweedie R. Trim and Fill: A Simple Funnel-Plot-Based Method of Testing and Adjusting for Publication Bias in Meta-Analysis. Biometrie. 2000; 56 :455–463. doi: 10.1111/j.0006-341X.2000.00455.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]55.
Hasheminasab FS, Hashemi SM, Dehghan A., Sharififar F., Setayesh M., Sasanpour P., Tasbandi M., Raeiszadeh M. Efectele unui compus din plante pe bază de Plantago ovata în prevenirea și tratamentul mucozitei orale la pacienții cu cancer de sân care primește chimioterapie: un studiu dublu-orb, randomizat, controlat încrucișat. J. Integr. Med. 2020; 18 :214–221. doi: 10.1016/j.joim.2020.02.008. [ PubMed ] [ CrossRef ] [ Google Scholar ]56.
Aghamohammadi A., Moslemi D., Akbari J., Ghasemi A., Azadbakht M., Asgharpour A., Hosseiimehr SJ Eficacitatea apei de gură cu extract de Zataria pentru gestionarea mucozitei orale induse de radiații la pacienți: un placebo randomizat. studiu controlat dublu-orb. Clin. Oral Investig. 2018; 22 :2263–2272. doi: 10.1007/s00784-017-2324-7. [ PubMed ] [ CrossRef ] [ Google Scholar ]57.
Arantes DAC, da Silva ACG, Freitas NMA, Lima EM, de Oliveira AC, Marreto RN, Mendonça EF, Valadares MC. mucozită orală indusă: studiu clinic triplu-orb, randomizat, controlat cu placebo. Gâtul capului. 2021; 43 :3922–3934. doi: 10.1002/hed.26892. [ PubMed ] [ CrossRef ] [ Google Scholar ]58.
Czakert J., Stritter W., Blakeslee SB, Grabowski JP, Sehouli J., Seifert G. „Like one part of a puzzle” — Aromoterapie individualizată pentru femeile cu cancere ginecologice în îngrijirea ulterioară: Rezultatele unui studiu mixt centrat pe calitate studiul metodelor. A sustine. Îngrijire Cancer. 2022; 31:80 . doi: 10.1007/s00520-022-07543-z. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]59.
Ho SSM, Kwong ANL, Wan KWS, Ho RML, Chow KM Experiențe de masaj cu aromoterapie în rândul pacienților cu cancer de sex feminin adult: un studiu calitativ. J. Clin. Asistente medicale. 2017; 26 :4519–4526. doi: 10.1111/jocn.13784. [ PubMed ] [ CrossRef ] [ Google Scholar ]60.
Oyston E., McGee M. Oncology Massage Research and Training Update. J. Aust. Tradit. Med. Soc. 2012; 18 :11–14. [ Google Scholar ]61.
Lee WJ, Park H. Efectele inhalării aromei asupra tensiunii arteriale, frecvenței cardiace, durerii și calității somnului în rândul pacienților cu cancer pulmonar care au suferit rezecție pulmonară. Asia Life Sci. 2018; 1 ((Suppl. 15)):901–912. [ Google Scholar ]62.
Cheong MJ, Ha WB, Cho HB, Choi UJ, Woo HJ, Han YH, Kang HW Îmbunătățirea calității vieții la o pacientă și îngrijitoare cu cancer de sân: Protocol pentru aplicarea modelului de servicii medicale integrative. Medicament. 2022; 101 :e32244. doi: 10.1097/MD.0000000000032244. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]63.
Xiao Y., Li L., Xie Y., Xu J., Liu Y. [Efectele aromoterapiei și intervenției muzicale asupra durerii și anxietății pentru pacienții cu cancer de sân în perioada perioperatorie] Zhong Nan Da Xue Xue Bao Yi Xue Ban J. Cent. Univ. Sud. Med. Sci. 2018; 43 :656–661. doi: 10.11817/j.issn.1672-7347.2018.06.013. [ PubMed ] [ CrossRef ] [ Google Scholar ]64.
Chang SY Efectele masajului cu aromă asupra mâinilor asupra durerii, stării de anxietate și depresiei la pacienții cu cancer terminal. J. coreeană Acad. Asistente medicale. 2008; 38 :493–502. doi: 10.4040/jkan.2008.38.4.493. [ PubMed ] [ CrossRef ] [ Google Scholar ]65.
Nekuzad N., Torab TA, Mojab F., Alavi-Majd H., Azadeh P., Ehtejab G. Efectul utilizării externe a uleiului de susan în prevenirea flebitei induse de chimioterapie. Iranul. J. Pharm. Res. 2012; 11 :1065–1071. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]66.
Yayla EM, Ozdemir L. Efectul aromaterapiei prin inhalare asupra durerii și anxietății procedurale după inserarea acului într-un cateter implantabil cu port venos central: un studiu pilot controlat cvasi-randomizat. Asistente de cancer. 2019; 42 :35–41. doi: 10.1097/NCC.0000000000000551. [ PubMed ] [ CrossRef ] [ Google Scholar ]67.
Corbin LW, Mellis BK, Beaty BL, Kutner JS Utilizarea terapiilor de medicină complementară și alternativă de către pacienții cu cancer avansat și durere într-un cadru hospice: un studiu descriptiv multicentric. J. Palliat. Med. 2009; 12 :7–8. doi: 10.1089/jpm.2008.0198. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]68.
Ovayolu Ö., Seviğ Ü., Ovayolu N., Sevinç A. Efectul aromaterapiei și masajului administrat în moduri diferite femeilor cu cancer de sân asupra simptomelor și calității vieții acestora. Int. J. Nurs. Practică. 2014; 20 :408–417. doi: 10.1111/ijn.12128. [ PubMed ] [ CrossRef ] [ Google Scholar ]69.
Elhadad MA, El-Negoumy E., Taalab MR, Ibrahim RS, Elsaka RO Efectul mușețelului topic în prevenirea mucozitei orale induse de chimioterapie: un studiu clinic randomizat. Dis. orală. 2022; 28 :164–172. doi: 10.1111/odi.13749. [ PubMed ] [ CrossRef ] [ Google Scholar ]70.
Haddaway NR, Page MJ, Pritchard CC, McGuinness LA PRISMA2020: Un pachet R și aplicație Shiny pentru producerea de diagrame de flux conforme cu PRISMA 2020, cu interactivitate pentru transparență digitală optimizată și sinteză deschisă. Campbell Syst. Rev. 2022; 18 :e1230. doi: 10.1002/cl2.1230. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]71.
Blackburn L., Achor S., Allen B., Bauchmire N., Dunnington D., Klisovic RB, Naber SJ, Roblee K., Samczak A., Tomlinson-Pinkham K., et al. Efectul aromoterapiei asupra insomniei și a altor simptome comune la pacienții cu leucemie acută. Oncol. Asistente medicale. Forum. 2017; 44 :E185–E193. doi: 10.1188/17.ONF.E185-E193. [ PubMed ] [ CrossRef ] [ Google Scholar ]72.
Blackburn L., Hill C., Lindsey AL, Sinnott LT, Thompson K., Quick A. Effect of Foot Reflexology and Aromatherapy on Anxiety and Pain during Brachytherapy for Cervical Cancer. Oncol. Asistente medicale. Forum. 2021; 48 :265–276. doi: 10.1188/21.ONF.265-276. [ PubMed ] [ CrossRef ] [ Google Scholar ]73.
Deng C., Xie Y., Liu Y., Li Y., Xiao Y. Aromaterapia Plus Muziterapie Îmbunătățește intensitatea durerii și scorurile de anxietate la pacienții cu cancer de sân în timpul perioadelor perioperatorii: Un studiu controlat randomizat: Aromaterapie plus terapie prin muzică pe Durere și anxietate. Clin. Cancer mamar. 2022; 22 :115–120. doi: 10.1016/j.clbc.2021.05.006. [ PubMed ] [ CrossRef ] [ Google Scholar ]74.
Ha YS, Kim JW, Lee JE, Yang JH, Lee MJ, Lee EY, You JY, Jung SP, Choi HJ, Park KH. sindrom: un studiu randomizat controlat încrucișat. EURO. J. Integr. Med. 2022; 54 :102167. doi: 10.1016/j.eujim.2022.102167. [ CrossRef ] [ Google Scholar ]75.
Ilter SM, Ovayolu O., Ovayolu N. Efectul aromaterapiei inhalatoare asupra durerii invazive, aderenței la procedură, semnelor vitale și saturației în timpul cateterismului portuar la pacienții cu oncologie. Holist. Asistente medicale. Practică. 2019; 33 :146–154. doi: 10.1097/HNP.0000000000000322. [ PubMed ] [ CrossRef ] [ Google Scholar ]76.
Izgu N., Ozdemir L., Efectul bazal FB al masajului aromaterapiei asupra durerii neuropatice periferice induse de chimioterapie și oboselii la pacienții care primesc oxaliplatin Un studiu pilot controlat cvasi-randomizat deschis. Asistente de cancer. 2019; 42 :139–147. doi: 10.1097/NCC.0000000000000577. [ PubMed ] [ CrossRef ] [ Google Scholar ]77.
Louis M., Kowalski SD Utilizarea aromoterapiei cu pacienții din spitale pentru a reduce durerea, anxietatea și depresia și pentru a promova un sentiment crescut de bunăstare. A.m. J. Hosp. Palliat. Med. 2022; 19 :381–386. doi: 10.1177/104990910201900607. [ PubMed ] [ CrossRef ] [ Google Scholar ]78.
Maddocks-Jennings W., Wilkinson JM, Cavanagh HM, Shillington D. Evaluarea efectelor uleiurilor esențiale Leptospermum scoparium (manuka) și Kunzea ericoides (kanuka) asupra mucozitei induse de radioterapie: un studiu de fezabilitate randomizat, controlat cu placebo. EURO. J. Oncol. Asistente medicale. 2009; 13 :87–93. doi: 10.1016/j.ejon.2009.01.002. [ PubMed ] [ CrossRef ] [ Google Scholar ]79.
Ndao DH, Ladas EJ, Cheng B., Sands SA, Snyder KT, Garvin JH, Jr., Kelly KM Aromoterapie prin inhalare la copii și adolescenți supuși perfuziei cu celule stem: Rezultatele unui studiu dublu-orb controlat cu placebo. Psiho-Oncol. 2012; 21 :247–254. doi: 10.1002/pon.1898. [ PubMed ] [ CrossRef ] [ Google Scholar ]80.
Shammas RL, Marks CE, Broadwater G., Le E., Glener AD, Sergesketter AR, Cason RW, Rezak KM, Phillips BT, Hollenbeck ST Efectul uleiului de lavandă asupra durerii perioperatorii, anxietății, depresiei și somnului după microvasculare Reconstrucția sânilor: un studiu prospectiv, simplu-orb, randomizat, controlat. J. Reconstr. Microchirurgie. 2021; 37 :530–540. doi: 10.1055/s-0041-1724465. [ PubMed ] [ CrossRef ] [ Google Scholar ]81.
Soden K., Vincent K., Craske S., Lucas C., Asley S. Un studiu randomizat controlat al masajului cu aromoterapie într-un cadru hospice. Palliat. Med. 2004; 18 :87–92. doi: 10.1191/0269216304pm874oa. [ PubMed ] [ CrossRef ] [ Google Scholar ]82.
Triana KY, Allenidekania A., Hayati H. Efectul inhalării aromoterapiei asupra reducerii durerii cronice pentru copiii cu cancer: un studiu pilot. Trends Sci. 2022; 19 :2669. doi: 10.48048/tis.2022.2669. [ CrossRef ] [ Google Scholar ]83.
Bruera E., Kim HN Cancer Pain. JAMA. 2003; 290 :2476–2479. doi: 10.1001/jama.290.18.2476. [ PubMed ] [ CrossRef ] [ Google Scholar ]84.
Ahles TA, Blanchard EB, Ruckdeschel JC Natura multidimensională a durerii asociate cancerului. Durere. 1983; 17 :277–288. doi: 10.1016/0304-3959(83)90100-8. [ PubMed ] [ CrossRef ] [ Google Scholar ]85.
Cleeland CS, Ryan KM Evaluarea durerii: Utilizarea globală a Inventarului scurt al durerii. Ann. Acad. Med. Singap. 1994; 23 :129–138. [ PubMed ] [ Google Scholar ]86.
Scuteri D., Watanabe C., Sakurada S., Hamamura K., Sakurada T., Tonin P., Bagetta G., Corasaniti MT Pharmacotechnological Advances for Clinical Translation of Essential Oils for the Treatment of Pain and Agitation in Severe Dementa . Procese. 2022; 10 :1340. doi: 10.3390/pr10071340. [ CrossRef ] [ Google Scholar ]87.
Scuteri D., Corasaniti MT, Tonin P., Nicotera P., Bagetta G. Noi tendințe în controlul farmacologic al simptomelor neuropsihiatrice ale demenței. Curr. Opinează. Pharmacol. 2021; 61 :69–76. doi: 10.1016/j.coph.2021.09.002. [ PubMed ] [ CrossRef ] [ Google Scholar ]88.
Scuteri D., Guida F., Boccella S., Palazzo E., Maione S., Rodríguez-Landa JF, Martínez-Mota L., Tonin P., Bagetta G., Corasaniti MT Effects of Palmitoylethanolamide (PEA) on Durerea nociceptivă, musculo-scheletică și neuropatică: revizuire sistematică și meta-analiză a dovezilor clinice. Farmaceutică. 2022; 14 :1672. doi: 10.3390/pharmaceutics14081672. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]89.
Scuteri D., Rombolà L., Watanabe C., Sakurada S., Corasaniti MT, Bagetta G., Tonin P., Russo R., Nucci C., Morrone LA Impact of nutraceuticals on glaucoma: A systematic review. Prog. Brain Res. 2020; 257 :141–154. [ PubMed ] [ Google Scholar ]90.
Scuteri D., Contrada M., Loria T., Sturino D., Cerasa A., Tonin P., Sandrini G., Tamburin S., Bruni AC, Nicotera P., et al. Tratamentul durerii și al agitației la pacienții cu demență severă: nevoia de traducere, adaptare și validare a scalei durerii italiene de mobilizare-observare-comportament-intensitate-demență (I-MOBID2) cu teste psihometrice. Biomed. Pharmacother. 2022; 150 :113013. doi: 10.1016/j.biopha.2022.113013. [ PubMed ] [ CrossRef ] [ Google Scholar ]91.
Scuteri D., Contrada M., Loria T., Tonin P., Sandrini G., Tamburin S., Nicotera P., Bagetta G., Corasaniti MT Pharmacological Treatment of Pain and Agitation in Severe Dementa and Responsiveness to Change of Scala de durere italiană de mobilizare-observare-comportament-intensitate-demență (I-MOBID2): protocol de studiu. Brain Sci. 2022; 12 :573. doi: 10.3390/brainsci12050573. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]92.
Scuteri D., Matamala-Gomez M., Bottiroli S., Corasaniti MT, De Icco R., Bagetta G., Tonin P. Pain assessment and treatment in dementa at the time of coronavirus disease COVID-19. Față. Neurol. 2020; 11 :890. doi: 10.3389/fneur.2020.00890. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]93.
Pignolo L., Tonin P., Nicotera P., Bagetta G., Scuteri D. ROBOCOP (ROBOtic Care of Poststroke Pain): Study Protocol for a Randomized Trial to Assess Robot-Assisted Functional and Motor Recovery and Impact on Poststroke Pain Dezvoltare. Față. Neurol. 2022; 13 :813282. doi: 10.3389/fneur.2022.813282. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]94.
Scuteri D., Contrada M., Tonin P., Corasaniti MT, Nicotera P., Bagetta G. Dementia and COVID-19: A Case Report and Literature Review on Pain Management. Produse farmaceutice. 2022; 15 :199. doi: 10.3390/ph15020199. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]95.
Scuteri D., Tonin P., Nicotera P., Bagetta G., Corasaniti MT Considerații din lumea reală pentru antagoniștii receptorilor CGRP recent aprobați în îngrijirea migrenei. Expert Rev. Neurother. 2022; 22 :221–230. doi: 10.1080/14737175.2022.2049758. [ PubMed ] [ CrossRef ] [ Google Scholar ]96.
Scuteri D., Tonin P., Nicotera P., Vulnera M., Altieri GC, Tarsitano A., Bagetta G., Corasaniti MT Pooled Analysis of Real-World Evidence Supports Anti-CGRP mAbs and OnabotulinumtoxinA Combined Trial in Chronic Migraine . Toxine. 2022; 14 :529. doi: 10.3390/toxins14080529. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]97.
Contrada M., Cerasa A., Tonin P., Bagetta G., Scuteri D. Aromatherapy in Stroke Patients: Is it Time to Begin? Față. Comportament. Neurosci. 2021; 15 :749353. doi: 10.3389/fnbeh.2021.749353. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]98.
Silva GL, Luft C., Lunardelli A., Amaral RH, Melo DA, Donadio MV, Nunes FB, de Azambuja MS, Santana JC, Moraes CM, et al. Efectele antioxidante, analgezice și antiinflamatorii ale uleiului esențial de lavandă. Un. Acad. Sutiene. Cienc. 2015; 87 :1397–1408. doi: 10.1590/0001-3765201520150056. [ PubMed ] [ CrossRef ] [ Google Scholar ]99.
Scuteri D., Rombolà L., Tridico L., Mizoguchi H., Watanabe C., Sakurada T., Sakurada S., Corasaniti MT, Bagetta G., Morrone LA Proprietăți neurofarmacologice ale uleiului esențial de bergamotă pentru clinica managementul BPSD-urilor legate de durere. Curr. Med. Chim. 2019; 26 :3764–3774. doi: 10.2174/0929867325666180307115546. [ PubMed ] [ CrossRef ] [ Google Scholar ]100.
Van de Ven TJ, John Hsia HL Cauzele și prevenirea durerii cronice postchirurgicale. Curr. Opinează. Crit. Îngrijire. 2012; 18 :366–371. doi: 10.1097/MCC.0b013e3283557a7f. [ PubMed ] [ CrossRef ] [ Google Scholar ]101.
Kitade M., Nakajima H., Tsujikawa T., Noriki S., Mori T., Kiyono Y., Okazawa H., Matsumine A. Evaluation of (R)-[11C]PK11195 PET/MRI for Spinal Cord- Durerea neuropatică asociată la pacienții cu tulburări ale coloanei vertebrale cervicale. J. Clin. Med. 2023; 12 :116. doi: 10.3390/jcm12010116. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
Articole din
Jurnalul Internațional de Științe Moleculare sunt furnizate aici prin amabilitatea
Multidisciplinary Digital Publishing Institute (MDPI)
Rata declinului cognitiv la vârstnici este foarte variabilă. Un factor potențial care contribuie la declinul cognitiv accelerat este inflamația sistemică cronică, deoarece a fost legată de tulburări cognitive și risc crescut de demență. Anumiți factori ai stilului de viață, cum ar fi excesul de greutate corporală și comportamentul sedentar, pot exacerba o stare proinflamatoare la adulții în vârstă, ducând la o inflamație cronică de grad scăzut. Suplimentarea dietei cu curcumină, un compus polifenolic antiinflamator din turmericul condiment cu curry, este o abordare potențială pentru a preveni declinul cognitiv accelerat prin contracararea proceselor inflamatorii cronice. Deși efectele antiinflamatorii ale curcuminei sunt bine stabilite, potențialele beneficii cognitive ale curcuminei au fost descoperite mai recent. Mai multe studii epidemiologice și animale asupra efectului suplimentării cu curcumină asupra cogniției au arătat rezultate promițătoare; cu toate acestea, studiile controlate randomizate pe oameni sunt limitate. În această revizuire, am identificat 5 studii randomizate controlate, dintre care doar 2 au observat un efect benefic al suplimentării cu curcumină asupra cogniției prin îmbunătățirea memoriei de lucru. Examinând în mod critic metodologiile acestor studii, am identificat câteva limitări, una dintre acestea fiind că niciunul dintre studii nu a explorat posibilitatea ca mecanismele antiinflamatorii să mediaze beneficiile cognitive (adică, niciun studiu nu a testat participanții cu inflamație de grad scăzut sau cu inflamație măsurată). biomarkeri). Alți factori care influențează probabilitatea unor rezultate concludente includ alegerea populației de studiu (nedeteriorată din punct de vedere cognitiv în comparație cu afectată), durata studiului, doza de curcumină și biodisponibilitatea acesteia și baterie de teste neurocognitive. Pe baza acestor constatări, oferim recomandări pentru studii viitoare care să examineze potențialele beneficii cognitive ale curcuminei la om, care includ evaluarea efectelor acesteia asupra funcției vasodilatatoare endoteliale cerebrale și creșterea efectelor cognitive prin combinarea acestuia cu omega-3 cu lanț lung. n–3) acizi grași.
Cuvinte cheie: curcumină, cogniție, declin cognitiv, inflamație, obezitate, acizi grași polinesaturați omega-3 cu lanț lung, funcție cerebrovasculară
Îmbătrânirea înregistrează o scădere a abilităților cognitive, cum ar fi raționamentul conceptual, memoria, viteza de procesare și funcția executivă, în timp ce alte abilități, cum ar fi limbajul, sunt păstrate ( 1 ). Cu toate acestea, rata declinului cognitiv este foarte variabilă între indivizi. Prin urmare, este important să se facă distincția între declinul cognitiv normal legat de vârstă și declinul cognitiv mai rapid legat de boală ( 1 , 2 ). Un posibil factor care contribuie la aceasta din urmă este inflamația sistemică cronică. Creșterea dovezilor arată o legătură între profilurile proinflamatorii și afectarea cognitivă și riscul de demență ( 3 ). Îmbătrânirea este asociată cu o inflamație intensă și prelungită ( 3), care poate fi exacerbată de anumiți factori ai stilului de viață, cum ar fi comportamentul sedentar și excesul de greutate corporală, în special adipozitatea viscerală, ceea ce duce la inflamație cronică de grad scăzut ( 4 ) . Adipocitele nu sunt doar depozite de stocare a energiei, ci acționează și ca un organ endocrin, exprimând proteine proinflamatorii și recrutând macrofage în absența oricărei leziuni acute ( 5 ). S-a demonstrat că adulții supraponderali și obezi prezintă deficiențe ale funcției executive și ale memoriei pe termen scurt în comparație cu adulții cu un IMC normal, independent de vârstă ( 6 ). În plus, riscul de a dezvolta demență este crescut cu 35% la persoanele supraponderale și cu 74% la persoanele care sunt obeze la mijlocul vârstei (40-45 ani) ( 7 ) .). Comportamentul sedentar contribuie, de asemenea, la mediul inflamator prin eliberarea crescută de proteine proinflamatorii, cum ar fi IL-6 și proteina C-reactivă (CRP) ( 4 ), sporind astfel și mai mult starea inflamatorie a adulților în vârstă sedentari, obezi.
Inflamația sistemică cronică și îmbătrânirea exagerează disfuncția endotelială, inclusiv creșterea permeabilității barierei hemato-encefalice (BBB), ducând la extravazarea monocitelor periferice în spațiile perivasculare, așa cum se arată în studii recente care implică modele de șobolani obezi hrăniți cu o dietă bogată în grăsimi ( 8 ) . Aceasta, la rândul său, stimulează activitatea microglială. La activare, aceste celule imune înnăscute ale sistemului nervos central suferă o proliferare semnificativă, care poate fagocita în mod agresiv componentele sinaptice, afectând astfel plasticitatea sinaptică și neuronală ( 9 ). Inflamația sistemică accelerează îmbătrânirea microglielor, ceea ce poate crește răspunsul hiperactiv al acestora ( 10). Hiperactivarea sau disfuncția cumulativă a microgliei poate accelera deteriorarea neuronală prin remodelarea sinaptică extinsă și eliberarea de molecule neurotoxice, cum ar fi factorii proinflamatori, speciile reactive de oxigen și proteinazele, contribuind la descompunerea în continuare a BBB (11 , 12 ) . Hipocampul și ganglionii bazali – care sunt structuri importante pentru memorie, atenție, emoție și percepție – conțin mai multe enzime implicate în procesele inflamatorii; prin urmare, aceste structuri prezintă un risc crescut de deteriorare din cauza neuroinflamației ( 3). Trebuie remarcat faptul că inflamația sistemică nu numai că perturbă BBB în sine, ci modifică și funcția anumitor transportatori BBB, care pot perturba, de exemplu, clearance-ul proteinei amiloid-β (Aβ), promovând acumularea acesteia și formarea plăcilor de amiloid, care este unul dintre semnele distinctive patologice ale bolii Alzheimer (AD) ( 13 , 14 ). Plăcile de amiloid pot îmbunătăți și mai mult activarea microglială și pot induce secreția de citokine proinflamatorii, exacerbând răspunsul neuroinflamator [revizuit mai detaliat în altă parte ( 15) .)]. În plus, inflamația cronică sistemică poate afecta și vasodilatația mediată de endoteliu a vaselor cerebrale, rezultând o stare cronică de hipoperfuzie, care a fost legată de severitatea deteriorării cognitive la vârstnici ( 16 , 17 ). Astfel, neuroinflamația poate avea consecințe dăunătoare de anvergură pentru funcția neuronală, cerebrovasculară și, în consecință, cognitivă.
Studiile epidemiologice sugerează că utilizarea uşoară până la moderată a medicamentelor antiinflamatoare nesteroidiene ar putea întârzia apariţia demenţei ( 3 ). Cu toate acestea, sa demonstrat că utilizarea cronică a medicamentelor antiinflamatoare nesteroidiene crește riscul de insuficiență renală acută, accident vascular cerebral și infarct miocardic la adulții în vârstă ( 18 ). Prin urmare, o abordare alternativă pentru a contracara procesele inflamatorii cronice este necesară urgent pentru a preveni declinul cognitiv accelerat asociat bolii. O abordare potențială este creșterea consumului de bioactive dietetice antiinflamatorii, cum ar fi curcumina, un compus polifenolic care este concentrat în curcuma cu condiment ( Curcuma longa) .). Curcumina (diferuloilmetan) a fost folosită de secole în Asia ca medicament pe bază de plante datorită capacității sale de a reduce durerea asociată afecțiunilor inflamatorii, cum ar fi reumatismul ușor și durerile articulare, precum și afecțiunile pielii, cum ar fi psoriazisul (19 ) . Există dovezi substanțiale pentru efectele antiinflamatorii ale curcuminei din studiile preclinice și clinice ( 20 ). La oameni, s-a demonstrat că curcumina scade semnificativ concentrația de citokine proinflamatorii, cum ar fi TNF-α, IL-6, TGF-β și proteina chemoatractantă monocitară 1 (MCP-1) (21 , 22 ) . Mai mult, Panahi et al. ( 23) au arătat că suplimentarea dietei cu curcumină (1 g/zi timp de 8 săptămâni) a îmbunătățit capacitatea antioxidantă, a scăzut peroxidarea lipidelor și a redus concentrațiile de CRP la 117 adulți cu sindrom metabolic.
La modelele animale, s-a demonstrat că curcumina nu numai că reduce markerii inflamatori sistemici, ci și că îmbunătățește funcția cognitivă. Într-un model de șoarece de obezitate la mijlocul vârstei, suplimentarea cu curcumină (1000 mg ⋅ kg -1 ⋅ d -1 ) timp de 12 săptămâni a dus la concentrații reduse de CRP, precum și o funcție executivă îmbunătățită, sugerând o posibilă legătură ( 24 ). Cu toate acestea, o legătură directă între îmbunătățirea stării inflamatorii și beneficiile cognitive datorate curcuminei trebuie încă stabilită. Cu toate acestea, mai multe studii pe animale au arătat beneficiile cognitive ale curcuminei. De exemplu, curcumina a crescut capacitatea de achiziție a memoriei și a îmbunătățit memoria non-spațială la șoarecii și șobolanii în vârstă ( 25 , 26) .) și îmbunătățirea funcțiilor de lucru și a memoriei cu indici într-un stadiu mai avansat de afectare cognitivă (modelul mouse-ului pentru AD) ( 27 ). La oameni, studiile epidemiologice sugerează, de asemenea, o legătură între consumul de curcumină și beneficiile cognitive. Ng şi colab. ( 28 ) au observat că subiecții cu cantități mai mari de consum de curry au avut scoruri cu 6% mai mari la Mini-Mental State Examination (MMSE) în comparație cu subiecții care nu consumă niciodată sau rareori curry.
Deși rezultatele studiilor pe animale și epidemiologice sunt promițătoare, numărul de studii de intervenție umană este încă foarte limitat, deoarece utilizarea curcuminei pentru beneficiile cognitive umane este un concept relativ nou. Scopul acestei revizuiri a fost următorul: 1 ) de a oferi o imagine de ansamblu asupra studiilor de intervenție existente care examinează efectele curcuminei asupra funcției cognitive la oameni, 2 ) de a evalua critic metodologiile și constatările studiilor, 3 ) de a oferi o abordare potențială pentru a spori efectele cognitive ale curcuminei și 4 ) pentru a face recomandări pentru studii de intervenție viitoare.
Poate curcumina să contracareze declinul cognitiv?
Dovezi actuale din studiile clinice
Am identificat 5 studii controlate randomizate cu design paralel care examinează efectul curcuminei asupra cogniției la oameni, publicate între 2008 și 2016. Doar 2 din cele 5 studii au găsit un efect asupra cogniției. Am identificat, de asemenea, 1 studiu caz-control ( 29 ); cu toate acestea, nu vom raporta mai departe acest studiu, deoarece se concentrează pe efectele curcuminei asupra simptomelor comportamentale și psihologice ale demenței la 3 pacienți cu AD, mai degrabă decât asupra funcției cognitive în sine.
Primul studiu a fost realizat în 2008 de Baum et al. ( 30 ) la 27 de adulți în vârstă cu AD cărora li sa administrat 1000 mg sau 4000 mg curcumină/zi (încapsulat sau sub formă de pulbere, procentul de curcumină nea raportat) sau placebo timp de 6 luni (tabelul 1). Scorul MMSE al participanților nu s-a schimbat semnificativ atunci când au comparat niciunul dintre grupurile de tratament cu placebo.
TABELUL 1
Studii controlate randomizate cu design paralel care examinează efectele curcuminei asupra funcției cognitive 1
400 mg curcumină/zi [Longvida (Verdure Science)] sau placebo
Doză unică, 1 oră și 3 ore (acută) și 4 săptămâni (cronică)
Acut: tendință de îmbunătățire a atenției susținute ( P = 0,168) și îmbunătățire semnificativă a memoriei de lucru ( P = 0,033); cronică: îmbunătățire semnificativă a memoriei de lucru ( P = 0,044)
1500 mg curcumină/zi [Biocurcumax (Sabinsa Corporation)] sau placebo
12 luni (evaluare intermediară la 6 luni)
Nicio diferență semnificativă în performanța cognitivă (RAVLT, COWAT, WDSS și bateria de evaluări CogState); fără timp semnificativ × interacțiunea grupului de tratament pentru funcția cognitivă globală (testul MoCA) prin utilizarea liniei de bază și a performanței de 12 luni
1 AD, boala Alzheimer; ADAS-cog, Scala de evaluare a bolii Alzheimer, subporțiune cognitivă; COWAT, Test de asociere orală controlată a cuvintelor; MMSE, Mini-Examen de stat mental; MoCA, Montreal Cognitive Assessment; RAVLT, Testul Rey Auditiv de Învățare Verbală; ref, referință; WDSS, scara simbolului cifrelor Wechsler.
2 Comparativ cu placebo.
Un alt studiu la pacienții cu AD a fost realizat de Ringman și colab. ( 31 ) în 2012. Treizeci de adulți în vârstă cu AD ușoară până la moderată au consumat 2000 mg sau 4000 mg curcumină complex C3/zi (70–80% curcumină) sau placebo timp de 6 luni. Pe lângă MMSE, acest studiu a inclus și Scala de evaluare a bolii Alzheimer, subporțiunea cognitivă (ADAS-cog), dar nu a fost observat niciun efect semnificativ al curcuminei asupra cogniției.
Studiul lui Lee et al. ( 32 ) a fost efectuat la 48 de adulți cu prediabet netratat, cărora li s-a suplimentat o doză unică de 1 g turmeric (procentul de curcumină nu este raportat), 2 g scorțișoară, o combinație de turmeric și scorțișoară sau placebo. La momentul inițial și la 6 ore după suplimentare, memoria de lucru a fost evaluată cu testul N-Back. Indivizii suplimentați cu turmeric s-au dovedit a avea o memorie de lucru semnificativ îmbunătățită.
Studiul lui Cox et al. ( 33 ) a fost primul studiu, după cunoștințele noastre, care a investigat efectul curcuminei asupra performanței cognitive la adulții în vârstă sănătoși, dintre care 60 au fost suplimentați cu 400 mg Longvida/zi (80 mg curcumină/zi, Verdure Science (Noblesville, IN). , SUA)) sau placebo. O singură doză de curcumină a avut tendința de a spori atenția susținută și de a îmbunătăți semnificativ memoria de lucru. Îmbunătățirea semnificativă a memoriei de lucru a fost susținută după suplimentarea cronică cu curcumină timp de 4 săptămâni.
Un studiu mai recent a fost realizat în 2016 de Rainey-Smith et al. ( 34), care a suplimentat 96 de adulți sănătoși cu 1500 mg Biocurcumax/zi (conținând 88% curcuminoizi totale, Sabinsa Corporation (Piscataway, NJ, SUA)) sau placebo timp de 12 luni. Funcția cognitivă a fost măsurată la momentul inițial și după 6 și 12 luni. Acest studiu nu a găsit modificări semnificative ale funcției cognitive, măsurate prin scoruri compozite (ne)computerizate, după suplimentarea cu curcumină în comparație cu placebo. Pentru testul cognitiv global [Montreal Cognitive Assessment (MoCA)], a fost observată o interacțiune semnificativă timp × grup de tratament. Cu toate acestea, acest rezultat semnificativ s-a datorat în principal unei scăderi a scorului MoCA în grupul placebo după 6 luni, ceea ce nu a fost observat în grupul de tratament. Nu a fost observată nicio diferență în scorul MoCA între grupurile de tratament și placebo după 12 luni.
Lecții învățate din studiile clinice
Curcumina și efectele sale potențiale asupra funcției cognitive au atras atenția doar recent (<15 ani). Prin urmare, numărul de studii controlate randomizate care evaluează efectele curcuminei asupra cogniției la oameni este foarte limitat ( n = 5). Pe baza dovezilor existente, suplimentarea acută și pe termen scurt (4 săptămâni) cu curcumină pare să îmbunătățească funcția cognitivă prin îmbunătățirea memoriei de lucru, dar nu a fost observat niciun efect asupra întârzierii declinului cognitiv în studiile pe termen lung (6-12 luni). ). Mecanismele care stau la baza efectelor curcuminei asupra cogniției sunt în mare parte speculative și se bazează pe studii pe animale. Toate cele 5 studii au menționat efectele antiinflamatorii ale curcuminei și, prin urmare, beneficiul său potențial pentru îmbunătățirea funcției cognitive, dar niciunul dintre studii nu a inclus participanți care ar putea avea o inflamație cronică de grad scăzut și numai Cox și colab. ( 33) au măsurat efectiv markeri inflamatori. Cu toate acestea, deoarece cohorta lor a fost sănătoasă, majoritatea markerilor inflamatori la momentul inițial au fost sub nivelul de cuantificare. Astfel, o limitare majoră este că niciunul dintre studii nu a explorat legătura mecanicistă propusă pentru a media beneficiile cognitive ale curcuminei. În afară de aceasta, există mai multe motive posibile pentru lipsa efectelor semnificative ale curcuminei asupra cogniției în 3 din cele 5 studii.
Populația de studiu
Alegerea populației de studiu ar putea fi un posibil motiv pentru care majoritatea studiilor nu au găsit beneficii cognitive semnificative după suplimentarea cu curcumină. Primele 2 studii efectuate de Baum et al. ( 30 ) şi Ringman şi colab. ( 31), care a analizat efectele curcuminei asupra întârzierii declinului cognitiv, au fost exploratorii și efectuate la pacienții cu AD. Niciun studiu nu a observat o schimbare a scorului MMSE după 6 luni de suplimentare cu curcumină în comparație cu placebo. În ambele studii, majoritatea participanților luau medicamente pentru AD, posibil deja atenuând progresia bolii, făcând mai dificilă observarea oricărui efect potențial al suplimentării cu curcumină. O altă posibilitate este că patologia de bază a AD ar fi putut fi prea avansată pentru ca curcumina să poată inversa deteriorarea BBB, acumularea de Aβ și deteriorările funcției vasodilatatoare ( 13 , 14 , 35) .). Prin urmare, studiile viitoare ar trebui să ia în considerare evaluarea curcuminei la adulții în vârstă care pot prezenta unele deficite subiective de memorie din cauza declinului cognitiv legat de vârstă, dar care nu suferă încă de demență.
Durata procesului și alegerea testelor neurocognitive
O altă diferență majoră între studiile care au observat un efect semnificativ al suplimentării cu curcumină asupra cogniției și cele care nu au observat este scopul studiului și, în consecință, durata studiului. Lee şi colab. ( 32 ) și Cox și colab. ( 33 ) și-a propus să examineze dacă curcumina poate îmbunătăți funcția cognitivă, analizând efectele acute (1-6 ore) și pe termen scurt (4 săptămâni) ale suplimentării cu curcumină și au observat îmbunătățiri ale memoriei de lucru. Studiile lui Baum et al. ( 30 ), Ringman şi colab. ( 31 ) și Rainey-Smith și colab. ( 34) au avut o durată mai lungă (6-12 luni), deoarece au urmărit să vadă dacă curcumina ar putea întârzia declinul cognitiv. Niciunul dintre aceste studii nu a găsit un efect semnificativ al curcuminei. Cu toate acestea, perioada de studiu a fost probabil insuficientă pentru a detecta orice modificare a declinului cognitiv, măsurată prin modificarea scorului MMSE sau MoCA, deoarece nu s-a observat niciun declin cognitiv în grupul placebo. O altă alegere de test cognitiv sau o baterie de teste cognitive mai cuprinzătoare care examinează anumite domenii cognitive, mai degrabă decât cogniția globală, ar fi putut fi mai sensibilă pentru a detecta orice declin cognitiv și orice posibil efect al suplimentării cu curcumină în Baum și colab. ( 30 ) şi Ringman şi colab. ( 31 ) studii. Rainey-Smith și colab. ( 34) a folosit o baterie de teste cognitive mai cuprinzătoare, examinând memoria verbală episodică, funcția executivă, memoria de lucru, atenția și fluența verbală în plus față de cunoașterea globală. Cu toate acestea, lipsa rezultatelor cognitive în acest studiu s-ar putea datora altor motive, cum ar fi rata mare de abandon și diferențele dintre grupuri la momentul inițial. Grupul placebo a avut rezultate semnificativ mai bune la momentul inițial în MoCA, sarcina cu simbolul cifrelor și testele necomputerizate (comunite cu scorul compus necomputerizat), care ar fi putut masca orice efect al suplimentării cu curcumină asupra cogniției.
Doza de curcumină și uzura
Pe lângă durata studiului, o altă diferență majoră între studiile lui Lee și colab. ( 32 ) și Cox și colab. ( 33 ) în comparație cu studiile lui Baum și colab. ( 30 ), Ringman şi colab. ( 31 ) și Rainey-Smith și colab. ( 34 ) a fost doza de curcumină. Primele 2 studii au folosit doze mai mici de curcumină, de 400 și, respectiv, 1000 mg/zi, și ar putea fi posibil ca o doză mai mică să fie mai eficientă în producerea de beneficii cognitive. În plus, dozele mai mari de curcumină par să cauzeze mai multe probleme gastrointestinale, ducând la mai multe retrageri în timpul studiului. Cox şi colab. ( 33 ) nu a avut nicio retragere în timpul studiului de 4 săptămâni după randomizare, în timp ce Rainey-Smith și colab. ( 34) a avut o rată de abandon de 25% din cauza evenimentelor adverse, în principal gastrointestinale. Rata mare de abandon școlar ar fi putut afecta puterea lor statistică și ar fi putut duce la grupuri dezechilibrate, ambele ar putea explica lipsa rezultatelor semnificative.
Biodisponibilitatea curcuminei
Biodisponibilitatea slabă a curcuminei ar fi putut contribui, de asemenea, la lipsa rezultatelor semnificative în majoritatea studiilor. Se știe că curcumina este hidrofobă și are o absorbție intestinală scăzută, o distribuție tisulară scăzută, un metabolism rapid și un clearance sistemic rapid ( 19 ). O altă problemă este glucuronidarea curcuminei, deoarece curcumina trebuie să fie neconjugată pentru a traversa BBB și a provoca efecte în creier ( 36 ). Doar 2 din cele 5 studii incluse în această revizuire au măsurat efectiv concentrațiile de curcumină în plasmă. Baum şi colab. ( 30 ) nu a detectat curcumină liberă în plasmă, ceea ce indică faptul că aproape toată curcumina a fost glucuronidată. Ringman şi colab. ( 31) au folosit o metodă mai sensibilă pentru măsurarea curcuminei și au găsit, de asemenea, concentrații mult mai mari de curcumină glucuronidată în comparație cu concentrațiile native de curcumină în plasmă la 3 ore după administrare în săptămâna 24. Un alt studiu uman în care 6 voluntari au luat 2 g curcumină împreună cu 5 mg piperină – o inhibitor al glucuronidării hepatice și intestinale – a arătat o dublare a absorbției curcuminei ( 37 ), indicând faptul că glucuronidarea este cel puțin parțial responsabilă pentru biodisponibilitatea scăzută a curcuminei.
Acest lucru ridică nevoia unor formulări de curcumină mai eficiente. Cox şi colab. ( 33 ) a folosit o nouă formulă de curcumină cu biodisponibilitate îmbunătățită. Curcuminul Longvida folosește tehnologia particulelor de curcumină lipide solide pentru a furniza curcumină țesuturilor țintă. Această tehnologie dizolvă curcumina într-o particulă lipidică, crescând astfel stabilitatea acesteia prin protejarea acesteia de degradarea acidului gastric și creșterea solubilității acesteia prin îmbunătățirea absorbției curcuminei ( 38 ). Gota și colab. ( 39) a testat biodisponibilitatea curcuminei Longvida prin suplimentarea a 11 pacienți cu osteosarcom fie cu curcumină Longvida, fie cu un extract de curcuminoid 95%. Ei au arătat că la 2 ore după o administrare de 650 mg de curcumină Longvida, concentrațiile plasmatice au crescut la 20 ng/mL, în timp ce nu a fost detectată curcumină în plasmă după 650 mg de extract de curcuminoid 95%.
Astfel, pe lângă luarea în considerare a populației de studiu, durata studiului, doza de curcumină și alegerea testelor neurocognitive, studiile viitoare de intervenție ar trebui să utilizeze o formulare optimizată de curcumină, cum ar fi Longvida, care poate crește biodisponibilitatea, pentru a crește potențialul curcuminei de a îmbunătăți cognitiv. funcţie.
Pot PUFA n-3 cu lanț lung să sporească efectele curcuminei?
O altă abordare potențială pentru a crește eficacitatea curcuminei ar putea fi combinarea curcuminei cu alți nutrienți bioactivi. O astfel de combinație este cu PUFA cu lanț lung (LC) n–3, cum ar fi EPA sau DHA, care se găsesc în surse marine, cum ar fi uleiul de pește și fructele de mare, deoarece au, de asemenea, proprietăți antiinflamatorii și s-a dovedit că îmbunătățesc anumite domenii cognitive ( 40 , 41 ). Această combinație ar putea exercita efecte suplimentare sau complementare, așa cum a fost arătat de Kakoti și colab. ( 42), care a combinat LC n-3 PUFA cu resveratrol și a arătat efecte sinergice asupra inflamației. Atât curcumina, cât și LC n-3 PUFA au, de asemenea, efecte vasodilatatoare, care ar putea îmbunătăți și mai mult performanța cognitivă. Interesant, un studiu la șobolani a arătat că curcumina poate crește conținutul de DHA în ficat și creier prin creșterea sintezei de DHA din acidul α-linoleic precursor n-3 (18:3n-3) (43 ) . Astfel, există multiple mecanisme prin care curcumina ar putea acționa în combinație cu LC n-3 PUFA sau alți nutrienți bioactivi pentru a îmbunătăți performanța cognitivă.
Mecanisme potențiale
Efecte antiinflamatorii
Curcumina este un inhibitor puternic al semnalizării NF-κB – un factor de transcripție implicat în reglarea în sus a mediatorilor inflamatori – prin inhibarea IκB kinazei β (IKKβ), blocând astfel activarea în continuare în aval (44 , 45 ) . Acest lucru îi permite să inhibe expresia ciclooxigenazei 2 (COX-2) și a citokinelor proinflamatorii, cum ar fi IL-1 și IL-6. Mai mult, semnalizarea inflamatorie este redusă datorită curcuminei care inhibă legarea proteinei activatoare 1 la ADN. S-a demonstrat, de asemenea, că curcumina scade stresul oxidativ și reduce concentrațiile de proteine oxidate ( 23 , 46) .). În plus, curcumina poate afecta plăcile de proteine Aβ, un loc potențial de origine al proceselor inflamatorii cronice din creier. Agregarea proteinei Aβ, formând plăci de amiloid, începe cu zeci de ani înainte de apariția primelor simptome clinice de afectare cognitivă. S-a demonstrat că curcuminul reduce concentrațiile insolubile de Aβ, previne agregarea Ap și reduce sarcina plăcii Ap în studiile pe animale ( 47 ). Cu toate acestea, studiile lui Baum et al. ( 30 ) şi Ringman şi colab. ( 31) au măsurat concentrațiile de Aβ în sânge și lichidul cefalorahidian la pacienții cu AD și nu au găsit nicio modificare semnificativă după suplimentare. O posibilitate ar putea fi ca, la oameni, curcumina poate preveni agregarea Ap într-un stadiu incipient, dar nu este capabilă să reducă sarcina plăcii Ap în AD stabilită; cu toate acestea, acest lucru trebuie evaluat în continuare.
LC n–3 PUFA pot inhiba, de asemenea, NF-kB prin scăderea fosforilării lui IkB, care este necesară pentru ca NF-kB să se disocieze de IkB și să se transloce în nucleu (46 ) . Cu toate acestea, efectele antiinflamatorii ale LC n-3 PUFA sunt mediate în mare parte de modificări ale compoziției FA a membranelor celulare, ducând la modificarea expresiei genelor și a modelelor de producție de mediatori lipidici ( 40 ) .). Un aport crescut de LC n-3 PUFA are ca rezultat o încorporare scăzută a acidului arahidonic n-6 (AA) în membranele celulare. Atât AA, cât și EPA produc eicosanoide, care sunt molecule de semnalizare. Cu toate acestea, eicosanoizii produși din AA au un profil proinflamator ridicat, în timp ce cei produși din EPA sunt mult mai puțin puternici și prezintă un profil proinflamator scăzut. Mai mult, EPA și DHA dau naștere la rezoluvine, care sunt antiinflamatoare și au proprietăți de rezolvare a inflamației prin prevenirea infiltrației neutrofile la locurile de inflamație și inhibarea citokinelor proinflamatorii IL-1β și TNF-α. LC n–3 PUFA pot activa, de asemenea, PPAR-γ, care posedă proprietăți antiinflamatorii puternice ( 40 ).
Mai multe studii au arătat efecte negative ale citokinelor proinflamatorii asupra cogniției, sugerând o legătură între inflamația cronică sistematică și deficitele cognitive. De exemplu, un studiu observațional de 2 ani la participanți în vârstă de Yaffe și colab. ( 48) au arătat că cei din cele mai înalte tertiluri ale IL-6 și CRP au avut scoruri MMSE semnificativ mai mici la momentul inițial, precum și la urmărire și au avut un risc cu 24% mai mare de declin cognitiv în comparație cu participanții din cele mai mici tertile. Reducerea proceselor inflamatorii sistemice prin suplimentarea cu nutrienți ar putea ajuta la prevenirea sau inversarea leziunilor BBB, precum și la inhibarea activării microgliei, prevenind astfel eliberarea de substanțe neurotoxice care conduc la neuroinflamație și neurodegradare și, în consecință, contracarând declinul cognitiv accelerat. Atât curcumina, cât și LC n-3 PUFA pot inhiba procesele inflamatorii, dar acționează prin diferite mecanisme pentru a rezolva inflamația. Astfel, combinația lor ar putea ajuta la ameliorarea mai eficientă a inflamației și, prin urmare, să obțină un efect benefic mai mare asupra funcției cognitive.
Efecte asupra funcției vasodilatatoare endoteliale
Mai multe studii au examinat efectul suplimentării LC n-3 PUFA asupra funcției vasodilatatoare endoteliale, majoritatea observând o creștere a dilatației mediate de flux (FMD) ( 49 ). Acest efect este probabil multifactorial, deoarece LC n–3 PUFA pot crește producția de NO și prostaciclină, ambele vasodilatatoare endoteliale, și pot inhiba producția de tromboxan A2, un vasoconstrictor puternic și facilitator al agregării trombocitelor ( 50 , 51 ).
Studiile privind efectul curcuminei asupra vasodilatației, deși limitate, au arătat, de asemenea, o creștere a FMD după suplimentare ( 52 , 53 ). Studiul lui Pu et al. ( 54 ) au arătat că, la șobolani, suplimentarea cu curcumină a restabilit vasodilatația cerebrovasculară afectată prin calea AMP kinazei (AMPK). S-a demonstrat că curcumina promovează fosforilarea AMPK, ceea ce duce la o fosforilare crescută a NO sintazei endoteliale și, în cele din urmă, duce la creșterea producției de NO și a biodisponibilității ( 55 ). O biodisponibilitate crescută a NO a fost observată și într-un studiu uman realizat de Santos-Parker și colab. ( 56) în 2017, în care 39 de adulți în vârstă sănătoși au fost suplimentați cu 2000 mg Longvida/zi (400 mg curcumină/zi) timp de 12 săptămâni. Ei au arătat o creștere cu 36% a febrei aftoase în grupul de tratament, în timp ce grupul placebo nu a arătat nicio modificare a febrei aftoase. Pe lângă biodisponibilitatea crescută a NO, autorii sugerează că creșterea FMD se datorează și reducerii observate a stresului oxidativ.
Inflamația ar putea avea nu numai un efect negativ direct asupra cogniției, ci și un efect indirect prin promovarea disfuncției endoteliale, despre care s-a sugerat că contribuie la un declin cognitiv accelerat și la progresia demenței (57-59 ) . Legătura dintre disfuncția endotelială și declinul cognitiv ar putea fi mediată de o reacție cerebrovasculară afectată la cerințele cognitive, ceea ce duce la o aprovizionare insuficientă cu sânge în regiunile active ale creierului ( 60 ). Se sugerează că funcția cerebrovasculară slabă joacă un rol important în inițierea și progresia demenței ( 16 ). Un semn distinctiv al disfuncției endoteliale este biodisponibilitatea redusă a NO, ceea ce duce la afectarea funcției vasodilatatoare ( 61). S-a demonstrat că curcumina și LC n–3 PUFA îmbunătățesc FMD prin creșterea disponibilității NO. Cu toate acestea, ambii nutrienți au și alte căi (modularea producției de tromboxan sau inhibarea stresului oxidativ) care pot avea ca rezultat o funcție vasodilatatoare endotelială îmbunătățită, crescând posibilitatea unui efect aditiv al combinației de nutrienți asupra funcției vasodilatatoare.
Luate împreună, atât inflamația, cât și funcția vasodilatatoare afectată par să joace un rol important în procesul de declin cognitiv și progresia spre demență. Curcumina și LC n-3 PUFA posedă ambele efecte antiinflamatorii și sunt capabile să îmbunătățească funcția vasodilatatoare endotelială sistemică. Diferitele căi utilizate de fiecare nutrient pentru a obține aceste efecte pot duce la un beneficiu aditiv al combinației lor asupra funcției cognitive, în special în populațiile cu inflamație cronică de grad scăzut (de exemplu adulții în vârstă sedentari, supraponderali sau obezi).
Lacune în literatură și implicații pentru studiile viitoare
O lacună majoră în literatură este că, până acum, niciun studiu de intervenție nu a încercat să lege efectele antiinflamatorii ale curcuminei cu potențialele efecte cognitive benefice; de aceea, sunt necesare studii viitoare în acest domeniu. În plus, potențialul curcuminei de a îmbunătăți funcția vasodilatatoare ar trebui evaluat nu numai în circulația sistemică, ci și în cea cerebrală. Dacă curcumina este capabilă să îmbunătățească funcția endotelială în mod egal în circulația cerebrală și sistemică, acesta ar putea fi un alt mecanism potențial care explică beneficiile cognitive. Astfel de studii ar trebui să ia în considerare cu atenție populația studiată, durata studiului, doza și formula de curcumină și alegerea bateriei de teste cognitive.
Participanții cel mai probabil să beneficieze de suplimentarea cu curcumină ar putea fi cei cu inflamație cronică de grad scăzut, care sunt predispuși la declin cognitiv accelerat, cum ar fi adulții în vârstă supraponderali sau obezi și sedentari.
În funcție de rezultatul studiului (adică, beneficiile cognitive sau întârzierea declinului cognitiv), studiile viitoare ar trebui să ia în considerare durata optimă a studiului. Pentru beneficiile cognitive, studiile trebuie încă să valideze beneficiile cognitive pe termen scurt, precum și să investigheze efectele pe termen mai lung pentru a vedea dacă curcumina are beneficii în alte domenii cognitive, pe lângă memoria de lucru. Pentru a vedea dacă curcumina poate întârzia declinul cognitiv, sunt necesare studii viitoare cu perioade de studiu mai lungi (>2 ani), astfel încât să poată fi detectată o rată mai lentă de declin în grupul de tratament în comparație cu grupul placebo.
Sunt necesare studii doză-răspuns pentru a identifica doza optimă de curcumină pentru a obține beneficii cognitive. Se pare că dozele mai mici de curcumină ar putea fi favorabile, deoarece dozele mari de curcumină (>1,5 g/zi) sunt mai susceptibile de a provoca efecte adverse, predominant probleme gastrointestinale ( 30 , 31 , 34 ). Un studiu cu durata de 3 luni a stabilit o limită superioară de 12 g/zi pentru curcumină; nu se știe dacă acest lucru este valabil pentru o perioadă mai lungă de timp ( 62). Doze mai mici de curcumină ar putea fi mai eficiente și pot duce la mai puține evenimente adverse și, prin urmare, rate mai mici de abandon în timpul studiilor. Studiile viitoare ar putea lua în considerare și administrarea de curcumină de 2 ori/zi, pentru a menține o concentrație stabilă de curcumină în sânge, deoarece curcumina are un clearance sistematic rapid. Mai mult, datorită biodisponibilității scăzute a curcuminei, trebuie utilizate formulări de curcumină mai optimizate și trebuie monitorizată concentrația de curcumină din plasma sanguină. În plus, studiile viitoare ar trebui să examineze combinația de curcumină cu alți nutrienți bioactivi, cum ar fi LC n-3 PUFA, pentru a vedea dacă o combinație poate crește eficacitatea curcuminei pentru a îmbunătăți funcția cognitivă.
În plus, ar trebui utilizată o baterie cuprinzătoare de teste cognitive pentru a examina efectele curcuminei în diferite domenii cognitive, cum ar fi memoria de lucru, atenția, funcția executivă, limbajul și fluența verbală.
Testarea combinației de curcumină și LC n–3 PUFA
În prezent, desfășurăm un studiu de intervenție alimentară, dublu-orb, controlat cu placebo, de 16 săptămâni, pentru a investiga efectele suplimentării cu curcumină sau suplimentarea cu LC n-3 PUFA, sau ambele, asupra rezultatelor inflamatorii, circulatorii și cerebrovasculare și cognitive la persoanele supraponderale sau obeze. și adulți în vârstă sedentari fără tulburări cognitive (ACTRN12616000732482p, https://www.anzctr.org.au/Trial/Registration/TrialReview.aspx?id=370788&isReview=true). Participanții vor fi alocați să primească fie AGPI LC n–3 (total de 2,4 g/zi), curcumină Longvida (total de 160 mg/zi), fie o combinație de AGPI LC n–3 și curcumină sau placebo și vor fi instruiți să ia suplimentele de 2 ori/zi. Am ales această populație specială de studiu deoarece acești adulți sunt foarte probabil să aibă inflamație cronică de grad scăzut și mai multe studii au arătat deficiențe ale funcției endoteliale sistemice ( 63 ), fluxului sanguin cerebral regional în cortexul prefrontal ( 64 ) și reactivitatea cerebrovasculară la obezi. adulți ( 65 ). Aceste afectiuni sunt inca reversibile, spre deosebire de pacientii cu AD care prezinta rigiditate arteriala cerebrala si afectare vasodilatatoare intr-un stadiu atat de avansat incat este probabil ca endoteliul sa nu fie reparat.35 ). În acest studiu, vom testa ipoteza că combinația de curcumină și LC n-3 PUFA poate restabili funcția endotelială cerebrală, mediată parțial de o reducere a inflamației sistemice, care, la rândul său, poate îmbunătăți funcția cognitivă.
Funcția circulatorie cerebrală va fi evaluată prin măsurarea cu ultrasunete Doppler transcranian a vitezei fluxului sanguin în artera cerebrală medie în repaus și în timpul hipercapniei și testelor cognitive, reflectând receptivitatea vasodilatatoare dependentă de endoteliu a microvasculaturii cerebrale la acești stimuli fiziologici și mentali (de exemplu, cuplarea neurovasculară). capacitate).
Rezultatele cognitive includ o baterie cuprinzătoare de teste cognitive care evaluează cogniția globală (Examenul cognitiv III al lui Addenbrooke), memoria de lucru și episodică, viteza de procesare, funcția executivă, flexibilitatea cognitivă, controlul inhibitor, atenția și limbajul.
La momentul inițial și la 16 săptămâni, se va preleva o probă de sânge pentru a evalua markerii metabolici și inflamatori, precum și curcumina plasmatică și PUFA LC n-3 eritrocitare (indicele Omega-3).
Acesta este primul studiu, după cunoștințele noastre, cu o baterie cuprinzătoare de evaluări care examinează următoarele: 1 ) relația dintre îmbunătățirea funcției cerebrovasculare și performanța cognitivă după suplimentarea cu curcumină, 2 ) posibilele efecte aditive ale combinației de LC n-3 PUFA și curcumină asupra acestor rezultate și 3 ) efectele antiinflamatorii ale LC n-3 PUFA și curcumină într-o populație de așteptat să aibă inflamație cronică de grad scăzut și relația lor cu funcția cerebrovasculară și performanța cognitivă.
Dovezile pentru curcumina care afectează cogniția la oameni sunt foarte limitate și sunt necesare urgent studii viitoare pentru a investiga în continuare potențialele beneficii cognitive ale curcuminei. Pe baza studiilor existente, curcumina pare să aibă un efect benefic acut și pe termen scurt asupra memoriei de lucru. Dacă curcumina poate îmbunătăți alte domenii cognitive și poate întârzia declinul cognitiv, trebuie investigat în continuare în studiile de intervenție pe termen lung. Limitarea majoră a studiilor de intervenție existente este că nu au examinat legătura mecanicistă propusă (adică, efectele antiinflamatorii ale curcuminei contribuie la beneficiile sale cognitive). Studiile viitoare ar trebui să exploreze această legătură și să examineze dacă curcumina poate îmbunătăți nu numai funcția endotelială sistemică, ci și cerebrală. Învățând din limitările celor 5 studii de intervenție existente, studiile viitoare trebuie să ia în considerare cu atenție populația de studiu, durata studiului (în funcție de faptul dacă rezultatul studiului este îmbunătățirea cognitivă sau declinul cognitiv), doza și formularea curcuminei și alegerea bateriei de teste cognitive. Mai mult, studiile viitoare ar trebui să testeze combinații de curcumină cu alți nutrienți bioactivi, cum ar fi LC n-3 PUFA, care ar putea ajuta la creșterea eficacității curcuminei prin diferite mecanisme de rezolvare a inflamației și de creștere a funcției vasodilatatoare endoteliale, ceea ce duce la un posibil efect aditiv asupra performanței cognitive.
Autorii au raportat că nu au primit finanțare pentru această revizuire.
Dezvăluiri autor: JCK, RHXW și PRCH, fără conflicte de interese.
Abrevieri folosite:AAacid arahidonicApamiloid-βANUNȚboala AlzheimerBBBbariera hemato-encefalicăCRPproteina C-reactivaFMDdilatare mediată de fluxIKKβIκB kinaza βLClanț lungMMSEMini-Examinare de stare mentalăMoCAEvaluarea Cognitivă la Montreal
Park HL, O’Connell JE, Thomson RG. O revizuire sistematică a declinului cognitiv în populația generală în vârstă . Int J Geriatr Psihiatrie 2003; 18 ( 12 ): 1121–34. [ PubMed ] [ Google Scholar ]3.
Sartori AC, Vance DE, Slater LZ, Crowe M. Impactul inflamației asupra funcției cognitive la adulții în vârstă: implicații pentru practica și cercetarea în domeniul sănătății . J Neurosci Nurs 2012; 44 ( 4 ): 206–17. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]4.
Henson J, Yates T, Edwardson CL, Khunti K, Talbot D, Gray LJ, Leigh TM, Carter P, Davies MJ. Timp sedentar și markeri ai inflamației cronice de grad scăzut într-o populație cu risc ridicat . PLoS One 2013; 8 ( 10 ): e78350. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]5.
Greenberg AS, Obin MS. Obezitatea și rolul țesutului adipos în inflamație și metabolism . Am J Clin Nutr 2006; 83 (Supliment):461S–5S. [ PubMed ] [ Google Scholar ]6.
Nguyen JC, Killcross AS, Jenkins TA. Obezitatea și declinul cognitiv: rolul inflamației și al modificărilor vasculare . Front Neurosci 2014; 8 :375. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]7.
Whitmer RA, Gunderson EP, Barrett-Connor E, Quesenberry CP Jr., Yaffe K. Obesity in middle age and future risk of dementa: a 27 years longitudinal population based study . BMJ 2005; 330 ( 7504 ):1360. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]8.
Buckman LB, Hasty AH, Flaherty DK, Buckman CT, Thompson MM, Matlock BK, Weller K, Ellacott KL. Obezitatea indusă de o dietă bogată în grăsimi este asociată cu creșterea pătrunderii celulelor imune în sistemul nervos central . Brain Behav Immun 2014; 35 :33–42. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]9.
Miller AA, Spencer SJ. Obezitatea și neuroinflamația: o cale către afectarea cognitivă . Brain Behav Immun 2014; 42 :10–21. [ PubMed ] [ Google Scholar ]10.
Wyss-Coray T. Îmbătrânirea, neurodegenerarea și întinerirea creierului . Natura 2016; 539 ( 7628 ):180–6. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]11.
Dheen ST, Kaur C, Ling EA. Activarea microglială și implicațiile sale în bolile creierului . Curr Med Chem 2007; 14 ( 11 ): 1189–97. [ PubMed ] [ Google Scholar ]12.
da Fonseca AC, Matias D, Garcia C, Amaral R, Geraldo LH, Freitas C, Lima FR. Impactul activării microgliale asupra barierei hematoencefalice în bolile creierului . Front Cell Neurosci 2014; 8 :362. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]13.
Erickson MA, Banks WA. Disfuncția barierei hematoencefalice ca cauză și consecință a bolii Alzheimer . J Cereb Blood Flow Metab 2013; 33 ( 10 ): 1500–13. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]14.
Selkoe DJ, Hardy J. Ipoteza amiloidă a bolii Alzheimer la 25 de ani . EMBO Mol Med 2016; 8 ( 6 ):595–608. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]15.
Marques F, Sousa JC, Sousa N, Palha JA. Bariere hemato-encefalice în îmbătrânire și în boala Alzheimer . Mol Neurodegener 2013; 8:38 . [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]16.
Ruitenberg A, den Heijer T, Bakker SL, van Swieten JC, Koudstaal PJ, Hofman A, Breteler MM. Hipoperfuzia cerebrală și debutul clinic al demenței: Studiul Rotterdam . Ann Neurol 2005; 57 ( 6 ):789–94. [ PubMed ] [ Google Scholar ]17.
Sun ZW, Zhu YX, Liu HY, Liu J, Zhu XQ, Zhou JN, Liu RY. Scăderea vitezei fluxului sanguin cerebral la purtătorii de alele apolipoproteinei E epsilon4 cu afectare cognitivă ușoară . Eur J Neurol 2007; 14 ( 2 ): 150–5. [ PubMed ] [ Google Scholar ]18.
Marcum ZA, Hanlon JT. Recunoașterea riscurilor consumului cronic de medicamente antiinflamatoare nesteroidiene la adulții în vârstă . Ann Longterm Care 2010; 18 ( 9 ):24–7. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]19.
Shakeri A, Sahebkar A. Formule optimizate de curcumină pentru tratamentul bolii Alzheimer: o evaluare a pacientului . J Neurosci Res 2016; 94 ( 2 ): 111–3. [ PubMed ] [ Google Scholar ]20.
Jurenka JS. Proprietățile antiinflamatorii ale curcuminei, un constituent major al Curcuma longa: o revizuire a cercetării preclinice și clinice . Altern Med Rev 2009; 14 ( 2 ): 141–53. [ PubMed ] [ Google Scholar ]21.
Panahi Y, Hosseini MS, Khalili N, Naimi E, Simental-Mendia LE, Majeed M, Sahebkar A. Efectele curcuminei asupra concentrațiilor serice de citokine la subiecții cu sindrom metabolic: o analiză post-hoc a unui studiu controlat randomizat . Biomed Pharmacother 2016; 82 :578–82. [ PubMed ] [ Google Scholar ]22.
Sahebkar A, Cicero AF, Simental-Mendia LE, Aggarwal BB, Gupta SC. Curcumina reglează în jos nivelurile factorului alfa de necroză tumorală umană: o revizuire sistematică și o meta-analiză a studiilor controlate randomizate . Pharmacol Res 2016; 107 :234–42. [ PubMed ] [ Google Scholar ]23.
Panahi Y, Hosseini MS, Khalili N, Naimi E, Majeed M, Sahebkar A. Efecte antioxidante și antiinflamatorii ale combinației curcuminoid-piperină la subiecții cu sindrom metabolic: un studiu controlat randomizat și o meta-analiză actualizată . Clin Nutr 2015; 34 ( 6 ): 1101–8. [ PubMed ] [ Google Scholar ]24.
Sarker MR, Franks S, Sumien N, Thangthaeng N, Filipetto F, Forster M. Curcumin imită efectele neurocognitive și antiinflamatorii ale restricției calorice într-un model de șoarece de obezitate la mijlocul vârstei . PLoS One 2015; 10 ( 10 ):e0140431. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]25.
Dong S, Zeng Q, Mitchell ES, Xiu J, Duan Y, Li C, Tiwari JK, Hu Y, Cao X, Zhao Z. Curcumina îmbunătățește neurogeneza și cogniția la șobolanii în vârstă: implicații pentru interacțiunile transcripționale legate de creștere și sinaptică plasticitate . PLoS One 2012; 7 ( 2 ): e31211. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]26.
Yu SY, Zhang M, Luo J, Zhang L, Shao Y, Li G. Curcumina ameliorează deficitele de memorie prin sintaza neuronală de oxid nitric la șoarecii în vârstă . Prog Neuropsychopharmacol Biol Psychiatry 2013; 45 :47–53. [ PubMed ] [ Google Scholar ]27.
Cheng KK, Yeung CF, Ho SW, Chow SF, Chow AH, Baum L. Nanoparticule de curcumină foarte stabilizate testate într-un model de barieră hematoencefalică in vitro și la șoarecii Tg2576 cu boala Alzheimer . AAPS J 2013; 15 ( 2 ): 324–36. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]28.
Ng TP, Chiam PC, Lee T, Chua HC, Lim L, Kua EH. Consumul de curry și funcția cognitivă la vârstnici . Am J Epidemiol 2006; 164 ( 9 ):898–906. [ PubMed ] [ Google Scholar ]29.
Hishikawa N, Takahashi Y, Amakusa Y, Tanno Y, Tuji Y, Niwa H, Murakami N, Krishna UK. Efectele turmericului asupra bolii Alzheimer cu simptome comportamentale și psihologice ale demenței . AYU 2012; 33 ( 4 ):499–504. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]30.
Baum L, Lam CW, Cheung SK, Kwok T, Lui V, Tsoh J, Lam L, Leung V, Hui E, Ng C și colab. Test clinic pilot de șase luni, randomizat, controlat cu placebo, dublu-orb, cu curcumină la pacienții cu boala Alzheimer . J Clin Psychopharmacol 2008; 28 ( 1 ): 110–3. [ PubMed ] [ Google Scholar ]31.
Ringman JM, Frautschy SA, Teng E, Begum AN, Bardens J, Beigi M, Gylys KH, Badmaev V, Heath DD, Apostolova LG, et al. Curcumină orală pentru boala Alzheimer: tolerabilitate și eficacitate într-un studiu randomizat, dublu orb, controlat cu placebo, de 24 de săptămâni . Alzheimers Res Ther 2012; 4 ( 5 ):43. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]32.
Lee MS, Wahlqvist ML, Chou YC, Fang WH, Lee JT, Kuan JC, Liu HY, Lu TM, Xiu L, Hsu CC și colab. Turmericul îmbunătățește memoria de lucru postprandială în pre-diabet, independent de insulină . Asia Pac J Clin Nutr 2014; 23 ( 4 ): 581–91. [ PubMed ] [ Google Scholar ]33.
Cox KH, Pipingas A, Scholey AB. Investigarea efectelor curcuminei lipide solide asupra cogniției și dispoziției la o populație în vârstă sănătoasă . J Psychopharmacol 2015; 29 ( 5 ): 642–51. [ PubMed ] [ Google Scholar ]34.
Rainey-Smith SR, Brown BM, Sohrabi HR, Shah T, Goozee KG, Gupta VB, Martins RN. Curcumină și cogniție: un studiu randomizat, controlat cu placebo, dublu-orb al adulților în vârstă care locuiesc în comunitate . Br J Nutr 2016; 115 ( 12 ): 2106–13. [ PubMed ] [ Google Scholar ]35.
Lim JS, Lee JY, Kwon HM, Lee YS. Corelația dintre pulsatilitatea arterială cerebrală și disfuncția cognitivă la pacienții cu boala Alzheimer . J Neurol Sci 2017; 373 :285–8. [ PubMed ] [ Google Scholar ]36.
Hickey MA, Zhu C, Medvedeva V, Lerner RP, Patassini S, Franich NR, Maiti P, Frautschy SA, Zeitlin S, Levine MS, et al. Îmbunătățirea neuropatologiei și a deficitelor transcripționale la șoarecii Knock-in CAG 140 susține un efect benefic al curcuminei alimentare în boala Huntington . Mol Neurodegener 2012; 7:12 . [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]37.
Anand P, Kunnumakkara AB, Newman RA, Aggarwal BB. Biodisponibilitatea curcuminei: probleme și promisiuni . Mol Pharm 2007; 4 ( 6 ): 807–18. [ PubMed ] [ Google Scholar ]38.
Gota VS, Maru GB, Soni TG, Gandhi TR, Kochar N, Agarwal MG. Siguranța și farmacocinetica unei formulări solide de particule lipidice de curcumină la pacienții cu osteosarcom și la voluntari sănătoși . J Agric Food Chem 2010; 58 ( 4 ):2095–9. [ PubMed ] [ Google Scholar ]40.
Mazereeuw G, Lanctot KL, Chau SA, Swardfager W, Herrmann N. Efectele acizilor grași omega-3 asupra performanței cognitive: o meta-analiză . Neurobiol Aging 2012; 33 ( 7 ): 1482, e17–29. [ PubMed ] [ Google Scholar ]42.
Kakoti BB, Hernandez-Ontiveros DG, Kataki MS, Shah K, Pathak Y, Panguluri SK. Resveratrol și acizi grași omega-3: implicațiile sale în bolile cardiovasculare . Front Cardiovasc Med 2015; 2:38 . [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]43.
Wu A, Noble EE, Tyagi E, Ying Z, Zhuang Y, Gomez-Pinilla F. Curcumin stimulează DHA în creier: Implicații pentru prevenirea tulburărilor de anxietate . Biochim Biophys Acta 2015; 1852 ( 5 ):951–61. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]44.
Kim SG, Veena MS, Basak SK, Han E, Tajima T, Gjertson DW, Starr J, Eidelman O, Pollard HB, Srivastava M și colab. Tratamentul cu curcumină suprimă activitatea IKKbeta kinazei a celulelor salivare ale pacienților cu cancer de cap și gât: un studiu pilot . Clin Cancer Res 2011; 17 ( 18 ):5953–61. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]45.
Marquardt JU, Gomez-Quiroz L, Arreguin Camacho LO, Pinna F, Lee YH, Kitade M, Dominguez MP, Castven D, Breuhahn K, Conner EA și colab. Curcumina inhibă eficient semnalizarea oncogenă NF-kappaB și restrânge caracteristicile stemness în cancerul hepatic . J Hepatol 2015; 63 ( 3 ): 661–9. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]46.
Gardener SL, Rainey-Smith SR, Martins RN. Dieta și inflamația în boala Alzheimer și bolile cronice asociate: o revizuire . J Alzheimers Dis 2016; 50 ( 2 ): 301–34. [ PubMed ] [ Google Scholar ]47.
Goozee KG, Shah TM, Sohrabi HR, Rainey-Smith SR, Brown B, Verdile G, Martins RN. Examinarea valorii clinice potențiale a curcuminei în prevenirea și diagnosticarea bolii Alzheimer . Br J Nutr 2016; 115 ( 3 ): 449–65. [ PubMed ] [ Google Scholar ]48.
Yaffe K, Lindquist K, Penninx BW, Simonsick EM, Pahor M, Kritchevsky S, Launer L, Kuller L, Rubin S, Harris T. Markeri inflamatori și cogniție la bătrâni afro-americani și albi care funcționează bine . Neurologie 2003; 61 ( 1 ): 76–80. [ PubMed ] [ Google Scholar ]49.
Wang Q, Liang X, Wang L, Lu X, Huang J, Cao J, Li H, Gu D. Efectul suplimentării cu acizi grași omega-3 asupra funcției endoteliale: o meta-analiză a studiilor controlate randomizate . Ateroscleroza 2012; 221 ( 2 ): 536–43. [ PubMed ] [ Google Scholar ]50.
Zanetti M, Grillo A, Losurdo P, Panizon E, Mearelli F, Cattin L, Barazzoni R, Carretta R. Omega-3 polynesaturated fatty acids: structural and functional effects on the vascular wall . Biomed Res Int 2015; 2015 :791978. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]52.
Akazawa N, Choi Y, Miyaki A, Tanabe Y, Sugawara J, Ajisaka R, Maeda S. Ingestia de curcumină și antrenamentul fizic îmbunătățesc funcția endotelială vasculară la femeile aflate în postmenopauză . Nutr Res 2012; 32 ( 10 ): 795–9. [ PubMed ] [ Google Scholar ]53.
Oliver JM, Stoner L, Rowlands DS, Caldwell AR, Sanders E, Kreutzer A, Mitchell JB, Purpura M, Jager R. Forma nouă de curcumină îmbunătățește funcția endotelială la indivizii tineri și sănătoși: un studiu dublu-orb controlat cu placebo . J Nutr Metab 2016; 2016 :1089653. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]54.
Pu Y, Zhang H, Wang P, Zhao Y, Li Q, Wei X, Cui Y, Sun J, Shang Q, Liu D și colab. Curcumina alimentară ameliorează disfuncția cerebrovasculară legată de îmbătrânire prin calea AMPK/proteina de decuplare 2 . Cell Physiol Biochem 2013; 32 ( 5 ): 1167–77. [ PubMed ] [ Google Scholar ]55.
Xia N, Forstermann U, Li H. Resveratrol și oxid nitric endotelial . Molecule 2014; 19 ( 10 ): 16102–21. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]56.
Santos-Parker JR, Strahler TR, Bassett CJ, Bispham NZ, Chochol MB, Seals DR. Suplimentarea cu curcumină îmbunătățește funcția endotelială vasculară la adulții sănătoși de vârstă mijlocie și în vârstă prin creșterea biodisponibilității oxidului nitric și reducerea stresului oxidativ . Aging (Albany NY) 2017; 9 ( 1 ): 187–208. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]57.
Dede DS, Yavuz B, Yavuz BB, Cankurtaran M, Halil M, Ulger Z, Cankurtaran ES, Aytemir K, Kabakci G, Ariogul S. Assessment of endothelial function in Alzheimer’s disease: is Alzheimer’s disease a vascular disease? J Am Geriatr Soc 2007; 55 ( 10 ): 1613–7. [ PubMed ] [ Google Scholar ]58.
Forman DE, Cohen RA, Hoth KF, Haley AP, Poppas A, Moser DJ, Gunstad J, Paul RH, Jefferson AL, Tate DF și colab. Sănătatea vasculară și funcția cognitivă la adulții în vârstă cu boli cardiovasculare . Artery Res 2008; 2 ( 1 ): 35–43. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]59.
Vendemiale G, Romano AD, Dagostino M, de Matthaeis A, Serviddio G. Endothelial dysfunction associated with mild cognitive impairment in elderly population . Aging Clin Exp Res 2013; 25 ( 3 ): 247–55. [ PubMed ] [ Google Scholar ]60.
Kuszewski JC, Wong RHX, Howe PRC. Efectele acizilor grași polinesaturați omega-3 cu lanț lung asupra funcției vasodilatatoare endoteliale și asupra cogniției — sunt ele interdependente? Nutrienți 2017; 9 ( 5 ):487–508. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]61.
Sena CM, Pereira AM, Seica R. Endothelial dysfunction—a major mediator of diabetic vascular disease . Biochim Biophys Acta 2013; 1832 ( 12 ): 2216–31. [ PubMed ] [ Google Scholar ]62.
Shankar SS, Steinberg HO. Obezitatea și disfuncția endotelială . Semin Vasc Med 2005; 5 ( 1 ): 56–64. [ PubMed ] [ Google Scholar ]64.
Willeumier KC, Taylor DV, Amen DG. IMC crescut este asociat cu scăderea fluxului sanguin în cortexul prefrontal folosind imagistica SPECT la adulții sănătoși . Obezitate (Primăvara Argint) 2011; 19 ( 5 ): 1095–7. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]65.
Rodriguez-Flores M, Garcia-Garcia E, Cano-Nigenda CV, Cantu-Brito C. Relația dintre obezitate și rezistență la insulină cu reactivitatea cerebrovasculară: un studiu de caz și control . Cardiovasc Diabetol 2014; 13 :2. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]
Articole din
Advances in Nutrition sunt furnizate aici prin amabilitatea
Axa microbiota-intestin-creier joacă un rol important în dezvoltarea bolilor neurodegenerative. Bacteriile enterice comensale și patogene pot influența funcția creierului și a sistemului imunitar prin producerea de lipopolizaharide și amiloid. Disbioza microbiomului intestinal induce inflamație locală și consecutiv sistemică mediată imun. Citokinele proinflamatorii declanșează apoi neuroinflamația și în final neurodegenerarea. Stresul oxidativ mediat de imun poate duce la o deficiență de vitamine și micronutrienți esențiali. În plus, compoziția greșită a microbiotei intestinale ar putea afecta aportul și metabolizarea nutrienților. La pacienții cu boala Alzheimer (AD) au fost demonstrate modificări semnificative ale microbiotei intestinale. Dieta occidentală standard, infecții, scăderea activității fizice și stresul cronic influențează compoziția și diversitatea microbiotei intestinale. O abundență mai mare de microbiotă intestinală „proinflamatoare” este asociată cu o inflamație sistemică îmbunătățită și cu procese neuroinflamatorii. Astfel, AD care începe în intestin este strâns legată de dezechilibrul microbiotei intestinale. Modularea microbiotei intestinale prin dieta mediteraneană, probioticele și curcumina poate încetini declinul cognitiv și poate modifica semnificativ microbiomul intestinal. O abordare de intervenție multi-domeniu care abordează cauzele subiacente ale AD (inflamație, infecții, modificări metabolice cum ar fi rezistența la insulină și deficiența de nutrienți, stres) pare foarte promițătoare pentru a reduce sau chiar a inversa declinul cognitiv prin exercitarea efectelor pozitive asupra microbiotei intestinale.
1. Beta amiloid și neuroinflamație în boala Alzheimer
Boala Alzheimer (AD) este cea mai comună formă de demență la populațiile în vârstă din întreaga lume. O incidență și prevalență crescândă a AD în societățile occidentale vor merge împreună cu o povară socioeconomică ridicată în următoarele câteva decenii, dacă nu sunt dezvoltate strategii eficiente de prevenire [ 1 , 2 ]. Depunerea extracelulară de beta amiloid (Aβ) (plăci neuritice) și acumularea intracelulară de tau hiperfosforilat (p-Tau; încurcături neurofibrilare) sunt încă principalele semne neuropatologice ale AD [3 ]]. Ap se formează din proteina precursor de amiloid (APP), o proteină multifuncțională care este foarte conservată de-a lungul evoluției și exprimată în neuroni și glia. În hipocampul uman, Aβ este esențial pentru învățare și formarea memoriei și îmbunătățește consolidarea memoriei [ 4 ]. Astfel, producția de Ap este necesară pentru procesele de învățare care sunt afectate la pacienții cu AD. Când AD progresează, memoria și orientarea spațială se înrăutățesc continuu [ 5 ] până când pacienții nu mai pot avea grijă de ei înșiși și au nevoie de întreținere ridicată.
Depozitele de Aβ au fost demonstrate în creierul pacienților cu AD [ 6 ], dar și în creierul pacienților vârstnici fără demență [ 7 ]. Aβ poate activa microglia – celulele imune înnăscute ale creierului pentru a induce un răspuns neuroinflamator. Acest răspuns inițial benefic în sistemul nervos central (SNC) poate avea efecte dăunătoare, dacă continuă pentru perioade mai lungi, deoarece declanșează eliberarea cronică de citokine proinflamatorii [ 8 ]. Aceste citokine hrănesc apoi cascade neuroinflamatorii cronice, care de-a lungul anilor duc la neurodegenerare ireversibilă, în special în hipocamp.
Cu toate acestea, date recente indică faptul că nu numai cascadele neuro-inflamatorii, ci și modificările microbiotei intestinale joacă un rol crucial în dezvoltarea neurodegenerării: datele care investighează microbiota intestinală la oameni sugerează cu tărie că neuroinflamația este strâns legată de disbioza intestinală (așa cum este revizuită în capitolele următoare).
2. Microbiota intestinală alterată în boala Alzheimer
Fiecare individ, asemănător cu amprentele digitale, are propriul său microbiom care îi influențează sănătatea în mod important [ 9 , 10 ]. În principal intestinele, mai departe pielea, plămânii, cavitatea bucală și nazală și alte medii mucoase sunt colonizate de o cantitate enormă de microorganisme, inclusiv bacterii, viruși, ciuperci și paraziți [11 , 12 , 13 ] . Studii recente descriu peste 5000 de taxoni bacterieni care locuiesc în intestinul uman, mai ales Firmicutes și Bacteroidetes . Numărul acestor microorganisme poate depăși 10 14— asta înseamnă că avem de aproximativ 10 ori mai multe bacterii în noi decât celulele umane. Cantitatea de conținut genomic al microbiotei noastre este chiar de 100 de ori mai mare decât cea a genomului uman [ 14 ] – deși aceste cifre sunt puse sub semnul întrebării de alții [ 15 ].
Copiii nascuti pe cale vaginala oglindesc microbiota similara cu vaginul mamei, in timp ce bebelusii nascuti prin cezariana prezinta taxoni intestinali similari cu cei asociati cu pielea. La vârsta de aproximativ 2,5 ani, diversitatea microbiotei intestinale o reflectă pe cea a microbiotei adulte și rămâne relativ stabilă în timpul vârstei adulte [ 16 ]. Produsele secretoare ale microbiotei pot fi puternici activatori proinflamatori și imunitari înnăscut [ 17 ], pe de altă parte, un microbiom intestinal divers și echilibrat poate produce, de asemenea, acizi grași cu lanț scurt (SCFA) antiinflamatori [ 18 ].
Microbiota intestinală cuprinde o comunitate de microorganisme care au multe funcții importante pentru sănătatea gazdei lor (vezi alte recenzii [ 19 ]): înving agenții patogeni invadatori, metabolizează nutrienții, produc neurotransmițători și reglează sistemul imunitar. În ultimii ani, a fost recunoscut rolul crucial al microbiotei intestinale pentru dezvoltarea diferitelor boli precum infecții, boli metabolice, autoimune și mentale [ 20 ]. O diversitate redusă și compoziția greșită a microbiomului intestinal par să joace un rol important în patogeneza acestor boli.
De fapt, studii recente au arătat o asociere a dezvoltării bolilor neurodegenerative precum AD cu modificări ale microbiotei intestinale [ 19 , 21 ]. De fapt, studiile cu pacienți cu AD și controale sănătoase efectuate la Centrul de Cercetare pentru boala Alzheimer au arătat diferențe semnificative în ceea ce privește compoziția microbiomului intestinal [ 22 ]. Pacienții cu AD au avut un număr scăzut de Firmicutes și Actinobacteria , în timp ce numărul de bacterii aparținând phyla Proteobacteria și Bacterioidetes a fost crescut și legat de severitatea AD [ 22 ]. În plus, Enterobacteriaceaeau fost asociate cu prezența și progresia AD. Numărul de „bacterii proinflamatorii” cum ar fi Gammaproteobacteria , Enterobacteriales și Enterobacteriaceae din filum Proteobacteria a crescut constant de la controale sănătoase la deficiență cognitivă ușoară și stadiu de demență. Aceste modificări s-au corelat, de asemenea, semnificativ cu severitatea clinică a AD [ 23 ].
Interesant este că nivelurile diferitelor bacterii intestinale au fost corelate cu markeri cefalorahidiani pentru AD, cum ar fi Ab42/Ab40 sau p-Tau [ 22 ]. În plus, un studiu chinezesc a confirmat diferențe semnificative în compoziția microbiotei intestinale între indivizii sănătoși și pacienții cu AD [ 24 ]. Cu toate acestea, schimbările calitative nu au fost comparabile în China și SUA. Probabil, diferitele diferențe în ceea ce privește stilul de viață, obiceiurile alimentare, etnia sau comorbiditățile explică cel mai bine această constatare.
Mai mult, datele din experimentele pe animale sugerează cu tărie că microbiota este implicată în dezvoltarea AD: studiile preclinice la șoareci fără germeni au arătat o reducere semnificativă a Aβ cerebrală în comparație cu șoarecii cu microbiotă intestinală [25 ] . Mai mult, transplantul de probe de fecale de la șoareci AD la șoareci fără germeni a fost asociat cu o creștere semnificativă a nivelurilor Aβ [ 26 ].
3. Disbioza intestinală, sistemul imunitar și neuroinflamația în boala Alzheimer
Sistemul imunitar este modulat de microbiota intestinală: mai mult de 70% din celulele imune disponibile sunt prezente în intestin [ 27 ] și disbioza microbiotei intestinale influențează dramatic „ecosistemul uman”: deoarece bacteriile reglează permeabilitatea intestinelor, de exemplu, prin secretarea de mucus, creșterea excesivă a microorganismelor „greșite” sau doar diversitatea redusă a microbiotei poate promova un „intestin permeabil”. În acest caz, invadarea agenților patogeni, lipopolizaharidele (LPS) și, de asemenea, anumiți metaboliți pot părăsi intestinul și intra în sânge. Ca răspuns la acest „atac” microbian, sistemul imunitar eliberează citokine proinflamatorii pentru a inhiba bacteriemia.
În timp ce această strategie a sistemului imunitar este necesară și foarte eficientă în situațiile acute, reacțiile inflamatorii cronice (persistente) ale mucoasei intestinale pot supraîncărca sistemul imunitar și pot duce la un răspuns imunitar cronic, supraactiv, dar ineficient. Acestea ar trebui să meargă împreună cu dezvoltarea multor boli cronice, cum ar fi bolile inflamatorii intestinale, diabetul, ateroscleroza și, de asemenea, bolile neurodegenerative [ 19 ]. Translocarea microbiană cronică poate duce, de asemenea, la insuficiență de organ la distanță și leziuni [ 28 ]. În plus, translocarea bacteriană și secreția de citokine pro-inflamatorii afectează semnificativ integritatea barierei hematoencefalice (BBB) și induc cascade neuroinflamatorii [ 29 , 30 , 31] .]. Microbii de la periferie pot pătrunde în creier sau pot reactiva periodic microbii din infecția latentă a țesutului cerebral, trecând astfel la inflamația cronică dăunătoare cu supraproducție de Ap și depunere sub formă de plăci senile. Atât microbii, cât și produsele lor, amiloidul și LPS [ 32 ] se pot infiltra în creier și pot iniția neuroinflamația. Prin urmare, disbioza microbiotei intestinale nu afectează numai intestinele, de exemplu, printr-o formare scăzută de SCFA [ 33 , 34 ] și producția de vitamine [ 35 ], ci influențează, de asemenea, echilibrul dintre microbiotă și celulele imune în mod crucial (figura 1).
Disbioza intestinală promovează inflamația intestinală și sistemică cu, în consecință, agregarea Ap și neuroinflamația conducând în final la neurodegenerare și boala Alzheimer. Abrevieri: Aβ = beta amiloid; PP = polifenoli; SCFA = acizi grași cu lanț scurt; TMAO = N -oxid de trimetilamină; IL = interleukină; TNF = factor de necroză tumorală; BBB = bariera hematoencefalică.
Dovezile tot mai mari sugerează că neuroinflamația cu componentele sale imune poate fi o cauză centrală a agregării Ap, hiperfosforilării tau și în cele din urmă a morții neuronale [ 36 , 37 ]. Citokinele proinflamatorii — inițial benefice și neuroprotectoare — provoacă în cele din urmă neurodegenerare progresivă [ 38 ]. Astfel, reacțiile inflamatorii ar putea fi atât benefice, numite euflamație [ 39 ], cât și dăunătoare creierului: în funcție de amploarea și, de asemenea, stadiul activării, neurodegenerarea poate fi intensificată ușor sau puternic.
Reacțiile inflamatorii cronice duc la supraactivarea microglială prin eliberarea unor cantități mari de agenți inflamatori, așa cum se observă în diferite neuropatologii [ 40 ]. În cele din urmă, dezechilibrul acestor răspunsuri inflamatorii inițiază procesul neurodegenerativ [ 41 , 42 ]. Răspunsurile neuroinflamatorii consecutive pot îmbunătăți apoi procesele neurodegenerative [ 36 , 43 ]. În condiții de disbioză, au fost descriși receptori de declanșare exprimați pe celulele mieloide (TREM-1/2), care unesc procesele inflamatorii în tulburările intestinale și neurodegenerative prin răspândirea compușilor inflamatori prin axa microbiotă-intestin-creier [44] .]. Astfel, pe lângă factorii de risc primari pentru AD, cum ar fi înaintarea în vârstă [ 45 ], inflamația intestinală cronică poate juca, de asemenea, un rol important [ 44 , 46 ].
Inflamația intestinală poate fi măsurată prin evaluarea concentrației de calprotectină din scaun, care se găsește crescută la pacienții cu dement [ 47 ]. Nivelurile de calprotectină sunt, de asemenea, crescute semnificativ în lichidul cefalorahidian și creierul pacienților cu AD, promovând agregarea amiloidului și co-agregarea cu Aβ [ 37 , 48 ]. Semnele activării imune pot fi detectate concomitent în ser (neopterina) și probele fecale (S100A12) ale pacienților cu AD [ 49 , 50 ].
Studiile anterioare au arătat că proteina S100 care leagă calciul A8 (S100A8) și S100A9 sunt co-exprimate într-o varietate de stări inflamatorii, iar nivelurile plasmatice crescute de S100A9 sunt asociate cu tulburări inflamatorii [51 ] . S100A9 proinflamator este secretat de macrofage și microglia în timpul formării plăcii de amiloid [ 48 ].
Disbioza intestinală și inflamația intestinală sunt direct asociate cu disfuncția barierei intestinale și cu creșterea permeabilității intestinale („intestin permeabil”) și, prin urmare, pot contribui la procesul de neurodegenerare [ 52 , 53 ]. Disbioza bacteriilor intestinale reduce integritatea barierei intestinale, permițând astfel invazia ușoară a bacteriilor patogene. Infecția cu agentul patogen condiționat Citrobacter, de exemplu, s-a dovedit că induce tulburări de memorie la șoareci, care au fost împreună cu scăderea expresiei factorului neurotrofic derivat din creier (BDNF) [ 54 ]. În mod interesant, scăderea nivelurilor de BDNF a fost demonstrată și în creierul și serul pacienților cu AD [ 55 ].
4. Influența metaboliților microbieni asupra permeabilității intestinale, a sistemului imunitar și a producției de neurotransmițători în boala Alzheimer
Diferiți microbi ai microbiotei umane generează cantități mari de amiloid, iar biodisponibilitatea acestuia la SNC crește odată cu vârsta [ 56 ]. Bacteriile Gram negative precum E. coli și Salmonella spp . produc fibre amiloide extracelulare cunoscute sub numele de curli printr-un sistem de secreție cu mai multe componente [ 57 ]. Curli sunt inductori puternici ai răspunsului inflamator al gazdei [ 58 ]. În plus, ciupercile produc LPS, amiloizi și alte exsudate microbiene [ 56 , 57] .]. Microbiota intestinală produce și eliberează continuu fibre proteice extracelulare pentru a crea și menține un biofilm, deoarece aceste fibre oferă protecție împotriva factorilor de stres din mediu; în plus, ele mediază aderența atât la suprafețele biotice, cât și la cele abiotice [ 59 ].
Amiloizii microbieni și cerebrali, similari din punct de vedere biologic și chimic în ceea ce privește modelul molecular asociat patogenului (PAMP), nu împărtășesc secvențele de aminoacizi cu Aβ1-42 uman [ 56 ]. Oricum, ele sunt recunoscute de același sistem receptor și activează, de asemenea, citokine proinflamatorii precum interleukina (IL)-17 și IL-22. Acest lucru poate fi dăunător în timpul îmbătrânirii, când mucoasa gastrointestinală, precum și BBB devin continuu mai permeabile [ 56 , 60 , 61] .]. Acești activatori proinflamatori puternici derivați de microbiom induc citokine proinflamatorii, activarea complementului și modifică imunogenitatea în creier, sporind agregarea amiloidului și reacțiile inflamatorii. Ambele, proteinele amiloide și LPS, sunt activatori puternici ai receptorului pentru produsele finale de glicație avansată (RAGE) și receptorii de tip Toll (TRL), perpetuând inflamația cronică în AD [62 ] .
Cu toate acestea, microbiota intestinală nu secretă numai metaboliți pro-inflamatori, dar sunt, de asemenea, capabile să producă diverși compuși „antiinflamatori” și imunomodulatori, cum ar fi butiratul SCFA, în funcție de compoziția și diversitatea lor [63 ] . Microbii intestinali comensali fermentează fibrele alimentare în SCFA, crescând astfel producția de proteine de joncțiune strânsă claudin-5 și ocludină, care sunt importante pentru o funcție de barieră strânsă și selectivă. În conformitate cu aceste date, s-ar putea demonstra că o abundență mai mare de bacterii producătoare de butirat și concentrații mai mari de butirat sunt protectoare împotriva invaziei Citrobacter rodentium la rozătoare [ 64 ]. În consecință, suplimentarea cu butirat a inhibat puternic creșterea Citrobacter rodentium[ 64 ]. Prin urmare, aceste date implică faptul că butiratul poate sprijini funcția de barieră a celulelor epiteliale intestinale și poate crește integritatea joncțiunii [ 65 ], reducând astfel invazia enteropatogenilor în fluxul sanguin și minimizând translocarea LPS în intestine [ 66 ]. În plus, butiratul este capabil să inhibe căile importante în producerea de citokine proinflamatorii [ 67 ] și, de asemenea, îmbunătățește semnalizarea insulinei [ 68 ].]. Foarte interesant, un studiu clinic recent ar putea arăta că amiloidoza creierului (cuantificată prin PET amiloid Florbetapir) a fost asociată semnificativ cu niveluri crescute de LPS din sânge și citokine proinflamatorii și invers corelată cu citokina antiinflamatoare IL-10 și butiratul SCFA. [ 69 ]. În plus, markerii disfuncției endoteliale au fost mai mari la pacienții cu niveluri crescute de citokine proinflamatorii, în timp ce nivelurile de butirat și IL-10 au fost reduse [ 69 ]. Inflamația sistemică indusă de LPS ar putea, prin urmare, să impună activarea endotelială și formarea Aβ [ 69 ].
La pacienții obezi, numărul bacteriilor producătoare de butirat numite Faecalibacterium prausnitzii a fost descris a fi semnificativ mai mic în comparație cu persoanele sănătoase cu greutate normală [ 70 ]. Pe de altă parte, un număr mai mare de Faecalibacterium prausnitzii merge împreună cu concentrații mai scăzute de citokine proinflamatorii [ 71 ] și CRP [ 72 ]. Interesant este că pacienții cu un IMC crescut (>25) cu niveluri mai mari de CRP au avut, de asemenea, o abundență mai mică de Lactobacili și Bifidobacterii , în timp ce speciile Escherichia coli și Bacteroides au fost mai abundente la aceiași pacienți [ 73] .]. Obezitatea și rezistența la insulină sunt de fapt un factor de risc major pentru declinul cognitiv [ 74 ], în special la persoanele în vârstă.
La indivizii în vârstă se observă o reducere marcată a biodiversității microbiene, care se caracterizează printr-o abundență mai mare de Proteobacterii și un număr scăzut de specii de Bifidobacterii și, consecutiv, printr-o scădere semnificativă a formării SCFA [ 41 , 75 , 76 ].
În cazul disbiozei legate de vârstă sau de dietă, influențele „metaboliților microbieni intestinali benefici” asupra integrității intestinale și asupra BBB se pierd [ 29 , 77 ]. Prin urmare, disbioza merge împreună cu o permeabilitate crescută a barierelor intestinale [ 78 ]. În plus, scade, de asemenea, permeabilitatea BBB [ 79 ] și sinteza și secreția de factori neurotrofici, cum ar fi factorul de creștere a nervilor derivati din creier (BDNF) și receptorii N -metil D-aspartat (NMDA) [ 80 ]. Nivelurile reduse de BDNF și semnalizarea NMDA sunt asociate cu declinul cognitiv [ 80 ].
La indivizii sănătoși, microbiota comensală produce în plus precursori ai unui număr de alți neurotransmițători importanți: acetilcolină, catecolamine, acid gama-aminobutiric, precum și metaboliți triptofan, chinurenină, melatonină și serotonină [41 ] . De fapt, mulți dintre acești neurotransmițători sunt implicați central în controlul semnalizării axei intestin-creier [ 81 , 82 ]. O revizuire recentă a lui Bosi și colegii [ 83 ] descrie modul în care metaboliții căii chinureninei, de exemplu, acidul chinolinic și acidul chinurenic, influențează activitatea neuronală în SNC și în periferie prin exercitarea efectelor neurotoxice și neuroprotective: producția de chinurenină a gazdei poate fi influențată de microbiota prin modularea activității TDO indusă de glucocorticoizi [84 ] și activitatea IDO mediată imun [ 85 ]. Foarte interesant, la rozătoarele fără germeni (GF), nivelurile plasmatice ale triptofanului cresc, în timp ce raportul chinurenină-triptofan crește (datorită modificărilor activității IDO și TDO) [33 , 86 , 87 , 88 ] . În mod interesant, administrarea de probiotice precum Bifidobacterium infantis normalizează raportul chinurenină-triptofan [ 87 ]. Dereglarea metabolismului triptofanului/chinureninei ar putea contribui în mod semnificativ la tulburările axei microbiotă-intestin-creier și ar putea facilita dezvoltarea simptomelor neuropsihiatrice [ 33] .]. Beneficiile microbiene ale altor cataboliți de triptofan au fost, de asemenea, revizuite recent [ 89 ]: indolii și triptamina, de exemplu, au fost descrise de Roager și colegii de muncă pentru a activa sistemul imunitar, a îmbunătăți bariera epitelială intestinală și a stimula motilitatea gastrointestinală [89 ] . În plus, acești cataboliți de triptofan derivați de bacterii intestinale sunt implicați în secreția hormonilor intestinali și pot avea efecte sistemice antiinflamatorii, antioxidante sau toxice [ 89 ].
La pacienții cu AD a fost demonstrată o creștere a raportului chinurenină/triptofan, care merge împreună cu nivelurile crescute de zonulină (ca marker pentru scăderea funcției barierei intestinale), iar suplimentarea cu un probiotic multi-tulpina a fost eficientă pentru a scădea nivelurile de zonulină, a crește abundența Faecalibacterium prausnitzii . și modifică raportul chinurenină/triptofan [ 90 ]. Prin urmare, probioticele ar putea exercita efecte benefice asupra gazdei prin modularea metabolismului triptofanului gazdei, prin creșterea producției de neurotransmițători și, probabil, de asemenea, prin susținerea proliferării tulpinilor bacteriene „antiinflamatorii” precum Faecalibacterium prausnitzii .
Pe de altă parte, dezechilibrul „ecosistemului microbiotei intestinale” uman poate avea, de asemenea, efecte semnificative asupra sistemului nervos enteric (ENS), o parte foarte importantă a sistemului nervos autonom. ENS, care poate fi privit ca un „al doilea creier”, este important pentru motilitatea gastrointestinală (GI), precum și pentru modularea răspunsului imun intestinal în sănătate și boală. Este important pentru activarea celulelor T și răspunsul la pericolul celular prin secretarea de citokine și molecule imunomodulatoare, creând un răspuns neuroinflamator și mecanisme de moarte neuronală. Aceste răspunsuri sunt localizate în principal în ENS, dar prin axa intestin-creier pot afecta și SNC [ 91 , 92 ]. Atât în ENS, cât și în SNC, aceleași proteine pliate greșit inițiază neurodegenerarea [ 93].
În plus, studii recente demonstrează că microbiota intestinală are un impact asupra reflexului inhibitor neuroinflamator mediat de sistemul nervos colinergic [ 94 ]. De fapt, pierderea neuronilor colinergici localizați în nucleul bazal al lui Meynert este asociată cu deficite cognitive la pacienții cu AD [ 95 ].
5. Rolul infecției parodontale, alți agenți patogeni infecțioși și antibiotice și AD
În plus, inflamația parodontală cronică poate contribui la modificări ale microflorei intestinale [ 96 , 97 , 98 , 99 ] și poate exacerba răspunsul inflamator sistemic al gazdei [ 97 , 100 , 101 ]. În cavitatea bucală sănătoasă comensalii rămân în echilibru fiziologic, dar tulburările acestei stări ecologice pot duce la comunități microbiene parodontale disbiotice și parodontită [ 96 ]. Parodontita este legată de mai multe boli sistemice, cum ar fi ateroscleroza, diabetul zaharat și obezitatea [ 96 , 102 , 103] .]. Saliva înghițită a pacienților cu parodontită conține cantități mari de bacterii: până la 10 12 bacterii pe zi [ 98 ]. Parodontita poate provoca secreția de agenți patogeni bacterieni precum LPS, flagelina, peptidoglicanul și alte molecule proinflamatorii, care sunt toți factori de risc modificabili pentru AD [ 104 , 105 ]. În consecință, parodontita este o boală inflamatorie disbiotică care poate media inflamația atât la locurile locale, cât și la distanțe, inclusiv creierul [ 96 , 103 , 106 ]. Efectele neuroinflamatorii pot fi provocate prin căi celulare, umorale și neuronale, care toate pot contribui la patogenia AD [ 96 , 103 ,106 ] (vezi de asemeneaFigura 2).
Modificarea microbiomului (de exemplu, prin dieta occidentală) care duce la disbioză intestinală duce la inflamație de grad scăzut în intestin și la creșterea permeabilității intestinale și BBB și consecutiv la neuroinflamație și declin cognitiv; Patobionții orali precum P. gingivalis conduc la oralizarea microbiotei intestinale, pe de o parte, conducând astfel, în plus, inflamația intestinală și, pe de altă parte, promovând procesele neuroinflamatorii prin translocarea bacteriilor în creier prin proteaze toxice. Abrevieri: IL = interleukină, TNF = factor de necroză tumorală; BBB = bariera hematoencefalică.
Într-un studiu pe animale, 5 săptămâni de administrare de Porphyromonas gingivalis (P. gingivalis) au dus la modificări ale compoziției microbiotei intestinale prin creșterea proporțiilor de celule Th17 intestinale în limfocitele mezenterice [ 107 ]. În plus, P. gingivalis a scăzut sensibilitatea la insulină și a crescut citokinele proinflamatorii TNF- α și IL-6, provocând un răspuns inflamator la nivelul ficatului prin formarea de picături de lipide, indicând că parodontita poate afecta boala ficatului gras nealcoolic (NAFLD; [ 108 ]). Într-un studiu de investigație recent, a fost observată o asociere semnificativă între prezența salivară a P. gingivalis și rezultate mai proaste la testele cognitive [ 101 ].
Infecția cu Aggregatibacter actinomycetemcomitans (A. actinomycetemcomitans) , un alt parodontopatogen a îmbunătățit steatoza hepatică la modelele de șoarece. Anticorpii A. actinomycetemcomitans au apărut și mai mari la pacienții cu NAFLD [ 109 ]. Studiile anterioare au arătat că starea dentară slabă și boala parodontală sunt legate de funcția cognitivă redusă și AD [ 101 , 110 , 111 ]. Poole și colegii au confirmat că LPS de la P. gingivalis poate intra în creierul AD, sugerând un rol inflamator în patologia AD. Ei bine, în conformitate cu aceste date, invazia activă a agentului patogen oral P. gingivalis în creier ar putea fi demonstrată în modele de șoarece [112 , 113 ]. Descoperiri recente descriu că bacteriile orale, cum ar fi speciile Klebsiella rezistente la antibiotice , pot coloniza intestinul și pot induce inflamația intestinală cronică prin activarea sistemului imunitar intestinal [ 97 , 99 ]. Astfel, tractul oral și gastrointestinal reprezintă amploarea încărcăturii microbiene umane, oferind oportunități noi și avansate de diagnostic și terapeutic în ceea ce privește neuroinflamația și neurodegenerarea [ 37 , 98 , 114 ].
Cu toate acestea, nu numai translocarea bacteriană ar putea contribui la neuroinflamație, dar și infecțiile virale ar putea juca un rol. Infecțiile virale duc adesea la o inducere puternică a răspunsului imun și au fost demonstrate concentrații crescute de neopterina la pacienții cu infecții cu virusul Epstein-Barr (EBV) și citomegalovirus (CMV) [ 115 , 116 ]. O revizuire recentă afirmă că infecțiile cu herpesvirusuri induc producția de Aβ, fosforilarea tau, stresul oxidativ și neuroinflamația în general [ 117 ]. Interesant, peptida Aβ are proprietăți antimicrobiene și este capabilă, de exemplu, să inhibe replicarea HSV-1 [ 117]. Astfel, reactivarea herpesvirusurilor ar putea iniția reacții în cascadă de amiloid la persoanele în vârstă vulnerabile, poate induce stres oxidativ și, în final, neuroinflamație cu acumularea de Aβ și p-Tau [ 118 ]. În plus, CMV, un alt membru al familiei virusului herpes, a fost descris pentru a promova demența [ 119 ]. Mai mult, de asemenea, o interacțiune între CMV și HIV-1 a fost descrisă ca fiind asociată cu un risc mai mare de a dezvolta AD [ 45 , 120 , 121 ]. Deoarece se știe că infecțiile virale induc eliberarea de citokine proinflamatorii, ele ar putea contribui și la neuroinflamație prin declanșarea secreției sistemice, dar și locale de citokine.
Ipoteza că agenții infecțioși declanșează dezvoltarea AD a fost deja propusă de Aloysius Alzheimer și propusă din nou în urmă cu câțiva ani [ 106 ]. Un studiu recent discută din nou această ipoteză și afirmă că studiile de autopsie și de asociere oferă dovezi ale poverii infecțioase (în principal bacterii precum Chlamydia pneumoniae și spirochetele și virusurile herpes) [ 122 ]. În conformitate cu ipoteza „infecției”, 1194 de pacienți care au supraviețuit sepsisului sever au avut o nouă afectare cognitivă substanțială și persistentă și dizabilitate funcțională [ 123] .]. S-a propus ca acumularea de Aβ pliat greșit în creier ca răspuns la infecție să fie responsabilă pentru declinul cognitiv al pacienților. Pe de altă parte, experimentele modelelor in vitro și in vivo sugerează mai degrabă că Ap, care este și un agent antimicrobian, ar putea fi produs pentru a preveni infecția creierului [ 124 ]. De fapt, mecanismele de protecție inițial pentru inhibarea proliferării agenților patogeni (de exemplu, prin formarea Aβ sau activarea imună), ar putea duce, de-a lungul timpului și în timpul stărilor prelungite de deficiență de nutrienți, la acumularea excesivă de Aβ în creier și la epuizarea cronică a sistemului imunitar. similar cu imunodeficiența cronică în infecția cu HIV).
As infections appear to contribute to gut dysbiosis, systemic inflammation and neuroinflammation, anti-infective treatment might be a reasonable therapeutic option. Antibiotics have saved thousands of lives, as they are very potent drugs that inhibit the proliferation of pathogens. On the other hand, they were also shown to affect cognition and memory function in rodent models and humans depending on the type of antibiotic and its targeting microbiome. Possible effects of antibiotics on the gut–brain axis and their possible influence on AD have been reviewed recently [125]: while amoxicillin, rifampicin, minocycline, rapamycin or doxycycline improved cognition, reduced Aβ deposition and accumulation of hyperphosphorylated tau, streptozotocin, ampicillin and cefepime were associated with memory deficits, reduced consciousness and confusion. [125]. In this review, a very important issue was also raised: the antibiotic streptozotocin is used to induce sporadic AD forms in animal models with effects on learning and memory performances [126]. Furthermore, streptozotocin is also employed to induce diabetes mellitus in animals [127], which is a frequent comorbidity of AD going along with early cognitive decline [128]. However, whether effects of antibiotics on cognition are due to microbiotic changes following antibiotic therapy or are caused by the antibiotic itself is not proven yet.
The use of antibiotics has furthermore also been related to depression and anxiety—which are frequently encountered in AD patients—by changing the gut microbiota and the brain–gut axis (see review by Hao et al. [129], Desbonnet et al. [130]), suggesting that we should use antibiotics wisely and only when indicated. Furthermore, long term effects of antibiotic treatment should be taken into consideration, especially in young children:
The infectiologist and microbiome researcher Martin Blaser was one of the first researchers proposing that losses of particular bacterial species of our ancestral microbiota (by over-use of antibiotics) have altered the context, in which immunological, metabolic and cognitive development occur in early life [131]. He also corroborated the hypothesis that “missing/disappearing microbes” contribute importantly to the epidemics of chronic disease [131], as the early use of antibiotics in infants goes along with substantial alterations of the gut microbiota and with the rise of the “modern plagues”: obesity, childhood diabetes, allergic diseases, chronic intestinal bowel disease and eczema [132].
Additionally, antibiotic treatment of very young mice induced cognitive deficits, altered dynamics of the tryptophan metabolic pathway, and significantly reduced BDNF, oxytocin and vasopressin expression in the adult brain [130]. Further studies investigating effects of antibiotics on the microbiota and cognitive and mental development of babies and infants should therefore be conducted to address this important topic.
6. Western Diet: Effects on Gut Microbiota, Inflammation, Metabolism and the Brain
Typically, human microbiota remain stable during longer periods of time [98,133], but changes in diet as well as antibiotic treatment can result in dysbiosis [134,135]. Standard Western diet (WD) with its high intake of refined sugars/grains, red and processed meat, saturated fats and a high consumption of sugared beverages goes along with substantial and fundamental changes of the gut microbiota (see also reviews by Noble et al. and Al Bander et al. [19,21]). The high contents of animal protein in WD appears to provide an optimal environment for anaerobic microorganisms and specific genera including Bacteroides and Bilophila [136 ]. Pe de altă parte, speciile Prevotella par să domine la indivizii care mănâncă multe plante [ 66 ] cu o mai mare diversitate a microbiotei fecale în comparație cu indivizii care consumă WD obișnuit [ 137 ]. În studiile pe animale, hrănirea rozătoarelor cu o dietă cu un consum ridicat de grăsimi saturate, sare și zahăr („dieta occidentală”) a crescut grăsimea abdominală, rezistența la insulină, ateroscleroza și inflamația [138 , 139 , 140 , 141 ] . În cele ce urmează, vor fi descrise efectele specifice ale WD asupra inflamației, precum și asupra metabolismului glucozei și grăsimilor:
6.1. WD și inflamație
WD are un potențial proinflamator semnificativ (figura 1), care a fost asociat cu boli precum bolile maligne și cardiovasculare [ 142 , 143 ]. O meta-analiză foarte recentă cu date nutriționale colectate prospectiv de > 200.000 de persoane ar putea arăta că o dietă „pro-inflamatoare” care conține cantități mari de cereale rafinate/zahăr/băuturi cu zahăr, carne procesată/roșie și grăsimi saturate este însoțită de markeri de inflamație mai mari. precum hsCRP și receptorul TNF-α 2 și o supraviețuire semnificativ mai proastă [ 144 ]. Persoanele care consumau o mulțime de legume, fructe, cereale integrale, ceai verde, vin și cafea, pe de altă parte, au prezentat markeri de inflamație mai mici și au avut un risc semnificativ mai scăzut pentru evenimente cardiovasculare [144] .]. Legumele și cerealele integrale sunt bogate în fibre, microbiotă accesibilă (adică, carbohidrați complecși) și substanțe bioactive puternice, care s-au demonstrat că au efecte benefice (vezi recenzia lui Moszak și colab. [145] ) . În studiile pe animale, WD scade și nivelul creierului de neurotrofine, cum ar fi BDNF și reduce capacitatea de a învăța [ 146 ].
WD poate crește neuroinflamația prin producția mai mare de LPS și prin reducerea numărului de bacterii comensale „antiinflamatoare” [ 147 ]: șoarecii care au primit transplanturi fecale cu dieta occidentală au avut niveluri crescute de endotoxină și markeri neuroinflamatori și o funcție cognitivă afectată [ 147 ]. În plus, WD a redus abundența A. muciniphilia , o specie de microbiotă intestinală „antiinflamatoare” [ 147 ].
6.2. WD și efectele sale asupra metabolismului și a creierului
Consumul de prea multe cereale rafinate/zahăr și băuturi cu zahăr este, de asemenea, împreună cu episoadele de hiperglicemie intermitentă și cu formarea crescută de produse finale de glicație avansată (AGE), care au fost, de asemenea, descrise ca joacă un rol proeminent în dezvoltarea AD [148, 149 ] . ]. AGE-urile declanșează formarea mai multor citokine proinflamatorii, cum ar fi factorul de necroză tumorală-alfa (TNF-α), interleukina (IL)-1 și IL-6 [150], care sunt bine stabilite pentru a agrava neuroinflamația și procesele de neurodegenerare și pentru a reduce și mai mult hipocampul productiv pentru adulți. neurogeneza [ 151 ]. AGE-urile activează, de asemenea, microglia prin interacțiunea cu receptorul său RAGE și eliberarea de specii reactive de oxigen (ROS) [ 152 ] (figura 1).
Cantități mari de zahăr rafinat în WD și niveluri crescute constant de glucoză pot duce la un dezechilibru al semnalizării insulinei [ 153 ], și anume rezistența la insulină, care ar putea fi crucială pentru progresia AD, deoarece insulina modifică activitatea neuronală [ 154 ] și îmbunătățește memoria. [ 155 ]. Interesant, s-a demonstrat că insulina reduce neuroinflamația, de exemplu, prin atenuarea eliberării de citokine induse de LPS în creier și a inflamației sistemice [ 156 ]. Mai multe studii au confirmat că semnalizarea insulinei este afectată în creierul pacienților cu AD [ 157 , 158] .]. Utilizarea glucozei cerebrale și fluxul sanguin sunt semnificativ diminuate deja în stadiile incipiente ale AD, în etapele ulterioare anomaliile metabolice și fiziologice se agravează – cu reduceri de 55-65% ale fluxului sanguin cerebral [159 ] . Astfel, metabolismul cerebral alterat al glucozei (similar cu modificările găsite în diabetul de tip 2) apare la scurt timp după apariția simptomelor legate de demență [ 160 ]. Mai multe studii și dovezi epidemiologice au legat rezistența periferică la insulină la subiecții sănătoși și, respectiv, la pacienții cu diabet, cu performanța cognitivă afectată [ 161 , 162 , 163] .]. Rezistența la insulină este propusă pentru a exacerba acumularea Ap, hiperfosforilarea tau și metabolismul energetic dezechilibrat. În plus, funcția hipocampului este alterată și inflamația este îmbunătățită (vezi recenzia de La Monte [ 164 ]). Constatarea că transferul microbiotei de la donatorii slabi sănătoși la pacienții cu rezistență la insulină îmbunătățește sensibilitatea la insulină a primitorilor și numărul de bacterii producătoare de butirat indică faptul că microbiota intestinală este implicată în mod crucial în semnalizarea insulinei [165] și dezvoltarea obezității . Mai mult, studiile pe animale, în care scaunul perechilor de gemeni obezi sau slabi umani a fost transferat la șoareci, au arătat rezultate similare: transferul de microbiom de la geamănul obez a dus la un metabolism afectat al glucozei la șoarecii primitori [166] .]. În același studiu, au fost investigate și efectele adăpostării în comun a șoarecilor transplantați cu microbiotă fecală obeză și slabă, ceea ce a arătat că a avut loc invazia unor membri specifici ai Bacteroidetes din microbiota slabă în microbiota obeză și a fost dependentă de dietă [ 166 ].
Experimentele cu șoareci hrăniți cu o dietă bogată în grăsimi au arătat că Lactobacillus intestinalis a fost singura specie a cărei abundență a fost corelată negativ cu modificările greutății corporale și ale masei de grăsime [ 167 ]. Dieta bogată în grăsimi a favorizat speciile bacteriene producătoare de propionat/acetat de SCFA, a căror abundență a fost strâns corelată cu adipozitatea [ 167 ]. Nivelurile ridicate de acetat și valerat SCFA s-au dovedit recent a fi asociate cu niveluri ridicate de LPS din sânge și citokine proinflamatorii, precum și cu o absorbție crescută de amiloid la pacienții cu demență [69 ] . Interesant este că dietele bogate în grăsimi pot afecta, de asemenea, neurogeneza adulților (vezi recenzia lui Zainuddin și Thuret [ 168 ).]) și poate afecta, de asemenea, negativ creierul prin modificarea formării adiponectinei: nivelurile scăzute ale adiponectinei serice s-au dovedit a fi legate de o funcție cognitivă mai proastă și de AD mai avansată [ 169 ].
Adiponectina produsă de țesutul adipos nu numai că crește absorbția de glucoză de către adipocite și monocite, dar are și efecte asupra sensibilității la insulină și reduce glucogeneza hepatică [ 169 ]. Mai mult, oxidarea acizilor grași liberi din mușchi este influențată în mod crucial de adiponectină, care previne și creșterea acizilor grași liberi și a trigliceridelor serice și are efecte antiinflamatorii și anti-aterogene [169 ] .
7. Dieta mediteraneană: Efecte asupra microbiotei intestinale
Cu toate acestea, dieta poate exercita și efecte pozitive asupra compoziției și inflamației microbiotei [ 170 ]. O dietă mediteraneană (DM) cu un consum mare de fructe și legume, un consum moderat de păsări de curte, pește, ouă și lactate și un consum redus de carne roșie și alimente procesate, pare să protejeze împotriva inflamației cronice și a bolilor asociate și este eficientă pentru a reduce risc pentru evenimente cardiovasculare [ 171 ].
The “PREDIMED” study [170], a prospective study investigating effects of a MD regarding cardiovascular outcome, showed that increased consumption of plant-based nutrients, such as vegetables and polysaccharides, coincided with higher counts of Bifidobacteria and higher total SCFA concentrations. Adherence to the MD was reflected by significantly higher levels of total SCFA. A higher ratio of Firmicutes–Bacteroidetes on the other hand was related to a lower adherence to the MD, while a higher abundance of Bacteroidetes was associated with lower animal protein intake. A high consumption of animal protein, saturated fats, and sugars (which is characteristic for a standard WD) affected gut microbiota diversity negatively. In normal-weight individuals, Christensenellaceae species were found more frequently compared to overweight participants. The authors concluded that diet and specific dietary components could affect microbiota composition, diversity, and activity (see also Figure 1), which may also affect host metabolism, e.g., by increasing the risk of Western diseases [170].
Adherence to the MD has been proposed as one of the top five modifiable factors against AD and cognitive decline, as several studies and also a meta-analysis have shown that high adherence to the MD is associated with a slower rate of cognitive decline [172,173,174,175]: compared with a low-fat diet, a dietary intervention with MD enhanced with either extra virgin olive oil or nuts appeared to improve cognition in middle-aged adults [176]. Furthermore, Mini Mental State examination (MMSE) and clock drawing test scores were higher for participants with MD versus a low-fat diet [176]. Therefore, two recent reviews also conclude that there are sufficient data to recommend the Mediterranean and Mediterranean-DASH (dietary approach to stop hypertension) Intervention for Neurodegenerative Delay (MIND) diets to delay the onset of AD [175,177].
In fact, recent data suggest that effects of MD are mainly due to positive effects on the gut microbiota: in the NU-AGE project dietary intervention with 1-year MD was effective to alter the gut microbiota, reduce frailty and improve cognitive function [178]. Data of 612 non-frail subjects across five European countries indicated that adherence to the MD diet provides a good ambience for bacteria, that produce butyrate, while gut microbiota that produce secondary bile acids, p-cresols, ethanol and carbon dioxide are rather diminished. Interestingly, the “beneficial, diet positive” gut microbiota taxa (Faecalibacterium prausnitzii, Roseburia (R. hominis), Eubacterium (E. rectale, E. eligens, E. xylanophilum), Bacteroides thetaiotaomicron, Prevotella copri and Anaerostipes hadrus) were positively associated with several markers of lower frailty and improved cognitive function, and negatively associated with inflammatory markers including CRP and IL-17 [178]. Increasing adherence to the MD increased the diversity of microbiota after one year of intervention significantly, there was a clear association between the microbiome and adherence to the MD. These changes in the intervention group were primarily driven by an increase in the intake of fibers, vitamins (C, B6, B9, thiamine) and minerals (Cu, K, Fe, Mn, Mg), while changes in the controls were associated with an increase in fat intake (saturated fats and mono-unsaturated fatty acids) relative to the MD intervention group. Interestingly, results of this study also demonstrated that there are country-specific patterns in dietary habits and that they are reflected by microbiome profiles. However, diet-responsive taxa identified across the entire cohort were largely shared across the different nationalities (Italian, UK/France, Netherlands/Poland) [178].
Not only the composition of nutrition, but also the quantity and the time period of nutrient intake appear to play an important role: positive effects of caloric restriction on the memory of rodents and humans have been demonstrated in intervention trials (see review by Zainuddin and Thuret [168]) and intermittent fasting has been proposed as promising approach for promoting brain health and preventing neurodegenerative disease in a recent review [179]. Intermittent fasting could exert beneficial effects by reducing insulin resistance, improving metabolic regulation, increasing autophagy, reducing inflammation and neuroinflammation, and increasing BDNF [179,180].
8. Systemic Inflammation, Nutrients and Neuroinflammation
Systemic immune activation appears to contribute significantly to the pathogenesis of AD outside the brain [17,100]: increased serum concentrations of inflammation markers like neopterin, sTNF-R75 and sIL2-R have been documented in patients with AD [181,182,183]. Neopterin is produced by macrophages upon stimulation with the proinflammatory cytokine interferon gamma (IFN-γ) in the elderly and to a greater extent in AD [184,185]. Neopterin was also documented to be elevated in patients with mild cognitive impairment [186], suggesting that systemic immune activation is already enhanced in the beginning of AD.
În paralel, citokina T-helper tip 1 (Th1) IFN-γ activează și enzima indoleamină 2,3-dioxigenaza-1 (IDO-1), care catalizează descompunerea triptofanului prin calea chinureninei [ 182 , 184 , 187 ] . S-a demonstrat că degradarea triptofanului este accelerată în AD [ 182 ]. Un studiu a demonstrat, de asemenea, că descompunerea triptofanului mediată imun, formarea neopterinei și peroxidarea lipidelor coincid la pacienții cu AD și sunt corelate între ele [ 188 ]. Peroxidarea lipidică îmbunătățită se datorează în principal stresului oxidativ, care este o consecință importantă a activării imune cronice [ 182] .]. S-a propus ca neopterina să fie un bun marker pentru stresul oxidativ și, de fapt, concentrațiile mari de neopterina s-au dovedit a fi asociate negativ cu concentrațiile de vitamine B acid folic și vitamina B12, care sunt oxidate ușor și ireversibil [182 ] . În plus, tetrahidrobiopterina, care este un cofactor important pentru formarea catecolaminelor și pentru metabolismul oxidului de azot, este ușor oxidată de radicalii liberi, sugerând că modificările axei dopaminei și adrenalinei, precum și scăderea reactivității vaselor sunt legate de stresul oxidativ (vezi, de asemenea,Figura 3).
Ciclul de metilare, ciclul folaților, stresul oxidativ și sintetizarea neurotransmițătorilor și depinde de nivelul vitaminei B. Abrevieri: NMDA = N -metil-D-aspartat; AD = boala Alzheimer; SAM = S-adenosil-metionină; BH4 = Tetrahidrobiopterina; CoA = coenzima A.
Disponibilitatea scăzută a vitaminelor B acid folic, vitamina B12, vitamina B3 și, de asemenea, vitamina B6 — probabil, de asemenea, produsă insuficient de microbiota intestinală [ 189 ] — afectează funcția multor căi biochimice importante: deoarece vitaminele B sunt cofactori esențiali pentru formarea neurotransmițătorilor serotoninei , dar și melatonina și vitamina B3 (niacină), conversia substraturilor în produsele finite poate fi afectată de disponibilitatea scăzută a vitaminei B (veziFigura 3). De asemenea, producția de catecolamine cum ar fi, de exemplu, adrenalina este dependentă de un aport suficient de vitamina B6 (pentru conversia tirozinei în L-DOPA), precum și de vitamina C, magneziu, fier și pteridină Tetrahidrobiopterina (THB).
Dacă acidul folic și vitaminele B6 și B12 sunt diminuate, crește și concentrațiile de homocisteină [ 190 ]. Homocisteina este toxică pentru celulele creierului, mai multe studii la animale ar putea arăta că homocisteina poate crește neurodegenerarea [ 191 ]. S-a demonstrat că deja mai devreme concentrații crescute de homocisteină coincid cu niveluri scăzute de vitamina B și concentrații crescute de neopterina marker de activare imună de tip Th1 [ 192 ] în sângele pacienților cu AD. Interesant, s-a demonstrat că suplimentarea pacienților obezi cu acid nicotinic de tip întârziat (vitamina B3) s-a dovedit că îmbunătățește biomarkerii pentru inflamație și sensibilitatea sistemică la insulină, împreună cu o creștere a abundenței Bacteroidetes [ 193 ] .]: s-a demonstrat că atât inflamația, cât și rezistența la insulină contribuie în mod important la neurodegenerarea în AD [ 194 ].
O declarație de consens internațional a clasificat concentrațiile plasmatice crescute de homocisteină totală drept factor de risc modificabil pentru dezvoltarea declinului cognitiv, a demenței și a bolii Alzheimer la persoanele în vârstă [190 ] . Mai multe studii prospective revizuite în această declarație de consens ar putea demonstra o relație de gradient între creșterea nivelului de homocisteină și afectarea cognitivă sau demența [ 195 , 196 , 197 , 198 ]. Deși se pune întrebarea dacă homocisteina în sine este factorul de risc pentru declinul cognitiv sau dacă este doar un marker pentru deficiența vitaminelor acid folic, vitamina B6 și B12 [199] .] rămâne de răspuns, măsurarea concentrațiilor de homocisteină pare a fi o metodă ieftină și simplă de a determina un factor de risc semnificativ pentru declinul cognitiv. Tratamentul cu vitamine B nu este eficient doar pentru a scădea homocisteina, ci și pentru a încetini rata atrofiei creierului întreg și regional și a declinului cognitiv semnificativ (așa cum a fost revizuit de Smith și colab. [190] ) . Prin urmare, declarația de consens a sfătuit, de asemenea, să se trateze homocisteina totală plasmatică moderat crescută (>11 μmol/L) la vârstnici cu vitamine B, care au fost considerate ca opțiune de tratament ușor, ieftin și sigur de tratat.
Cu toate acestea, nu numai scăderea disponibilității vitaminei B ar putea afecta metabolismul uman, ci și degradarea îmbunătățită a triptofanului mediată imun și epuizarea altor nutrienți importanți: o aprovizionare adecvată cu triptofan, de exemplu, este esențială pentru biosinteza proteinelor și, de asemenea, pentru reglarea în jos a cascadelor de activare imună. 200 ]. La pacienții cu boli cardiovasculare concentrațiile crescute de neopterina au fost asociate cu concentrații mai mici de triptofan [ 201 ], concentrații mai mari de homocisteină și concentrații reduse de vitamina B [ 202 ] și o supraviețuire mai proastă [ 203 , 204 ]. De asemenea, sa demonstrat că concentrațiile crescute de neopterina coincid cu concentrațiile scăzute de vitamina C, E [ 205] și vitamina D [ 206 ] indicând faptul că la pacienții cu boli cardiovasculare activarea/inflamația imună cronică este asociată cu scăderea concentrațiilor de vitamine, care se datorează probabil unui consum crescut de vitamine.
În mod similar, și la pacienții cu deficit de nutrienți și vitamine AD este bine documentat: într-o meta-analiză de Wilde și colaboratorii [ 207 ] descriu o disponibilitate mai scăzută a acidului docosahexaenoic (DHA), colină, vitamina B12, acid folic și LCR/creier în sânge și în LCR, vitamina C și vitamina E. În plus, concentrațiile de vitamina D sunt adesea scăzute la pacienții cu AD și au fost legate de riscul de declin cognitiv și demență la adulții în vârstă [ 208 , 209 ]. Deoarece creierul este dependent în mod critic de furnizarea de nutrienți, a fost sugerată o intervenție nutrițională cu mai multe componente [ 207 ]. O revizuire recentă a sugerat că, pe lângă starea nutrițională a pacienților, nivelurile de aminoacizi circulanți modificate la pacienții cu AD ar putea fi, de asemenea, din cauza depunerii de Aβ în sine [ 210] .]. Aceste modificări ar putea duce la dificultăți în formarea energiei musculare cu un consum crescut de acid aspartic, hipercatabolism muscular și un consum crescut de sisteme antioxidante [ 210 ].
9. Alți factori de risc importanți pentru dezvoltarea AD: activitatea fizică, tulburările de somn, stresul cronic și toxinele de mediu
Cu toate acestea, studii recente arată că și stilul de viață pare să joace un rol subestimat în dezvoltarea AD și poate, de asemenea, atenua factorii de risc genetici: un studiu recent care investighează rolul unui stil de viață sănătos și al riscului genetic la aproape 200.000 de pacienți fără tulburări cognitive la nivelul vârsta de 60 de ani ar putea arăta că atât un stil de viață nefavorabil, cât și un risc genetic ridicat au fost legate de un risc mai mare de a face demență [ 211 ]. Pe de altă parte, a fost asociat un stil de viață favorabil (care a fost definit ca interzicerea fumatului, activitate fizică regulată de cel puțin 75 de minute viguroase sau 150 de minute activitate moderată/săptămână, o dietă urmând recomandări privind prioritățile dietetice pentru sănătatea cardiometabolică și un consum redus de alcool). cu un risc mai mic de demență – și la pacienții cu risc genetic ridicat [ 211]. Efectele diferiților factori de stil de viață și, de asemenea, ale toxinelor din mediu asupra cogniției și microbiotei sunt analizate în următoarele subsecțiuni.
9.1. Activitate fizica
O revizuire realizată de Stephen și colegii [ 212 ] a investigat relația dintre activitatea fizică și incidentul AD: în mod foarte interesant, activitatea fizică a fost invers asociată cu riscul de AD în 18 studii, iar activitatea fizică în timpul liber a fost superioară celei legate de muncă. activitate fizica. Deja mai devreme, sa demonstrat că activitatea fizică moderată, dar prelungită (antrenamentul de fitness aerobic) crește volumul creierului la oamenii în vârstă [ 213 ], în timp ce activitatea fizică scăzută la mijlocul vârstei și, de asemenea, la vârstnici a fost asociată cu un risc mai mare de demență și AD [ 214 ].]. În plus, rezultatele unui studiu coreean prospectiv publicat recent, cu mai mult de 247.149 de indivizi urmăriți timp de 6 ani, confirmă faptul că activitatea fizică regulată continuă la pacienții cu deficiențe cognitive minore este asociată cu un efect protector împotriva dezvoltării demenței/AD [215], mediat de o creștere mai mare . Diversitatea microbiotei intestinale datorită stilului de viață activ [ 216 ].
9.2. Tulburări de somn
În plus, tulburările de somn par să contribuie în mod important la dezvoltarea AD: o meta-analiză realizată de Bubu și colegii [ 217 ] a arătat că persoanele cu probleme de somn au un risc de 1,68 ori mai mare pentru rezultatul combinat al tulburării cognitive și/sau AD. decât cei fără tulburări de somn. De fapt, aproximativ 15% din cazurile de AD la această populație au fost chiar atribuite problemelor de somn [ 217] .]—ceea ce este alarmant, deoarece astăzi mulți pacienți raportează probleme de somn. Privarea de somn este foarte critică pentru sănătate din mai multe motive: nu numai că modifică ritmurile circadiene, ci se asociază și cu o activitate crescută a sistemului nervos simpatic și a axei hipotalamo-hipofizo-suprarenale (axa HPA), exercită efecte metabolice importante. (de exemplu, crește riscul de obezitate [ 218 ]) și, de asemenea, induce răspunsuri proinflamatorii [ 219 ]. Întreruperea somnului pe termen scurt este asociată cu o sensibilitate crescută la stres, durere somatică, calitate redusă a vieții, suferință emoțională și tulburări de dispoziție și deficite cognitive, de memorie și de performanță la adulții altfel sănătoși [219 ]]. Experimentele arată că privarea de somn (față de valoarea inițială) are ca rezultat o creștere semnificativă a sarcinii Aβ în hipocampul și talamusul drept și că aceste creșteri sunt asociate cu înrăutățirea dispoziției [ 220 ].
Poor regeneration during sleep can impair learning and memory significantly, as it decreases neuronal plasticity and also neurogenesis [221]. It has been shown that functional neurons can be generated from adult neural precursors throughout life in the hippocampus and other regions of the brain [222]. The hippocampus appears to be especially vulnerable/sensitive to chronically restricted and disrupted sleep [223]. Thus, impaired hippocampal plasticity and reduced hippocampal cell proliferation (neurogenesis) may contribute importantly to cognitive disorders and psychiatric diseases [221,224]. During sleep, hormones like insulin-like growth factor 1 (IGF-1), growth hormone (GH), melatonin as well as BDNF, are upregulated and can promote hippocampal neurogenesis [225]. Cortisol, which in high concentrations inhibits neurogenesis in the hippocampus, is downregulated during normal sleep. This fact may also explain why prolonged sleep deprivation—as caused by shift working—superinduces the sympathetic system and impairs cognitive function [225]. Additionally, the composition, diversity and metabolic function of the gut microbiota are significantly altered in patients with insomnia compared to healthy individuals [226]. Eficiența crescută a somnului și timpul total de somn sunt corelate pozitiv cu diversitatea microbiomului, în timp ce trezirea după debutul somnului a fost corelată negativ cu diversitatea [ 227 ]. Eficiența somnului, concentrațiile de IL-6 și gândirea abstractă au fost mai bune la pacienții cu un număr mare de Bacteroidetes și Firmicutes , în timp ce alți taxoni ( Lachnospiraceae, Corynebacterium și Blautia ) au fost corelați negativ cu măsurile de somn [ 227 ].
9.3. Stresul cronic
În plus, supraactivarea persistentă a axei HPA de către stresul cronic pare să fie implicată în mod crucial în declinul cognitiv [ 228 ], posibil și prin modularea microbiotei: s-a demonstrat că stresul cronic scade numărul de Lactobacili [ 229 ] .
O dereglare precoce a axei HPA – care interacționează strâns cu microbiota intestinală – a fost bine documentată la pacienții cu AD și duce la o suprasecreție de cortizol. Glucocorticoizii intră cu ușurință în creier și afectează negativ hipocampul și, de asemenea, cortexul prefrontal, susținând astfel declinul cognitiv care apare în AD [ 228 ]. Stresul cronic este implicat critic în dezvoltarea și progresia bolii neurodegenerative [ 230 ]: prin dereglarea axei HPA, stresul poate accelera depunerea plăcii Aβ extracelulare și hiperfosforilarea intracelulară a tau [ 231 , 232 , 233 ] în modelele de șoarece pentru AD. În plus, poate declanșa, de asemenea, depresie comorbidă în bolile neurodegenerative.234 , 235 ]. S-a demonstrat că nivelurile crescute cronice de corticosteroizi modifică morfologia neuronilor și reduc volumul hipocampului la șobolani [ 236 , 237 ]. Nivelurile crescute de cortizol sunt găsite la pacienții cu AD în stadiu incipient și au fost legate de o progresie mai rapidă a AD [ 238 ]. Nivelurile plasmatice ridicate de cortizol au fost chiar identificate ca biomarker de încredere pentru AD [ 239 ]. Nivelurile inițiale mai ridicate de cortizol din LCR la pacienții cu AD au fost asociate cu o agravare clinică mai rapidă și declin cognitiv [ 240 ]. Nivelurile ridicate de cortizol consecutive stresului acut de mare intensitate s-au dovedit, de asemenea, a veni împreună cu memoria de lucru și memoria vizio-spațială afectate [ 241] .]. Disfuncția HPA a fost, de asemenea, legată de o permeabilitate intestinală crescută, motilitate și producție de mucus [ 242 ], ceea ce ar indica faptul că și microbiota intestinală ar putea fi afectată. De fapt, datele modelelor animale indică, de asemenea, că sunt implicate modificări ale microbiotei: la șoarecii expuși la stres psihosocial cronic, întreruperile funcției neurologice au fost asociate cu schimbări complexe ale microbiotei și răspunsuri imunoreglatoare modificate [243 ] . Stresul cronic a scăzut diversitatea generală a microbiomului cu niveluri semnificativ mai scăzute de Coriobacteriaceae și niveluri tendențial mai scăzute de Akkermansia muciniphilia [ 243 ].
În mod interesant, administrarea de lactobacillus helveticus NS8 a îmbunătățit disfuncția comportamentală (anxietate și depresie) și cognitivă indusă de reținere cronică la un model de șobolan, arătând un efect similar (sau chiar mai bun) cu citalopramul [244 ] . În plus, tratamentul cu Lactobacillus helveticus NS8 a fost, de asemenea, împreună cu niveluri mai scăzute de corticosteron plasmatic și hormoni adrenocorticotropi, niveluri mai mari de interleukină-10 în plasmă, niveluri restabilite de serotonină și norepinefrină (NE) din hipocamp și expresie mai mare a ARNm BDNF din hipocamp decât la șobolanii cu stres cronic. [ 244 ].
Supraactivarea persistentă a axei HPA afectează, de asemenea, neurogeneza și contracția hipocampului adult. Prin urmare, s-a propus ca AD să fie o boală progresivă care duce la deteriorarea ireversibilă a hipocampului, iar mai târziu, de asemenea, zonele creierului din cortex [ 245 ]. În timp ce studiile anterioare au propus că neurogeneza este posibilă numai în timpul fazei embrionare și perinatale, acum este bine stabilit că neuronii funcționali pot fi generați din precursori neuronali adulți de-a lungul vieții în hipocamp și în alte regiuni ale creierului [222 ] . Astfel, îmbătrânirea singură nu pare a fi cauzală pentru dezvoltarea AD [ 246 ], ci mai degrabă o facilitează prin impunerea proceselor, care duc în cele din urmă la neuroinflamație și neurodegenerare (vezi recenzia lui Nehls [2]).246 ]).
De fapt, studiile pe animale și pe oameni ar putea demonstra că diverși factori de stres pot afecta negativ compoziția microbiotei [ 247 , 248 , 249 ], cu, de exemplu, o creștere a Clostridiales și o scădere a Lactobacililor [ 250 ]. Gubert și colegii de muncă au analizat recent foarte cuprinzător efectele stresului, dietei și exercițiilor fizice ca modulatori ai microbiotei intestinale și au ajuns la concluzia că „modularea de mediu” a microbiotei intestinale ar trebui investigată prin studii ulterioare, deoarece poate oferi noi abordări terapeutice pentru neurodegenerative și tulburări psihice [ 251 ].
9.4. Toxine de mediu
Environmental toxins may in fact represent another, so far completely underestimated problem afflicting and troubling our gut microbiota and also our brain. Smoking, which has long been recognized as dangerous due to the inhalation of various toxins, can alter gut microbiota profiles by increasing Bacteroides-Prevotella species [252]. However, also effects of other environmental toxins on gut microbiota and on mental health should be investigated in more detail: recent reviews depict that neurotoxic metals such as lead, mercury, aluminum, cadmium and arsenic, as well as some pesticides and metal-based nanoparticles (NP) are involved in AD pathogenesis, as they are able to increase Aβ peptide and the phosphorylation of Tau protein (P-Tau), causing amyloid plaques and neurofibrillary tangles characteristic of AD [253,254].
O revizuire recentă descrie modificarea compoziției microbiotei intestinale și a profilurilor metabolice ca o consecință a expunerii la metale grele și, de asemenea, afirmă că perturbările microbiotei intestinale de către metalele grele pot provoca boli metabolice [255 ] . Microbiota intestinală poate afecta absorbția și metabolismul metalelor grele, dar expunerea cronică induce disbioza intestinală, care poate fi contracarată de probiotice [ 255 ].
În experimentele șoareci și aviare, expunerea cronică la glifosat a afectat negativ microbiota intestinală, în special la o vârstă mai fragedă [ 256 , 257 ]. Tratamentul cronic cu glifosat a indus o creștere a comportamentelor de anxietate și depresie și a modificat compoziția microbiotei intestinale în ceea ce privește abundența relativă și diversitatea filogenă în mod semnificativ la șoareci: Corynebacteria, Firmicutes, Bacteroidetes și Lactobacilli au fost diminuate la șoarecii tratați [ 256 ]. Expunerea pre- și neonatală la glifosat, care are și proprietăți antibiotice, a afectat fertilitatea șoarecilor și a dus la modificări ale comportamentului matern la mamele expuse [ 256] .]. Cu toate acestea, efectele asupra descendenților au fost și mai alarmante: glifosatul a suprimat microbii potențial benefici la o vârstă fragedă, iar animalele expuse la glifosat au prezentat o scădere consecutivă a activității locomotorii, sociabilității, învățării și memoriei pe termen scurt și lung ca șoarecii adulți, precum și modificări ale sistemelor lor colinergice și dopaminergice [ 256 ]. Microglia și astrocitele activate au fost observate la descendenții expuși la glifosat, iar neuroinflamația a fost demonstrată în cortexul prefrontal medial și hipocampus, mergând împreună cu anomalii cognitive [ 256 ]. O revizuire recentă discută efectele diferiților poluanți de mediu și mecanismele moleculare implicate în patogeneza tulburărilor neurologice [ 258 ].
Deja mai devreme s-a recunoscut că metabolismul dependent de bacterii al poluanților modulează toxicitatea toxinelor de mediu pentru gazdă [ 259 ], deoarece microbii intestinali au o capacitate mare de a metaboliza substanțele chimice din mediu. Pe de altă parte, contaminanții de mediu precum nanoparticulele de argint utilizate în industria alimentară sunt capabili să modifice puternic compoziția și/sau activitatea metabolică a bacteriilor gastrointestinale [ 259 ].
Pentru a rezuma, există un număr tot mai mare de „stresori de mediu”, care pot pune în pericol integritatea ecosistemului nostru uman/microbiotă. De fapt, pare esențial să realizăm că stilul nostru de viață influențează semnificativ și microbiota intestinală: disbioza microbiotei intestinale poate induce răspuns imunitar sistemic și neuroinflamație, dar, de asemenea, poate modifica metabolismul în mod important (de exemplu, prin inducerea rezistenței la insulină). Aceste modificări pot fi compensate de organism timp de mulți ani, dar în timpul îmbătrânirii capacitățile de a compensa scăderea și apar daune ireversibile – în cele din urmă merg împreună cu declinul cognitiv progresiv.
AD a fost privită ca o boală care derivă numai din SNC fără influența periferică timp de multe decenii. Mecanismele fiziopatologice de bază sunt încă descrise ca acumulare a proteinelor anormale Aß și tau hiperfosorilat [ 3 ]. Cu toate acestea, rapoarte mai recente sugerează un rol important al infecțiilor periferice cu bacterii orale și intestinale care induc cascade inflamatorii sistemice care duc apoi la leziuni neurologice prin declanșarea neuroinflamației [260 ] . Microbiota controlează aspectele de bază ale imunității și comportamentului în sănătate și boală [ 261]: o microbiotă intestinală bine echilibrată și diversă este necesară pentru reactivitatea, sociabilitatea și cogniția normale la stres. În plus, menține homeostazia SNC prin reglarea permeabilității intestinale, a funcției imunitare și a integrității BBB. Sinteza neurotransmițătorilor, sistemele de semnalizare sinaptică și neurotrofică și neurogeneza sunt influențate în mod crucial de microbiota intestinală.
Microbiomul intestinal disbiotic produce un amestec multi-component de produse metabolice microbiene, care activează semnificativ sistemul imunitar și cresc producția de citokine și mediatori inflamatori [ 41 ]. Acești compuși scad integritatea barierei mucoasei intestinale și a BBB, intensifică reacțiile inflamatorii sistemice și induc agregarea amiloidului. Scăderea etanșeității mucoasei intestinale și creșterea permeabilității BBB prin „compoziția greșită” a microbiotei intestinale sau o diversitate redusă la vârstnici facilitează astfel intrarea unei cantități mari de amiloid bacterian și LPS în circulație și SNC, unde cascadele inflamatorii induc tulburări neuropatologice. cu o componentă amiloidogenă [ 41 ].
Mai mult, este probabil ca infecțiile precum CMV să fie un declanșator al proceselor neuro-inflamatorii din creier [ 262 ]. Infecțiile periferice cronice și microflora intestinală disbiotică pot duce la multe modificări patologice în diferite țesuturi, de asemenea la distanță de sursa infecției [ 9 , 103]]. Inițial, organismul este capabil să reziste acestor modificări. Cu toate acestea, în timpul îmbătrânirii și mediată de diferiți factori ai stilului de viață nesănătos (cum ar fi o dietă nesănătoasă, de exemplu, WD standard, scăderea activității fizice, stresul cronic, tulburările de somn sau toxinele din mediu), capacitatea de regenerare și capacitatea de a limita infecțiile scad, ducând la procese neurodegenerative și in sfarsit la dementa. Astfel, modificările patologice pot începe în și în afara creierului cu mulți ani înainte de apariția simptomelor clinice ale AD.
Determinarea mai detaliată a interacțiunilor gazdă-microbiotă va permite progrese în medicina personalizată și în dezvoltarea probioticelor care se potrivesc cu modele individualizate [ 263 , 264 , 265 ]. Strategiile viitoare vor crea medicamente care modifică boala, precum și probiotice cu prebiotice și suplimente alimentare [ 266 , 267 ] în stadiile presimptomatice ale AD înainte de dezvoltarea demenței deschise. Opțiunile posibile de tratament vor fi analizate în capitolul următor.
11. Opțiuni/Perspective terapeutice pentru a încetini declinul cognitiv
Până acum, opțiunile de tratament pentru AD s-au concentrat în primul rând pe medicamente unice care interferează cu neurotransmisia glutamatergică și colinergică [ 3] .]. Cu toate acestea, aceste medicamente se adresează doar căilor biochimice unice într-un stadiu avansat al bolii. Date recente care arată influența microbiotei intestinale indică faptul că noile strategii de tratament la pacienții cu deficiențe cognitive ușoare ar putea fi mai utile pentru a încetini sau chiar a inversa declinul cognitiv: intestinul oferă cea mai mare interfață fizică între mediu (inclusiv microbiom) și individ. . Sunt necesare investigații clinice mai detaliate ale comunicării microbiotă-intestin-creier și ale rolului tulpinilor bacteriene probiotice specifice. Identificarea microorganismelor gastrointestinale folosind metode metagenomice și metabolomice poate crește capacitatea de a examina interacțiuni mai subtile dintre gazdă și microbiom [ 268] .]. Studiile ulterioare pe animale ne vor permite să evaluăm mai bine rolul diferiților nutrienți, dar și al toxinelor/stresorilor de mediu asupra microbiotei intestinale, a sistemului imunitar și, de asemenea, a sistemului neuro-endocrin: aceste date ne vor permite să judecăm efectele asupra „uman- ecosistem de microbiotă”, care sunt necesare urgent. În plus, efectele intervențiilor dietetice, cum ar fi vitaminele/micronutrienții, precum și compușii vegetali secundari și probioticele asupra microbiotei și a stării de inflamație ar trebui investigate mai detaliat.
Studiile recente încep și ele să abordeze acest subiect interesant și promițător și vor fi revizuite în cele ce urmează. Nu în ultimul rând, vor fi descrise abordări de intervenție multi-domeniu care abordează diferiți factori de stil de viață (și genetici) și va fi propusă medicina personalizată ca o nouă opțiune de tratament interesantă.
11.1. Vitamine și micronutrienți
Deficiența de nutrienți este o problemă bine stabilită în AD [ 207 ]. Suplimentarea pacienților cu substanțe deficitare (țintind în mod ideal deficiențele documentate prin diagnosticul prealabil de laborator) pare rezonabilă. După cum sa menționat deja mai sus, vitaminele B sunt recomandate ca tratament ușor, ieftin și sigur de tratat pentru a trata homocisteina totală plasmatică moderat crescută (>11 μmol/L) la vârstnici, în special la pacienții cu deficite cognitive [269 ] . Vitaminele B nu sunt doar esențiale pentru buna funcționare a multor căi biochimice în gazdă, ci și pentru viabilitatea/metabolismul anumitor tulpini bacteriene auxotrofe ale microbiotei intestinale: partajarea vitaminei B a fost propusă pentru a promova stabilitatea microbiană intestinală. comunități [ 270]. Cu toate acestea, până acum există puține informații despre posibilele efecte ale suplimentelor cu vitamina B asupra microbiotei până acum. Din cunoștințele noastre, sunt disponibile doar datele care investighează efectele acidului nicotinic cu eliberare întârziată (Vitamina B3) la pacienții obezi: suplimentarea cu acid nicotinic cu eliberare întârziată a fost capabilă să crească abundența Bacteroidetes și să îmbunătățească biomarkerii pentru sensibilitatea sistemică la insulină și inflamația metabolică [ 193 ] . ].
O revizuire sistematică recentă a studiilor controlate randomizate care investighează dacă alte intervenții alimentare au avut efecte asupra cogniției pacienților cu AD a concluzionat că acidul gras omega-3 a arătat efecte pozitive la diferite doze [271 ] . Acest lucru ar putea fi parțial legat de efectele microbiomului intestinal, deoarece s-a demonstrat că acizii grași omega-3 influențează abundența Faecalibacteria, Bacteriodetes și Lachnospiraceae . Se sugerează că interacțiunea dintre microbiota intestinală și acizii grași omega-3 ajută la menținerea integrității peretelui intestinal și exercită efecte benefice asupra interacțiunii cu celulele imune ale gazdei [272] .]. În plus, un supliment oral cu o combinație de acid eicosapentaenoic (EPA), acid docosahexaenoic (DHA), fosfolipide, uridin monofosfat, colină, seleniu și vitaminele B6, B12, B9, C și E părea să fie eficient în stadiile incipiente. din AD.
Conform unei analize Cochrane din 2019, există, de asemenea, dovezi de calitate moderată dintr-un singur studiu că suplimentarea cu vitamina E poate încetini declinul funcțional în AD [ 273 ]. Un studiu pilot foarte interesant de 10 săptămâni a investigat efectele administrării de micronutrienți cu spectru larg asupra conținutului de microbiom fecal la pacienții tineri cu tulburare de deficit de atenție/hiperactivitate [274] : abundența diferențială și frecvența relativă a Actinobacteriilor au scăzut semnificativ tratamentul post-micronutrient. , iar diversitatea microbiomului a crescut [ 274 ].
11.2. Substanțe derivate din plante și polifenoli
Ginsengul, inozitolul și formulele nutriționale specializate par să aibă un efect pozitiv asupra cogniției [ 271 ]. De fapt, sa demonstrat că ginseng îmbunătățește creșterea microbiotei benefice, inclusiv Lactobacillus și Bacteroides [ 275 ]. În plus, s-a demonstrat că inozitol crește producția microbiană de SCFA și butirat și crește creșterea lactobacililor [ 276] .]. Deși calitatea acestor intervenții a fost scăzută până la moderată, studii mai randomizate și controlate cu ingrediente vegetale secundare, cum ar fi polifenolii dietetici, ar putea fi interesante, în special cu polifenoli și curcumină. Aceste substanțe pluripotente sunt în mare parte captatori de radicali puternici, modulează cascadele proinflamatorii (așa cum au fost revizuite de Dias și colab. [ 277 ] și Monje și colab. [ 151 ]) și pot chiar influența formarea amiloidului [ 278 , 279 ]. De fapt, un studiu pilot dublu-orb, controlat cu placebo, la oameni nedementați, ar putea demonstra că o formă biodisponibilă de curcumină a condus la beneficii semnificative de memorie și atenție [ 280] .]. În plus, scanările FDDNP-PET efectuate înainte și după 18 luni de tratament cu curcumină au sugerat că beneficiile comportamentale și cognitive au fost asociate cu scăderea acumulării de plăci și încurcături în regiunile creierului care modulează starea de spirit și memoria [280 ] . Aceste rezultate promițătoare par să merite testate în studii prospective mai mari [ 281 ].
Atât resveratrolul, cât și curcumina modulează microbiota intestinală și ar putea fi utile pentru tratamentul sindromului metabolic [ 282 , 283 ] și a afecțiunilor asociate precum AD într-un concept terapeutic combinat [ 284 ]. Tratamentul cu curcumină la om modifică considerabil raportul dintre bacteriile benefice și cele patogene prin creșterea abundenței de Bifidobacterii, Lactobacili și reducerea încărcăturii de Enterobacterie și Enterococi proinflamatorii [ 283 ]. Curcumina exercită diverse efecte neuroprotectoare (vezi recenzia lui Cole și colab. [ 285 ]) prin diferite căi, de exemplu, prin reducerea amiloidului și inhibarea toxicității amiloidului [ 285]]. În plus, curcumina este foarte eficientă pentru a îmbunătăți sinteza unui alt nutrient esențial pentru creier, și anume DHA [ 286 ].
Similarly, resveratrol has potent antioxidant, anti-inflammatory, anticarcinogenic, anti-aging and neuroprotective effects (which were reviewed recently by Arbo et al. [287]), but in clinical trials so far effects on cognition were not significant—possibly due to the low bioavailability of resveratrol [287]. Still, a higher dietary intake of flavonoids has been associated with a reduced risk of developing Alzheimer dementia [288,289,290]. Flavonoids could further change the composition of gut microbiota by increasing the growth of Bifidobacteria and Lactobacilli [291], especially by fermentation procedures of fruit or vegetables rich in antioxidant plant compounds, e.g., fermented papaya was shown to reduce oxidative stress in AD patients, thereby probably reducing neuronal damage [292]. Furthermore, mango was shown to positively affect intestinal microbiota by increasing the presence of Faecalibacterium, Bifidobacterium, Prevotella and Bacteriodes which are beneficial for human gut health [293]. A recent study further demonstrated that pomegranate stimulated the growth of Akkermansia munciniphila, which plays an important role in the breakdown of phenolic intestinal compounds and may thereby contribute to its prebiotic effects [294]. Positive effects of various single nutrients on AD are summarized in Table 1—combination supplements are discussed above.
Table 1
Positive effects of various nutrients on AD.
Nutrient
Positive Effect
Reference
omega-3 fatty acid
smaller decline in MMSE and ADAS-cog
Freund-Levi et al. [295] Quinn et al. [296] Shinto et al. [297]
MMSE = Mini-Mental State Exam; ADAS-cog = Alzheimer’s Disease Assessment Scale-Cognitive Subscale; MWM = Morris Water Maze test; AD = Alzheimer’s disease; FU = Follow-up.
11.3. Probiotics
The first study (double-blind, placebo-controlled) performed in patients with AD showed that supplementation with Lactobacillus acidophilus, Lactobacillus casei, Bifidobacterium bifidum, and Lactobacillus fermentum resulted in a significant improvement in MMSE score [304]. In addition, decreases in plasma concentrations of serum high-sensitivity C-reactive protein, malondialdehyde, insulin resistance, and serum triglycerides were encountered compared to the control group [304]. Another Iranian study examining probiotic and selenium co-supplementation showed a significant increase in MMSE score in patients with AD, in parallel also a significant reduction in hsCRP, lower insulin levels and a significant increase in total antioxidant capacity and glutathione levels was seen [305].
Very recent animal studies [306,307], clinical trials [308] and a meta-analysis of randomized controlled clinical trials [309] also suggest that altering microbiota with probiotics can lead to changes in brain function including cognitive functions. Mice in an early stage of AD were treated with a probiotic formulation, that decreased their cognitive decline compared with controls and reduced accumulation of amyloid β aggregates [310]. Furthermore, the probiotics affected the composition of gut microbiota, plasma concentrations of inflammatory cytokines and key metabolic hormones [310]. Furthermore, results of a recent Austrian study suggest that probiotics have beneficial effects on cognition and mood [311]: healthy students taking a multi-strain probiotic during a stressful period of exams performed better in recognition tasks than students who got placebo or without treatment (85% recognition compared to 70%). Furthermore, they were happier and functional brain MRI scans showed that brain regions for memory, motor function and attention were better perfused in students taking probiotics than in those with placebo or no treatment [311]. In emotional decision-making processes participants from the probiotic group showed increased alertness and were more confident and definite in making decisions. Interestingly, microbiome composition was related with scores for response accuracy, depressiveness and hopelessness.
In an earlier exploratory intervention study, supplementation of a multispecies probiotic for 4 weeks resulted in a significant increase of the anti-inflammatory fecal bacterial strain Faecalibacterium prausnitzii in patients with AD. In parallel zonulin, a leaky gut marker significantly decreased by this intervention [90], while inflammatory markers (neopterin, Kynurenine/tryptophan ratio) slightly increased.
Previous animal studies showed that administration of probiotics induced changes in genes involved in inflammation and neural plasticity with a positive impact on neural functions [312]. Mice treated with probiotics showed higher plasma levels of gut hormones ghrelin and leptin: ghrelin counteracts memory deficits and synaptic degeneration in AD animal models [313], and leptin acts as neurotrophic factor and exerts neuroprotective effects against toxicity induced by Aβ oligomers in vitro [314]. Oral intake of probiotics furthermore also showed beneficial consequences on mood and psychological distress in rats and healthy human volunteers [315]. As depression and chronic stress are frequently observed in patients with AD, and supplementation with Lactobacillus helveticus NS8 was effective to improve chronic stress-induced depression/anxiety and cognitive dysfunction also in a mouse model [244], probiotic supplementation might be beneficial to treat several symptoms of AD patients in parallel.
Still, despite these promising data, further and larger clinical trials are necessary to assess the efficacy of probiotics in AD patients more exactly. A study performed in patients with severe AD could not observe significant changes regarding cognition, inflammation markers (TNF-a, IL-6) and oxidative stress markers (malondiadehyde, glutathione, total antioxidant capacity) after 12 weeks of probiotic supplementation [316]. Probably, effects of probiotic supplementation depend on several variables: the composition of the probiotic supplement, the number of bacteria used, and of course also the severity of dementia or other co-morbidities of patients driving inflammation. Additionally, also the diet of patients, supplementation with vitamins, micronutrients, phytochemicals, and physical activity, as well as the load of environmental toxins and stress might influence the effects of probiotic supplementation.
Fecal microbiota transplantation (FMT) is increasingly applied as a therapeutic in conditions associated with gut dysbiosis. FMT of fecal microflora from healthy individuals to patients with AD could help restore intestinal microflora and reduce the negative impact of the dysbiotic microbiome on the gut and brain function, though human FMT studies in neurodegenerative diseases are still lacking [41,317,318].
However, as FMT is an invasive method, other less invasive methods should be tried first: in fact, dietary intervention strategies like adherence to a MD appear to be very effective to improve cognitive decline and enlarge the diversity of our gut microbiota (see also chapter 7).
11.4. Multi-Domain Intervention and Personalized Medicine
To modify unfavorable lifestyle factors, multi-domain intervention with changes of diet, increased physical activity, cognitive training and the monitoring of vascular risk factors appears as very promising therapeutic option: in the large, long-term, randomized controlled FINGER trial this therapeutic approach was effective to improve cognitive functioning in at-risk elderly people from the general population compared to a control group (general health advice) [319]. This multi-domain lifestyle intervention shall now also be tested by the The World-Wide FINGERS (WW-FINGERS) global network for dementia risk reduction and prevention, in over 25 countries [320].
However, as adherence decreases with increasing intervention complexity and intensity [321], personalized medicine approaches might be the treatment of choice: in fact, the neurologist Dale Bredesen, who is involved in AD research since many decades, has developed such a personalized treatment approach. He proposes that AD (like other chronic illnesses) is an age—associated network imbalance featuring many underlying mechanisms, and many or all of these mechanisms may need to be addressed therapeutically for optimal clinical efficacy [322]. Therefore, he has developed a novel, comprehensive, and personalized therapeutic program, which addresses underlying metabolic alterations and also overwhelming inflammatory cascades [323,324]. He suggests to first diagnose metabolic alterations, nutrient deficiencies and inflammation by various laboratory analyses/functional brain scans/cognitive tests. Results of these laboratory analyses and tests show, which processes (local and systemic inflammation, wrong diet [leading to dysybiosis, systemic inflammation, deficiency of essential nutrients, insulin resistance, neuroinflammation], chronic stress, sleep disturbances, toxins and hormonal imbalances) are dysregulated [322]. In response to these underlying causes patients are recommended to, e.g., modify their diet, do more sports, reduce stress by yoga or take supplements or hormones for diagnosed deficiencies. Additionally, also the adequate treatment of infections (e.g., of periodontitis) and the elimination of environmental toxins are scheduled in this program. This personalized treatment ensures that patients are treated with necessary and effective measures, so that they also adhere to this multi-domain therapy.
This “system biological approach” by Bredesen has already been shown to be effective in pilot studies: in a pilot study with 10 patients cognitive decline could be reversed in nine patients (Bredesen 2014). Furthermore, a follow-up study with results of 100 treated patients (treated by different physicians) could demonstrate that the concept works [325]. To our mind, this treatment approach for AD appears to be very promising and should be investigated in further clinical trials enrolling more patients. However, not only Bredesen, but also other scientists signify the importance of personalized medicine in AD patients. Peng and coworkers suggest that in-depth information (from genome sequencing, brain imaging, peripheral biomarkers, and even functional assays on neurons derived from patient-specific induced pluripotent cells) in combination with demographic information (APOE genotype, age, gender, education, environmental exposure, life style, and medical history) will enable a better understanding of the disease mechanisms underlying AD and facilitate the design of personalized stage specific therapeutic intervention in AD [326].
Times are changing and in the “age of chronic diseases” system biological treatment approaches using personalized medicine therapy should be examined for their efficacy by well conducted clinical trials to guarantee sustainable and effective medical care.
Conceptualization, F.L. and D.F.; methodology, F.L., K.K. and D.F.; validation, D.E., K.S. and L.L.; resources, D.F. and F.L.; data curation, F.L.; writing—original draft preparation, F.L. and K.K.; writing—review and editing, D.E., K.S., S.G., D.F., and L.L.; visualization, L.L. All authors have read and agreed to the published version of the manuscript.
The authors declare no conflict of interest. K. Kurz is currently performing a placebo-controlled double blind study investigating effects of probiotics in patients with recurrent infections and post-infectious fatigue, for which study medication is provided (without costs) by an Austrian firm producing probiotics.
6. Murphy M.P., LeVine H., 3rd Alzheimer’s disease and the amyloid-beta peptide. J. Alzheimers Dis. 2010;19:311–323. doi: 10.3233/JAD-2010-1221. [PMC free article] [PubMed] [CrossRef] [Google Scholar]7.
Aizenstein HJ, Nebes RD, Saxton JA, Price JC, Mathis CA, Tsopelas ND, Ziolko SK, James JA, Snitz BE, Houck PR și colab. Depozite frecvente de amiloid fără afectare cognitivă semnificativă la vârstnici. Arc. Neurol. 2008; 65 :1509–1517. doi: 10.1001/archneur.65.11.1509. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]8.
Fuster-Matanzo A., Llorens-Martín M., Hernández F., Avila J. Rolul neuroinflamației în neurogeneza adultului și boala Alzheimer: abordări terapeutice. Mediat. Inflamm. 2013; 2013 :260925. doi: 10.1155/2013/260925. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]9.
Abid MB, Koh CJ Probiotice în sănătate și boală: O păcăli pe Mama Natură? Infecţie. 2019; 47 :911–917. doi: 10.1007/s15010-019-01351-0. [ PubMed ] [ CrossRef ] [ Google Scholar ]10.
Johnson K. Compoziția și diversitatea microbiomului intestinal sunt legate de trăsăturile personalității umane. Zumzet. Microbiome J. 2020; 15 :100069. doi: 10.1016/j.humic.2019.100069. [ CrossRef ] [ Google Scholar ]11.
Ursell LK, Haiser HJ, Van Treuren W., Garg N., Reddivari L., Vanamala J., Dorrestein PC, Turnbaugh PJ, Knight R. Metabolomul intestinal: o intersecție între microbiotă și gazdă. Gastroenterologie. 2014; 146 :1470–1476. doi: 10.1053/j.gastro.2014.03.001. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]12.
Zheng D., Liwinski T., Elinav E. Interacțiunea dintre microbiotă și imunitate în sănătate și boală. Cell Res. 2020; 30 :492–506. doi: 10.1038/s41422-020-0332-7. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]13.
Thursby E., Juge N. Introducere în microbiota intestinală umană. Biochim. J. 2017; 474 :1823–1836. doi: 10.1042/BCJ20160510. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]15.
Sender R., Fuchs S., Milo R. Suntem cu adevărat depășiți numeric cu mult? Revizuirea raportului dintre bacterii și celule gazdă la oameni. Celulă. 2016; 164 :337–340. doi: 10.1016/j.cell.2016.01.013. [ PubMed ] [ CrossRef ] [ Google Scholar ]16.
Grice EA, Segre JA Microbiomul uman: al doilea genom al nostru. Annu. Rev. Genom. Zumzet. Genet. 2012; 13 :151–170. doi: 10.1146/annurev-genom-090711-163814. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]17.
Giau VV, Wu SY, Jamerlan A., An SSA, Kim SY, Hulme J. Gut Microbiota and Their Neuroinflammatory Implications in Alzheimer’s Disease. Nutrienți. 2018; 10 :1765. doi: 10.3390/nu10111765. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]18.
Venegas DP, De la Fuente MK, Landskron G., González MJ, Quera R., Dijkstra G., Harmsen HJM, Faber KN, Hermoso MA Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Imune Regulation and its relevance pentru boli inflamatorii intestinale. Față. Imunol. 2019; 10 doi: 10.3389/fimmu.2019.00277. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]19.
Al Bander Z., Nitert MD, Mousa A., Naderpoor N. Microbiota intestinală și inflamația: o privire de ansamblu. Int. J. Environ. Res. Sănătate Publică. 2020; 17 :7618. doi: 10.3390/ijerph17207618. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]20.
Wang B., Yao M., Lv L., Ling Z., Li L. Microbiota umană în sănătate și boală. Inginerie. 2017; 3 :71–82. doi: 10.1016/J.ENG.2017.01.008. [ CrossRef ] [ Google Scholar ]21.
Noble EE, Hsu TM, Kanoski SE Gut to Brain Dysbiosis: Mecanisms Linking Western Diet Consumption, the Microbiome, and Cognitive Impairment. Față. Comportament. Neurosci. 2017; 11 :9. doi: 10.3389/fnbeh.2017.00009. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]22.
Vogt NM, Kerby RL, Dill-McFarland KA, Harding SJ, Merluzzi AP, Johnson SC, Carlsson CM, Asthana S., Zetterberg H., Blennow K., et al. Alterări ale microbiomului intestinal în boala Alzheimer. Sci. Rep. 2017; 7 :13537. doi: 10.1038/s41598-017-13601-y. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]23.
Liu P., Wu L., Peng G., Han Y., Tang R., Ge J., Zhang L., Jia L., Yue S., Zhou K. și colab. Microbiomii alterați disting boala Alzheimer de afectarea cognitivă ușoară amnestică și sănătatea într-o cohortă chineză. Comportamentul creierului. Imun. 2019; 80 :633–643. doi: 10.1016/j.bbi.2019.05.008. [ PubMed ] [ CrossRef ] [ Google Scholar ]24.
Zhuang ZQ, Shen LL, Li WW, Fu X., Zeng F., Gui L., Lü Y., Cai M., Zhu C., Tan YL și colab. Microbiota intestinală este alterată la pacienții cu boala Alzheimer. J. Alzheimer Dis. 2018; 63 :1337–1346. doi: 10.3233/JAD-180176. [ PubMed ] [ CrossRef ] [ Google Scholar ]25.
Harach T., Marungruang N., Duthilleul N., Cheatham V., Mc Coy KD, Frisoni G., Neher JJ, Fåk F., Jucker M., Lasser T., et al. Reducerea patologiei amiloidului Abeta la șoarecii transgenici APPPS1 în absența microbiotei intestinale. Sci. Rep. 2017; 7 :41802. doi: 10.1038/srep41802. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]26.
Jiang C., Li G., Huang P., Liu Z., Zhao B. Microbiota intestinală și boala Alzheimer. J. Alzheimer Dis. 2017; 58 :1–15. doi: 10.3233/JAD-161141. [ PubMed ] [ CrossRef ] [ Google Scholar ]27.
Vighi G., Marcucci F., Sensi L., Di Cara G., Frati F. Allergy and the gastrointestinal system. Clin. Exp. Imunol. 2008; 153 (Supl. 1):3–6. doi: 10.1111/j.1365-2249.2008.03713.x. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]28.
Mittal R., Coopersmith CM Redefinirea intestinului ca motor al bolii critice. Trends Mol. Med. 2014; 20 :214–223. doi: 10.1016/j.molmed.2013.08.004. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]29.
Sampson TR, Mazmanian SK Controlul dezvoltării, funcției și comportamentului creierului de către microbiom. Microbul gazdă celulară. 2015; 17 :565–576. doi: 10.1016/j.chom.2015.04.011. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]30.
Westfall S., Lomis N., Kahouli I., Dia SY, Singh SP, Prakash S. Microbiome, probiotice și boli neurodegenerative: Descifrarea axei creierului intestinal. Cell Mol. Life Sci. 2017; 74 :3769–3787. doi: 10.1007/s00018-017-2550-9. [ PubMed ] [ CrossRef ] [ Google Scholar ]31.
De-Paula VDJR, Forlenza AS, Forlenza OV Relevanța microbiotei intestinale în cogniție, comportament și boala Alzheimer. Pharmacol. Res. 2018; 136 :29–34. doi: 10.1016/j.phrs.2018.07.007. [ PubMed ] [ CrossRef ] [ Google Scholar ]32.
Zhao Y., Dua P., Lukiw WJ Surse microbiene de amiloid și relevanță pentru amiloidogeneză și boala Alzheimer (AD) J. Alzheimers Dis. Parc. 2015; 5 :177. doi: 10.4172/2161-0460.1000177. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]33.
Cryan JF, Dinan TG Mind-altering microorganismes: The impact of the gut microbiota on creier and behavior. Nat. Pr. Neurosci. 2012; 13 :701–712. doi: 10.1038/nrn3346. [ PubMed ] [ CrossRef ] [ Google Scholar ]34.
Geirnaert A., Calatayud M., Grootaert C., Laukens D., Devriese S., Smagghe G., De Vos M., Boon N., Van de Wiele T. Bacterii producătoare de butirat suplimentate in vitro la boala Crohn microbiota pacientului a crescut producția de butirat și a îmbunătățit integritatea barierei epiteliale intestinale. Sci. Rep. 2017; 7 :11450. doi: 10.1038/s41598-017-11734-8. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]35.
Yang Q., Liang Q., Balakrishnan B., Belobrajdic DP, Feng Q.-J., Zhang W. Role of Dietary Nutrients in the Modulation of Gut Microbiota: A Narrative Review. Nutrienți. 2020; 12 :381. doi: 10.3390/nu12020381. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]36.
Kowalski K., Mulak A. Brain-Gut-Microbiota Axis in Alzheimer’s Disease. J. Neurogastroenterol. Motil. 2019; 25 :48–60. doi: 10.5056/jnm18087. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]38.
Bronzuoli MR, Iacomino A., Steardo L., Scuderi C. Targeting neuroinflammation in Alzheimer’s disease. J. Inflamm. Res. 2016; 9 :199–208. doi: 10.2147/JIR.S86958. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]39.
Liu X., Nemeth DP, Tarr AJ, Belevych N., Syed ZW, Wang Y., Ismail AS, Reed NS, Sheridan JF, Yajnik AR și colab. Euinflamația atenuează neuroinflamația indusă de inflamația periferică și atenuează semnalizarea imună la creier. Comportamentul creierului. Imun. 2016; 54 :140–148. doi: 10.1016/j.bbi.2016.01.018. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]40.
Khandelwal PJ, Herman AM, Moussa CEH Inflamația în stadiile incipiente ale patologiei neurodegenerative. J. Neuroimmunol. 2011; 238 :1–11. doi: 10.1016/j.jneuroim.2011.07.002. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]41.
Sochocka M., Donskow-Łysoniewska K., Diniz BS, Kurpas D., Brzozowska E., Leszek J. The Gut Microbiome Alterations and Inflammation-Driven Pathogenesis of Alzheimer’s Disease-A Critical Review. Mol. Neurobiol. 2019; 56 :1841–1851. doi: 10.1007/s12035-018-1188-4. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]42.
Sochocka M., Diniz BS, Leszek J. Răspuns inflamator în SNC: prieten sau dușman? Mol. Neurobiol. 2017; 54 :8071–8089. doi: 10.1007/s12035-016-0297-1. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]43.
Le Page A., Dupuis G., Frost EH, Larbi A., Pawelec G., Witkowski JM, Fulop T. Rolul sistemului imunitar înnăscut periferic în dezvoltarea bolii Alzheimer. Exp. Gerontol. 2018; 107 :59–66. doi: 10.1016/j.exger.2017.12.019. [ PubMed ] [ CrossRef ] [ Google Scholar ]44.
Barnes LL, Capuano AW, Aiello AE, Turner AD, Yolken RH, Torrey EF, Bennett DA Infecția cu citomegalovirus și riscul bolii Alzheimer la persoanele în vârstă alb-negru. J. Infectează. Dis. 2015; 211 :230–237. doi: 10.1093/infdis/jiu437. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]46.
Bonaz B., Bazin T., Pellissier S. The Vagus Nerve at the Interface of the Microbiota-Gut-Brain Axis. Față. Neurosci. 2018; 12 doi: 10.3389/fnins.2018.00049. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]47.
Leblhuber F., Geisler S., Steiner K., Fuchs D., Schütz B. Calprotectina fecală crescută la pacienții cu demență Alzheimer indică intestin permeabil. J. Neural Transm. (Viena) 2015; 122 :1319–1322. doi: 10.1007/s00702-015-1381-9. [ PubMed ] [ CrossRef ] [ Google Scholar ]48.
Wang C., Klechikov AG, Gharibyan AL, Wärmländer SK, Jarvet J., Zhao L., Jia X., Narayana VK, Shankar SK, Olofsson A. și colab. Rolul S100A9 proinflamator în cascada amiloid-neuroinflamatoare a bolii Alzheimer. Acta neuropathol. 2014; 127 :507–522. doi: 10.1007/s00401-013-1208-4. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]49.
Leblhuber F., Strasser B., Steiner K., Gostner J., Schuetz B., Fuchs D. Despre rolul microbiotei intestinale la pacientii cu declin cognitiv. J. Pharm. Pharmacol. 2017; 5 :648–653. [ Google Scholar ]50.
Leblhuber F., Egger M., Schuetz B., Fuchs D. Comentariu: Efectul suplimentării probiotice asupra funcției cognitive și a statusului metabolic în boala Alzheimer: un studiu randomizat, dublu-orb și controlat. Față. Îmbătrânirea Neurosci. 2018; 10 doi: 10.3389/fnagi.2018.00054. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]51.
Zhang C., Liu Y., Gilthorpe J., van der Maarel JRC MRP14 (S100A9) Proteina interacționează cu peptida beta-amiloid Alzheimer și induce fibrilizarea acesteia. Plus unu. 2012; 7 :e32953. doi: 10.1371/journal.pone.0032953. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]52.
Marizzoni M., Provasi S., Cattaneo A., Frisoni GB Microbiota și bolile neurodegenerative. Curr. Opinează. Neurol. 2017; 30 :630–638. doi: 10.1097/WCO.0000000000000496. [ PubMed ] [ CrossRef ] [ Google Scholar ]53.
Spielman LJ, Gibson DL, Klegeris A. Intestul nesănătos, creierul nesănătos: Rolul microbiotei intestinale în bolile neurodegenerative. Neurochema. Int. 2018; 120 :149–163. doi: 10.1016/j.neuint.2018.08.005. [ PubMed ] [ CrossRef ] [ Google Scholar ]54.
Gareau MG, Wine E., Rodrigues DM, Cho JH, Whary MT, Philpott DJ, Macqueen G., Sherman PM Infecția bacteriană provoacă disfuncție de memorie indusă de stres la șoareci. Intestin. 2011; 60 :307–317. doi: 10.1136/gut.2009.202515. [ PubMed ] [ CrossRef ] [ Google Scholar ]55.
Michalski B., Corrada MM, Kawas CH, Fahnestock M. Factorul neurotrofic derivat din creier și expresia TrkB în „cel mai vechi-vechi”, studiul 90+: corelația cu statusul cognitiv și nivelurile de beta-amiloid solubil. Neurobiol. Îmbătrânire. 2015; 36 :3130–3139. doi: 10.1016/j.neurobiolaging.2015.08.022. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]56.
Hill JM, Lukiw WJ Amiloizi generați de microbi și boala Alzheimer (AD) Front. Îmbătrânirea Neurosci. 2015; 7 :9. doi: 10.3389/fnagi.2015.00009. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]57.
Taylor JD, Matthews SJ O nouă perspectivă asupra controlului molecular al amiloizilor funcționali bacterieni. Față. Infectarea celulelor. Microbiol. 2015; 5:33 . doi: 10.3389/fcimb.2015.00033. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]58.
Romero D., Aguilar C., Losick R., Kolter R. Fibrele amiloide asigură integritate structurală biofilmelor de Bacillus subtilis. Proc. Natl. Acad. Sci. STATELE UNITE ALE AMERICII. 2010; 107 :2230–2234. doi: 10.1073/pnas.0910560107. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
60. Hammer N.D., Wang X., McGuffie B.A., Chapman M.R. Amyloids: Friend or foe? J. Alzheimers Dis. 2008;13:407–419. doi: 10.3233/JAD-2008-13406. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
61. Zhao Y., Lukiw W.J. Microbiome-generated amyloid and potential impact on amyloidogenesis in Alzheimer’s disease (AD) J. Nat. Sci. 2015;1:e138. [PMC free article] [PubMed] [Google Scholar]
62. Gąsiorowski K., Brokos B., Echeverria V., Barreto G.E., Leszek J. RAGE-TLR Crosstalk Sustains Chronic Inflammation in Neurodegeneration. Mol. Neurobiol. 2018;55:1463–1476. doi: 10.1007/s12035-017-0419-4. [PubMed] [CrossRef] [Google Scholar]
63. Tan J., McKenzie C., Potamitis M., Thorburn A.N., Mackay C.R., Macia L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014;121:91–119. doi: 10.1016/b978-0-12-800100-4.00003-9. [PubMed] [CrossRef] [Google Scholar]
64. Osbelt L., Thiemann S., Smit N., Lesker T.R., Schröter M., Gálvez E.J.C., Schmidt-Hohagen K., Pils M.C., Mühlen S., Dersch P., et al. Variations in microbiota composition of laboratory mice influence Citrobacter rodentium infection via variable short-chain fatty acid production. PLOS Pathog. 2020;16:e1008448. doi: 10.1371/journal.ppat.1008448. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
65. Mathewson N.D., Jenq R., Mathew A.V., Koenigsknecht M., Hanash A., Toubai T., Oravecz-Wilson K., Wu S.R., Sun Y., Rossi C., et al. Gut microbiome-derived metabolites modulate intestinal epithelial cell damage and mitigate graft-versus-host disease. Nat. Immunol. 2016;17:505–513. doi: 10.1038/ni.3400. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
66. Hartstra A.V., Bouter K.E., Bäckhed F., Nieuwdorp M. Insights into the role of the microbiome in obesity and type 2 diabetes. Diabetes Care. 2015;38:159–165. doi: 10.2337/dc14-0769. [PubMed] [CrossRef] [Google Scholar]
67. Segain J.P., de la Blétière D.R., Bourreille A., Leray V., Gervois N., Rosales C., Ferrier L., Bonnet C., Blottière H.M., Galmiche J.P. Butyrate inhibits inflammatory responses through NFkappaB inhibition: Implications for Crohn’s disease. Gut. 2000;47:397–403. doi: 10.1136/gut.47.3.397. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
68. Gao Z., Yin J., Zhang J., Ward R.E., Martin R.J., Lefevre M., Cefalu W.T., Ye J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes. 2009;58:1509–1517. doi: 10.2337/db08-1637. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
69. Marizzoni M., Cattaneo A., Mirabelli P., Festari C., Lopizzo N., Nicolosi V., Mombelli E., Mazzelli M., Luongo D., Naviglio D., et al. Short-Chain Fatty Acids and Lipopolysaccharide as Mediators Between Gut Dysbiosis and Amyloid Pathology in Alzheimer’s Disease. J. Alzheimer’s Dis. 2020;78:683–697. doi: 10.3233/JAD-200306. [PubMed] [CrossRef] [Google Scholar]
70. Ryan P.M., Delzenne N.M. Chapter 18—Gut Microbiota and Metabolism. In: Hyland N., Stanton C., editors. The Gut-Brain Axis. Academic Press; Cambridge, MA, USA: 2016. pp. 391–401. [CrossRef] [Google Scholar]
71. van den Munckhof I.C.L., Kurilshikov A., Ter Horst R., Riksen N.P., Joosten L.A.B., Zhernakova A., Fu J., Keating S.T., Netea M.G., de Graaf J., et al. Role of gut microbiota in chronic low-grade inflammation as potential driver for atherosclerotic cardiovascular disease: A systematic review of human studies. Obes. Rev. 2018;19:1719–1734. doi: 10.1111/obr.12750. [PubMed] [CrossRef] [Google Scholar]
72. Furet J.P., Kong L.C., Tap J., Poitou C., Basdevant A., Bouillot J.L., Mariat D., Corthier G., Doré J., Henegar C., et al. Differential adaptation of human gut microbiota to bariatric surgery-induced weight loss: Links with metabolic and low-grade inflammation markers. Diabetes. 2010;59:3049–3057. doi: 10.2337/db10-0253. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
73. Rajkumar H., Mahmood N., Kumar M., Varikuti S.R., Challa H.R., Myakala S.P. Effect of probiotic (VSL#3) and omega-3 on lipid profile, insulin sensitivity, inflammatory markers, and gut colonization in overweight adults: A randomized, controlled trial. Mediat. Inflamm. 2014;2014:348959. doi: 10.1155/2014/348959. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
74. Cheke L.G., Bonnici H.M., Clayton N.S., Simons J.S. Obesity and insulin resistance are associated with reduced activity in core memory regions of the brain. Neuropsychologia. 2017;96:137–149. doi: 10.1016/j.neuropsychologia.2017.01.013. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
75. Caracciolo B., Xu W., Collins S., Fratiglioni L. Cognitive decline, dietary factors and gut–brain interactions. Mech. Ageing Dev. 2014;136-137:59–69. doi: 10.1016/j.mad.2013.11.011. [PubMed] [CrossRef] [Google Scholar]
76. Stadlbauer V., Engertsberger L., Komarova I., Feldbacher N., Leber B., Pichler G., Fink N., Scarpatetti M., Schippinger W., Schmidt R., et al. Dysbiosis, gut barrier dysfunction and inflammation in dementia: A pilot study. BMC Geriatr. 2020;20:248. doi: 10.1186/s12877-020-01644-2. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
77. Silva Y.P., Bernardi A., Frozza R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. (Lausanne) 2020;11 doi: 10.3389/fendo.2020.00025. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
79. Bienenstock J., Kunze W., Forsythe P. Microbiota and the gut–brain axis. Nutr. Rev. 2015;73:28–31. doi: 10.1093/nutrit/nuv019. [PubMed] [CrossRef] [Google Scholar]
80. Junges V.M., Closs V.E., Nogueira G.M., Gottlieb M.G.V. Crosstalk between Gut Microbiota and Central Nervous System: A Focus on Alzheimer’s Disease. Curr. Alzheimer Res. 2018;15:1179–1190. doi: 10.2174/1567205015666180904155908. [PubMed] [CrossRef] [Google Scholar]
81. Kennedy P.J., Cryan J.F., Dinan T.G., Clarke G. Kynurenine pathway metabolism and the microbiota-gut-brain axis. Neuropharmacology. 2017;112:399–412. doi: 10.1016/j.neuropharm.2016.07.002. [PubMed] [CrossRef] [Google Scholar]
82. Agus A., Planchais J., Sokol H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe. 2018;23:716–724. doi: 10.1016/j.chom.2018.05.003. [PubMed] [CrossRef] [Google Scholar]
83. Bosi A., Banfi D., Bistoletti M., Giaroni C., Baj A. Tryptophan Metabolites Along the Microbiota-Gut-Brain Axis: An Interkingdom Communication System Influencing the Gut in Health and Disease. Int. J. Tryptophan Res. IJTR. 2020;13 doi: 10.1177/1178646920928984. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
84. O’Farrell K., Harkin A. Stress-related regulation of the kynurenine pathway: Relevance to neuropsychiatric and degenerative disorders. Neuropharmacology. 2017;112:307–323. doi: 10.1016/j.neuropharm.2015.12.004. [PubMed] [CrossRef] [Google Scholar]
85. Strasser B., Becker K., Fuchs D., Gostner J.M. Kynurenine pathway metabolism and immune activation: Peripheral measurements in psychiatric and co-morbid conditions. Neuropharmacology. 2017;112:286–296. doi: 10.1016/j.neuropharm.2016.02.030. [PubMed] [CrossRef] [Google Scholar]
86. Wikoff W.R., Anfora A.T., Liu J., Schultz P.G., Lesley S.A., Peters E.C., Siuzdak G. Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proc. Natl. Acad. Sci. USA. 2009;106:3698–3703. doi: 10.1073/pnas.0812874106. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
87. Desbonnet L., Garrett L., Clarke G., Bienenstock J., Dinan T.G. The probiotic Bifidobacteria infantis: An assessment of potential antidepressant properties in the rat. J. Psychiatry Res. 2008;43:164–174. doi: 10.1016/j.jpsychires.2008.03.009. [PubMed] [CrossRef] [Google Scholar]
88. Martin-Gallausiaux C., Larraufie P., Jarry A., Béguet-Crespel F., Marinelli L., Ledue F., Reimann F., Blottière H.M., Lapaque N. Butyrate Produced by Commensal Bacteria Down-Regulates Indolamine 2,3-Dioxygenase 1 (IDO-1) Expression via a Dual Mechanism in Human Intestinal Epithelial Cells. Front. Immunol. 2018;9:2838. doi: 10.3389/fimmu.2018.02838. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
89. Roager H.M., Licht T.R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018;9:3294. doi: 10.1038/s41467-018-05470-4. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
90. Leblhuber F., Steiner K., Schuetz B., Fuchs D., Gostner J.M. Probiotic Supplementation in Patients with Alzheimer’s Dementia – An Explorative Intervention Study. Curr. Alzheimer Res. 2018;15:1106–1113. doi: 10.2174/1389200219666180813144834. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
91. Köhler C.A., Maes M., Slyepchenko A., Berk M., Solmi M., Lanctôt K.L., Carvalho A.F. The Gut-Brain Axis, Including the Microbiome, Leaky Gut and Bacterial Translocation: Mechanisms and Pathophysiological Role in Alzheimer’s Disease. Curr. Pharm. Des. 2016;22:6152–6166. doi: 10.2174/1381612822666160907093807. [PubMed] [CrossRef] [Google Scholar]
92. Seguella L., Capuano R., Sarnelli G., Esposito G. Play in advance against neurodegeneration: Exploring enteric glial cells in gut-brain axis during neurodegenerative diseases. Expert Rev. Clin. Pharmacol. 2019;12:555–564. doi: 10.1080/17512433.2019.1612744. [PubMed] [CrossRef] [Google Scholar]
93. Natale G., Pasquali L., Paparelli A., Fornai F. Parallel manifestations of neuropathologies in the enteric and central nervous systems. Neurogastroenterol. Motil. 2011;23:1056–1065. doi: 10.1111/j.1365-2982.2011.01794.x. [PubMed] [CrossRef] [Google Scholar]
94. Parekh P.J., Nayi V.R., Johnson D.A., Vinik A.I. The Role of Gut Microflora and the Cholinergic Anti-inflammatory Neuroendocrine System in Diabetes Mellitus. Front. Endocrinol. (Lausanne) 2016;7 doi: 10.3389/fendo.2016.00055. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
95. Liu A.K.L., Chang R.C.-C., Pearce R.K.B., Gentleman S.M. Nucleus basalis of Meynert revisited: Anatomy, history and differential involvement in Alzheimer’s and Parkinson’s disease. Acta Neuropathol. 2015;129:527–540. doi: 10.1007/s00401-015-1392-5. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
96. Hajishengallis G. Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015;15:30–44. doi: 10.1038/nri3785. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
97. Atarashi K., Suda W., Luo C., Kawaguchi T., Motoo I., Narushima S., Kiguchi Y., Yasuma K., Watanabe E., Tanoue T., et al. Ectopic colonization of oral bacteria in the intestine drives T(H)1 cell induction and inflammation. Science. 2017;358:359–365. doi: 10.1126/science.aan4526. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
98. Kodukula K., Faller D.V., Harpp D.N., Kanara I., Pernokas J., Pernokas M., Powers W.R., Soukos N.S., Steliou K., Moos W.H. Gut Microbiota and Salivary Diagnostics: The Mouth Is Salivating to Tell Us Something. BioResearch Open Access. 2017;6:123–132. doi: 10.1089/biores.2017.0020. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
99. Lira-Junior R., Boström E.A. Oral-gut connection: One step closer to an integrated view of the gastrointestinal tract? Mucosal Immunol. 2018;11:316–318. doi: 10.1038/mi.2017.116. [PubMed] [CrossRef] [Google Scholar]
100. Leblhuber F., Steiner K., Geisler S., Fuchs D., Gostner J.M. On the Possible Relevance of Bottom-up Pathways in the Pathogenesis of Alzheimer’s Disease. Curr. Top. Med. Chem. 2020;20:1415–1421. doi: 10.2174/1568026620666200514090359. [PubMed] [CrossRef] [Google Scholar]
101. Leblhuber F., Huemer J., Steiner K., Gostner J.M., Fuchs D. Knock-on effect of periodontitis to the pathogenesis of Alzheimer’s disease? Wien. Klin. Wochenschr. 2020;132:493–498. doi: 10.1007/s00508-020-01638-5. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
102. Slocum C., Kramer C., Genco C.A. Immune dysregulation mediated by the oral microbiome: Potential link to chronic inflammation and atherosclerosis. J. Intern. Med. 2016;280:114–128. doi: 10.1111/joim.12476. [PubMed] [CrossRef] [Google Scholar]
103. Wadhawan A., Reynolds M.A., Makkar H., Scott A.J., Potocki E., Hoisington A.J., Brenner L.A., Dagdag A., Lowry C.A., Dwivedi Y., et al. Periodontal Pathogens and Neuropsychiatric Health. Curr. Top. Med. Chem. 2020;20:1353–1397. doi: 10.2174/1568026620666200110161105. [PubMed] [CrossRef] [Google Scholar]
104. Olsen I., Singhrao S.K. Can oral infection be a risk factor for Alzheimer’s disease? J. Oral Microbiol. 2015;7:29143. doi: 10.3402/jom.v7.29143. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
105. Ide M., Harris M., Stevens A., Sussams R., Hopkins V., Culliford D., Fuller J., Ibbett P., Raybould R., Thomas R., et al. Periodontitis and Cognitive Decline in Alzheimer’s Disease. PLoS ONE. 2016;11:e0151081. doi: 10.1371/journal.pone.0151081. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
106. Sochocka M., Zwolińska K., Leszek J. The Infectious Etiology of Alzheimer’s Disease. Curr. Neuropharmacol. 2017;15:996–1009. doi: 10.2174/1570159X15666170313122937. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
107. Sato K., Takahashi N., Kato T., Matsuda Y., Yokoji M., Yamada M., Nakajima T., Kondo N., Endo N., Yamamoto R., et al. Aggravation of collagen-induced arthritis by orally administered Porphyromonas gingivalis through modulation of the gut microbiota and gut immune system. Sci. Rep. 2017;7:6955. doi: 10.1038/s41598-017-07196-7. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
108. Arimatsu K., Yamada H., Miyazawa H., Minagawa T., Nakajima M., Ryder M.I., Gotoh K., Motooka D., Nakamura S., Iida T., et al. Oral pathobiont induces systemic inflammation and metabolic changes associated with alteration of gut microbiota. Sci. Rep. 2014;4:4828. doi: 10.1038/srep04828. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
109. Komazaki R., Katagiri S., Takahashi H., Maekawa S., Shiba T., Takeuchi Y., Kitajima Y., Ohtsu A., Udagawa S., Sasaki N., et al. Periodontal pathogenic bacteria, Aggregatibacter actinomycetemcomitans affect non-alcoholic fatty liver disease by altering gut microbiota and glucose metabolism. Sci. Rep. 2017;7:13950. doi: 10.1038/s41598-017-14260-9. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
110. Poole S., Singhrao S.K., Kesavalu L., Curtis M.A., Crean S. Determining the presence of periodontopathic virulence factors in short-term postmortem Alzheimer’s disease brain tissue. J. Alzheimers Dis. 2013;36:665–677. doi: 10.3233/JAD-121918. [PubMed] [CrossRef] [Google Scholar]
111. Tremlett H., Bauer K.C., Appel-Cresswell S., Finlay B.B., Waubant E. The gut microbiome in human neurological disease: A review. Ann. Neurol. 2017;81:369–382. doi: 10.1002/ana.24901. [PubMed] [CrossRef] [Google Scholar]
112. Poole S., Singhrao S.K., Chukkapalli S., Rivera M., Velsko I., Kesavalu L., Crean S. Active invasion of Porphyromonas gingivalis and infection-induced complement activation in ApoE-/- mice brains. J. Alzheimers Dis. 2015;43:67–80. doi: 10.3233/JAD-140315. [PubMed] [CrossRef] [Google Scholar]
113. Ilievski V., Zuchowska P.K., Green S.J., Toth P.T., Ragozzino M.E., Le K., Aljewari H.W., O’Brien-Simpson N.M., Reynolds E.C., Watanabe K. Chronic oral application of a periodontal pathogen results in brain inflammation, neurodegeneration and amyloid beta production in wild type mice. PLoS ONE. 2018;13:e0204941. doi: 10.1371/journal.pone.0204941. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
114. Dominy S.S., Lynch C., Ermini F., Benedyk M., Marczyk A., Konradi A., Nguyen M., Haditsch U., Raha D., Griffin C., et al. Porphyromonas gingivalis in Alzheimer’s disease brains: Evidence for disease causation and treatment with small-molecule inhibitors. Sci. Adv. 2019;5:eaau3333. doi: 10.1126/sciadv.aau3333. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
115. Kern P., Rokos H., Dietrich M. Raised serum neopterin levels and imbalances of T-lymphocyte subsets in viral diseases, acquired immune deficiency and related lymphadenopathy syndromes. Biomed. Pharmacother. 1984;38:407–411. [PubMed] [Google Scholar]
116. Schennach H., Hessenberger G., Mayersbach P., Schönitzer D., Fuchs D. Acute cytomegalovirus infections in blood donors are indicated by increased serum neopterin concentrations. Med. Microbiol. Immunol. 2002;191:115–118. doi: 10.1007/s00430-002-0148-8. [PubMed] [CrossRef] [Google Scholar]
117. Qin Q., Li Y. Herpesviral infections and antimicrobial protection for Alzheimer’s disease: Implications for prevention and treatment. J. Med. Virol. 2019;91:1368–1377. doi: 10.1002/jmv.25481. [PubMed] [CrossRef] [Google Scholar]
118. Harris S.A., Harris E.A. Molecular Mechanisms for Herpes Simplex Virus Type 1 Pathogenesis in Alzheimer’s Disease. Front. Aging Neurosci. 2018;10:48. doi: 10.3389/fnagi.2018.00048. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
119. Lee K.H., Kwon D.E., Do Han K., La Y., Han S.H. Association between cytomegalovirus end-organ diseases and moderate-to-severe dementia: A population-based cohort study. BMC Neurol. 2020;20:216. doi: 10.1186/s12883-020-01776-3. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
120. Strandberg T.E., Pitkala K.H., Linnavuori K., Tilvis R.S. Cognitive impairment and infectious burden in the elderly. Arch. Gerontol. Geriatr. Suppl. 2004:419–423. doi: 10.1016/j.archger.2004.04.053. [PubMed] [CrossRef] [Google Scholar]
121. Lövheim H., Olsson J., Weidung B., Johansson A., Eriksson S., Hallmans G., Elgh F. Interaction between Cytomegalovirus and Herpes Simplex Virus Type 1 Associated with the Risk of Alzheimer’s Disease Development. J. Alzheimers Dis. 2018;61:939–945. doi: 10.3233/JAD-161305. [PubMed] [CrossRef] [Google Scholar]
122. Giridharan V.V., Masud F., Petronilho F., Dal-Pizzol F., Barichello T. Infection-Induced Systemic Inflammation Is a Potential Driver of Alzheimer’s Disease Progression. Front. Aging Neurosci. 2019;11:122. doi: 10.3389/fnagi.2019.00122. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
123. Iwashyna T.J., Ely E.W., Smith D.M., Langa K.M. Long-term Cognitive Impairment and Functional Disability Among Survivors of Severe Sepsis. JAMA. 2010;304:1787–1794. doi: 10.1001/jama.2010.1553. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
124. Soscia S.J., Kirby J.E., Washicosky K.J., Tucker S.M., Ingelsson M., Hyman B., Burton M.A., Goldstein L.E., Duong S., Tanzi R.E., et al. The Alzheimer’s disease-associated amyloid beta-protein is an antimicrobial peptide. PLoS ONE. 2010;5:e9505. doi: 10.1371/journal.pone.0009505. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
125. Angelucci F., Cechova K., Amlerova J., Hort J. Antibiotics, gut microbiota, and Alzheimer’s disease. J. Neuroinflamm. 2019;16:108. doi: 10.1186/s12974-019-1494-4. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
126. Ravelli K.G., Rosário B.D., Camarini R., Hernandes M.S., Britto L.R. Intracerebroventricular Streptozotocin as a Model of Alzheimer’s Disease: Neurochemical and Behavioral Characterization in Mice. Neurotox. Res. 2017;31:327–333. doi: 10.1007/s12640-016-9684-7. [PubMed] [CrossRef] [Google Scholar]
127. Furman B.L. Streptozotocin-Induced Diabetic Models in Mice and Rats. Curr. Protoc. Pharmacol. 2015;70:5.47.41–45.47.20. doi: 10.1002/0471141755.ph0547s70. [CrossRef] [Google Scholar]
128. Walker J.M., Harrison F.E. Shared Neuropathological Characteristics of Obesity, Type 2 Diabetes and Alzheimer’s Disease: Impacts on Cognitive Decline. Nutrients. 2015;7:7332–7357. doi: 10.3390/nu7095341. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
129. Hao W.Z., Li X.J., Zhang P.W., Chen J.X. A review of antibiotics, depression, and the gut microbiome. Psychiatry Res. 2020;284:112691. doi: 10.1016/j.psychres.2019.112691. [PubMed] [CrossRef] [Google Scholar]
130. Desbonnet L., Clarke G., Traplin A., O’Sullivan O., Crispie F., Moloney R.D., Cotter P.D., Dinan T.G., Cryan J.F. Gut microbiota depletion from early adolescence in mice: Implications for brain and behaviour. Brain Behav. Immun. 2015;48:165–173. doi: 10.1016/j.bbi.2015.04.004. [PubMed] [CrossRef] [Google Scholar]
131. Blaser M.J. The theory of disappearing microbiota and the epidemics of chronic diseases. Nat. Rev. Immunol. 2017;17:461–463. doi: 10.1038/nri.2017.77. [PubMed] [CrossRef] [Google Scholar]
132. Blaser M.J. Missing Microbes: How the Overuse of Antibiotics is Fueling Our Modern Plagues. Henry Holt and Company; New York City, NY, USA: 2014. [PMC free article] [PubMed] [Google Scholar]
133. Faith J.J., Guruge J.L., Charbonneau M., Subramanian S., Seedorf H., Goodman A.L., Clemente J.C., Knight R., Heath A.C., Leibel R.L., et al. The Long-Term Stability of the Human Gut Microbiota. Science. 2013;341:1237439. doi: 10.1126/science.1237439. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
134. Turnbaugh P.J., Ridaura V.K., Faith J.J., Rey F.E., Knight R., Gordon J.I. The effect of diet on the human gut microbiome: A metagenomic analysis in humanized gnotobiotic mice. Sci. Trans. Med. 2009;1:6ra14–16ra14. doi: 10.1126/scitranslmed.3000322. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
135. Cho I., Yamanishi S., Cox L., Methé B.A., Zavadil J., Li K., Gao Z., Mahana D., Raju K., Teitler I., et al. Antibiotics in early life alter the murine colonic microbiome and adiposity. Nature. 2012;488:621–626. doi: 10.1038/nature11400. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
136. Magnusson K.R., Hauck L., Jeffrey B.M., Elias V., Humphrey A., Nath R., Perrone A., Bermudez L.E. Relationships between diet-related changes in the gut microbiome and cognitive flexibility. Neuroscience. 2015;300:128–140. doi: 10.1016/j.neuroscience.2015.05.016. [PubMed] [CrossRef] [Google Scholar]
137. Leeming E.R., Johnson A.J., Spector T.D., Le Roy C.I. Effect of Diet on the Gut Microbiota: Rethinking Intervention Duration. Nutrients. 2019;11:2862. doi: 10.3390/nu11122862. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
138. Christ A., Günther P., Lauterbach M.A.R., Duewell P., Biswas D., Pelka K., Scholz C.J., Oosting M., Haendler K., Baßler K., et al. Western Diet Triggers NLRP3-Dependent Innate Immune Reprogramming. Cell. 2018;172:162–175. doi: 10.1016/j.cell.2017.12.013. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
139. Nunemaker C.S., Chen M., Pei H., Kimble S.D., Keller S.R., Carter J.D., Yang Z., Smith K.M., Wu R., Bevard M.H., et al. 12-Lipoxygenase-knockout mice are resistant to inflammatory effects of obesity induced by Western diet. Am. J. Physiol. Endocrinol. Metab. 2008;295:E1065–E1075. doi: 10.1152/ajpendo.90371.2008. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
140. Schreyer S.A., Wilson D.L., LeBoeuf R.C. C57BL/6 mice fed high fat diets as models for diabetes-accelerated atherosclerosis. Atherosclerosis. 1998;136:17–24. doi: 10.1016/S0021-9150(97)00165-2. [PubMed] [CrossRef] [Google Scholar]
141. Surwit R.S., Kuhn C.M., Cochrane C., McCubbin J.A., Feinglos M.N. Diet-Induced Type II Diabetes in C57BL/6J Mice. Diabetes. 1988;37:1163–1167. doi: 10.2337/diab.37.9.1163. [PubMed] [CrossRef] [Google Scholar]
142. Huang E.Y., Devkota S., Moscoso D., Chang E.B., Leone V.A. The role of diet in triggering human inflammatory disorders in the modern age. Microbes Infect. 2013;15:765–774. doi: 10.1016/j.micinf.2013.07.004. [PubMed] [CrossRef] [Google Scholar]
144. Li J., Lee D.H., Hu J., Tabung F.K., Li Y., Bhupathiraju S.N., Rimm E.B., Rexrode K.M., Manson J.E., Willett W.C., et al. Dietary Inflammatory Potential and Risk of Cardiovascular Disease Among Men and Women in the U.S. J. Am. Coll. Cardiol. 2020;76:2181–2193. doi: 10.1016/j.jacc.2020.09.535. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
145. Moszak M., Szulińska M., Bogdański P. You Are What You Eat-The Relationship between Diet, Microbiota, and Metabolic Disorders-A Review. Nutrients. 2020;12:1096. doi: 10.3390/nu12041096. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
146. Molteni R., Barnard R.J., Ying Z., Roberts C.K., Gómez-Pinilla F. A high-fat, refined sugar diet reduces hippocampal brain-derived neurotrophic factor, neuronal plasticity, and learning. Neuroscience. 2002;112:803–814. doi: 10.1016/S0306-4522(02)00123-9. [PubMed] [CrossRef] [Google Scholar]
147. Bruce-Keller A.J., Salbaum J.M., Luo M., Blanchard E.T., Taylor C.M., Welsh D.A., Berthoud H.R. Obese-type gut microbiota induce neurobehavioral changes in the absence of obesity. Biol. Psychiatry. 2015;77:607–615. doi: 10.1016/j.biopsych.2014.07.012. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
148. Perrone L., Grant W.B. Observational and ecological studies of dietary advanced glycation end products in national diets and Alzheimer’s disease incidence and prevalence. J. Alzheimers Dis. 2015;45:965–979. doi: 10.3233/JAD-140720. [PubMed] [CrossRef] [Google Scholar]
149. Hoffman R., Gerber M. Food Processing and the Mediterranean Diet. Nutrients. 2015;7:7925–7964. doi: 10.3390/nu7095371. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
150. Berbaum K., Shanmugam K., Stuchbury G., Wiede F., Körner H., Münch G. Induction of novel cytokines and chemokines by advanced glycation endproducts determined with a cytometric bead array. Cytokine. 2008;41:198–203. doi: 10.1016/j.cyto.2007.11.012. [PubMed] [CrossRef] [Google Scholar]
152. Younessi P., Yoonessi A. Advanced glycation end-products and their receptor-mediated roles: Inflammation and oxidative stress. Iran. J. Med. Sci. 2011;36:154–166. [PMC free article] [PubMed] [Google Scholar]
153. Crane P.K., Walker R., Hubbard R.A., Li G., Nathan D.M., Zheng H., Haneuse S., Craft S., Montine T.J., Kahn S.E., et al. Glucose Levels and Risk of Dementia. N. Engl. J. Med. 2013;369:540–548. doi: 10.1056/NEJMoa1215740. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
154. Schmitz L., Kuglin R., Bae-Gartz I., Janoschek R., Appel S., Mesaros A., Jakovcevski I., Vohlen C., Handwerk M., Ensenauer R., et al. Hippocampal insulin resistance links maternal obesity with impaired neuronal plasticity in adult offspring. Psychoneuroendocrinology. 2018;89:46–52. doi: 10.1016/j.psyneuen.2017.12.023. [PubMed] [CrossRef] [Google Scholar]
155. Benedict C., Hallschmid M., Schmitz K., Schultes B., Ratter F., Fehm H.L., Born J., Kern W. Intranasal Insulin Improves Memory in Humans: Superiority of Insulin Aspart. Neuropsychopharmacology. 2007;32:239–243. doi: 10.1038/sj.npp.1301193. [PubMed] [CrossRef] [Google Scholar]
156. Adzovic L., Lynn A.E., D’Angelo H.M., Crockett A.M., Kaercher R.M., Royer S.E., Hopp S.C., Wenk G.L. Insulin improves memory and reduces chronic neuroinflammation in the hippocampus of young but not aged brains. J. Neuroinflamm. 2015;12:63. doi: 10.1186/s12974-015-0282-z. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
157. Craft S., Asthana S., Cook D.G., Baker L.D., Cherrier M., Purganan K., Wait C., Petrova A., Latendresse S., Watson G.S., et al. Insulin dose-response effects on memory and plasma amyloid precursor protein in Alzheimer’s disease: Interactions with apolipoprotein E genotype. Psychoneuroendocrinology. 2003;28:809–822. doi: 10.1016/S0306-4530(02)00087-2. [PubMed] [CrossRef] [Google Scholar]
158. Kullmann S., Heni M., Hallschmid M., Fritsche A., Preissl H., Häring H.U. Brain Insulin Resistance at the Crossroads of Metabolic and Cognitive Disorders in Humans. Physiol. Rev. 2016;96:1169–1209. doi: 10.1152/physrev.00032.2015. [PubMed] [CrossRef] [Google Scholar]
159. Hoyer S. Brain glucose and energy metabolism abnormalities in sporadic Alzheimer disease. Causes and consequences: An update. Exp. Gerontol. 2000;35:1363–1372. doi: 10.1016/S0531-5565(00)00156-X. [PubMed] [CrossRef] [Google Scholar]
160. Hoyer S. Glucose metabolism and insulin receptor signal transduction in Alzheimer disease. Eur. J. Pharmacol. 2004;490:115–125. doi: 10.1016/j.ejphar.2004.02.049. [PubMed] [CrossRef] [Google Scholar]
161. Yaffe K., Blackwell T., Whitmer R.A., Krueger K., Connor E.B. Glycosylated hemoglobin level and development of mild cognitive impairment or dementia in older women. J. Nutr. Health Aging. 2006;10:293–295. [PubMed] [Google Scholar]
162. Cheng G., Huang C., Deng H., Wang H. Diabetes as a risk factor for dementia and mild cognitive impairment: A meta-analysis of longitudinal studies. Intern. Med. J. 2012;42:484–491. doi: 10.1111/j.1445-5994.2012.02758.x. [PubMed] [CrossRef] [Google Scholar]
164. de la Monte S.M. Insulin resistance and Alzheimer’s disease. BMB Rep. 2009;42:475–481. doi: 10.5483/BMBRep.2009.42.8.475. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
165. Vrieze A., Van Nood E., Holleman F., Salojärvi J., Kootte R.S., Bartelsman J.F.W.M., Dallinga–Thie G.M., Ackermans M.T., Serlie M.J., Oozeer R., et al. Transfer of Intestinal Microbiota from Lean Donors Increases Insulin Sensitivity in Individuals With Metabolic Syndrome. Gastroenterology. 2012;143:913–916. doi: 10.1053/j.gastro.2012.06.031. [PubMed] [CrossRef] [Google Scholar]
166. Ridaura V.K., Faith J.J., Rey F.E., Cheng J., Duncan A.E., Kau A.L., Griffin N.W., Lombard V., Henrissat B., Bain J.R., et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science. 2013;341:1241214. doi: 10.1126/science.1241214. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
167. Lecomte V., Kaakoush N.O., Maloney C.A., Raipuria M., Huinao K.D., Mitchell H.M., Morris M.J. Changes in gut microbiota in rats fed a high fat diet correlate with obesity-associated metabolic parameters. PLoS ONE. 2015;10:e0126931. doi: 10.1371/journal.pone.0126931. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
169. Polito R., Di Meo I., Barbieri M., Daniele A., Paolisso G., Rizzo M.R. Adiponectin Role in Neurodegenerative Diseases: Focus on Nutrition Review. Int. J. Mol. Sci. 2020;21:9255. doi: 10.3390/ijms21239255. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
170. Garcia-Mantrana I., Selma-Royo M., Alcantara C., Collado M.C. Shifts on Gut Microbiota Associated to Mediterranean Diet Adherence and Specific Dietary Intakes on General Adult Population. Front. Microbiol. 2018;9:890. doi: 10.3389/fmicb.2018.00890. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
171. Casas R., Sacanella E., Estruch R. The immune protective effect of the Mediterranean diet against chronic low-grade inflammatory diseases. Endocr. Metab. Immune Disord. Drug Targets. 2014;14:245–254. doi: 10.2174/1871530314666140922153350. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
172. Wu L., Sun D. Adherence to Mediterranean diet and risk of developing cognitive disorders: An updated systematic review and meta-analysis of prospective cohort studies. Sci. Rep. 2017;7:41317. doi: 10.1038/srep41317. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
173. Berti V., Walters M., Sterling J., Quinn C.G., Logue M., Andrews R., Matthews D.C., Osorio R.S., Pupi A., Vallabhajosula S., et al. Mediterranean diet and 3-year Alzheimer brain biomarker changes in middle-aged adults. Neurology. 2018;90:e1789–e1798. doi: 10.1212/WNL.0000000000005527. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
174. Cherian L., Wang Y., Fakuda K., Leurgans S., Aggarwal N., Morris M. Mediterranean-Dash Intervention for Neurodegenerative Delay (MIND) Diet Slows Cognitive Decline After Stroke. J. Prev. Alzheimers Dis. 2019;6:267–273. doi: 10.14283/jpad.2019.28. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
175. Bartochowski Z., Conway J., Wallach Y., Chakkamparambil B., Alakkassery S., Grossberg G.T. Dietary Interventions to Prevent or Delay Alzheimer’s Disease: What the Evidence Shows. Curr. Nutr. Rep. 2020;9:210–225. doi: 10.1007/s13668-020-00333-1. [PubMed] [CrossRef] [Google Scholar]
176. Martínez-Lapiscina E.H., Clavero P., Toledo E., Estruch R., Salas-Salvadó J., San Julián B., Sanchez-Tainta A., Ros E., Valls-Pedret C., Martinez-Gonzalez M. Mediterranean diet improves cognition: The PREDIMED-NAVARRA randomised trial. J. Neurol. Neurosurg. Psychiatry. 2013;84:1318–1325. doi: 10.1136/jnnp-2012-304792. [PubMed] [CrossRef] [Google Scholar]
177. van den Brink A.C., Brouwer-Brolsma E.M., Berendsen A.A.M., van de Rest O. The Mediterranean, Dietary Approaches to Stop Hypertension (DASH), and Mediterranean-DASH Intervention for Neurodegenerative Delay (MIND) Diets Are Associated with Less Cognitive Decline and a Lower Risk of Alzheimer’s Disease-A Review. Adv. Nutr. 2019;10:1040–1065. doi: 10.1093/advances/nmz054. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
178. Ghosh T.S., Rampelli S., Jeffery I.B., Santoro A., Neto M., Capri M., Giampieri E., Jennings A., Candela M., Turroni S., et al. Mediterranean diet intervention alters the gut microbiome in older people reducing frailty and improving health status: The NU-AGE 1-year dietary intervention across five European countries. Gut. 2020;69:1218–1228. doi: 10.1136/gutjnl-2019-319654. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
179. Francis N. Intermittent Fasting and Brain Health: Efficacy and Potential Mechanisms of Action. OBM Geriatr. 2020;4:1–19. doi: 10.21926/obm.geriatr.2002121. [CrossRef] [Google Scholar]
180. de Cabo R., Mattson M.P. Effects of Intermittent Fasting on Health, Aging, and Disease. N. Engl. J. Med. 2019;381:2541–2551. doi: 10.1056/NEJMra1905136. [PubMed] [CrossRef] [Google Scholar]
182. Widner B., Leblhuber F., Walli J., Tilz G.P., Demel U., Fuchs D. Tryptophan degradation and immune activation in Alzheimer’s disease. J. Neural Transm. (Vienna) 2000;107:343–353. doi: 10.1007/s007020050029. [PubMed] [CrossRef] [Google Scholar]
183. Leblhuber F., Walli J., Tilz G.P., Wachter H., Fuchs D. Systemic changes of the immune system in patients with Alzheimer’s dementia. Dtsch. Med. Wochenschr. 1998;123:787–791. doi: 10.1055/s-2007-1024069. [PubMed] [CrossRef] [Google Scholar]
184. Blasko I., Knaus G., Weiss E., Kemmler G., Winkler C., Falkensammer G., Griesmacher A., Würzner R., Marksteiner J., Fuchs D. Cognitive deterioration in Alzheimer’s disease is accompanied by increase of plasma neopterin. J. Psychiatry Res. 2007;41:694–701. doi: 10.1016/j.jpsychires.2006.02.001. [PubMed] [CrossRef] [Google Scholar]
186. Frick B., Gruber B., Schroecksnadel K., Leblhuber F., Fuchs D. Homocysteine but not neopterin declines in demented patients on B vitamins. J. Neural Transm. (Vienna) 2006;113:1815–1819. doi: 10.1007/s00702-006-0539-x. [PubMed] [CrossRef] [Google Scholar]
187. Jacobs K.R., Lim C.K., Blennow K., Zetterberg H., Chatterjee P., Martins R.N., Brew B.J., Guillemin G.J., Lovejoy D.B. Correlation between plasma and CSF concentrations of kynurenine pathway metabolites in Alzheimer’s disease and relationship to amyloid-β and tau. Neurobiol. Aging. 2019;80:11–20. doi: 10.1016/j.neurobiolaging.2019.03.015. [PubMed] [CrossRef] [Google Scholar]
188. Greilberger J., Fuchs D., Leblhuber F., Greilberger M., Wintersteiger R., Tafeit E. Carbonyl proteins as a clinical marker in Alzheimer’s disease and its relation to tryptophan degradation and immune activation. Clin. Lab. 2010;56:441–448. [PubMed] [Google Scholar]
189. Yoshii K., Hosomi K., Sawane K., Kunisawa J. Metabolism of Dietary and Microbial Vitamin B Family in the Regulation of Host Immunity. Front. Nutr. 2019;6:48. doi: 10.3389/fnut.2019.00048. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
190. Smith A.D., Refsum H., Bottiglieri T., Fenech M., Hooshmand B., McCaddon A., Miller J.W., Rosenberg I.H., Obeid R. Homocysteine and Dementia: An International Consensus Statement. J. Alzheimer’s Dis. 2018;62:561–570. doi: 10.3233/JAD-171042. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
191. Herrmann W., Obeid R. Homocysteine: A biomarker in neurodegenerative diseases. Clin. Chem. Lab. Med. 2011;49:435–441. doi: 10.1515/CCLM.2011.084. [PubMed] [CrossRef] [Google Scholar]
192. Schroecksnadel K., Leblhuber F., Frick B., Wirleitner B., Fuchs D. Association of Hyperhomocysteinemia in Alzheimer Disease with Elevated Neopterin Levels. Alzheimer Dis. Assoc. Disord. 2004;18:129–133. doi: 10.1097/01.wad.0000127443.23312.31. [PubMed] [CrossRef] [Google Scholar]
193. Fangmann D., Theismann E.M., Türk K., Schulte D.M., Relling I., Hartmann K., Keppler J.K., Knipp J.R., Rehman A., Heinsen F.A., et al. Targeted Microbiome Intervention by Microencapsulated Delayed-Release Niacin Beneficially Affects Insulin Sensitivity in Humans. Diabetes Care. 2018;41:398–405. doi: 10.2337/dc17-1967. [PubMed] [CrossRef] [Google Scholar]
194. Kellar D., Craft S. Brain insulin resistance in Alzheimer’s disease and related disorders: Mechanisms and therapeutic approaches. Lancet Neurol. 2020;19:758–766. doi: 10.1016/S1474-4422(20)30231-3. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
195. Ravaglia G., Forti P., Maioli F., Martelli M., Servadei L., Brunetti N., Porcellini E., Licastro F. Homocysteine and folate as risk factors for dementia and Alzheimer disease. Am. J. Clin. Nutr. 2005;82:636–643. doi: 10.1093/ajcn/82.3.636. [PubMed] [CrossRef] [Google Scholar]
196. Oulhaj A., Refsum H., Beaumont H., Williams J., King E., Jacoby R., Smith A.D. Homocysteine as a predictor of cognitive decline in Alzheimer’s disease. Int. J. Geriatr. Psychiatry. 2010;25:82–90. doi: 10.1002/gps.2303. [PubMed] [CrossRef] [Google Scholar]
197. Clarke R., Smith A.D., Jobst K.A., Refsum H., Sutton L., Ueland P.M. Folate, Vitamin B12, and Serum Total Homocysteine Levels in Confirmed Alzheimer Disease. Arch. Neurol. 1998;55:1449–1455. doi: 10.1001/archneur.55.11.1449. [PubMed] [CrossRef] [Google Scholar]
198. Tucker K.L., Qiao N., Scott T., Rosenberg I., Spiro A., 3rd High homocysteine and low B vitamins predict cognitive decline in aging men: The Veterans Affairs Normative Aging Study. Am. J. Clin. Nutr. 2005;82:627–635. doi: 10.1093/ajcn/82.3.627. [PubMed] [CrossRef] [Google Scholar]
199. Widner B., Leblhuber F., Frick B., Laich A., Artner-Dworzak E., Fuchs D. Moderate hyperhomocysteinaemia and immune activation in Parkinson’s disease. J. Neural Transm. (Vienna) 2002;109:1445–1452. doi: 10.1007/s00702-002-0758-8. [PubMed] [CrossRef] [Google Scholar]
200. Schröcksnadel K., Wirleitner B., Winkler C., Fuchs D. Monitoring tryptophan metabolism in chronic immune activation. Clin. Chim. Acta. 2006;364:82–90. doi: 10.1016/j.cca.2005.06.013. [PubMed] [CrossRef] [Google Scholar]
201. Murr C., Grammer T.B., Kleber M.E., Meinitzer A., März W., Fuchs D. Low serum tryptophan predicts higher mortality in cardiovascular disease. Eur. J. Clin. Investig. 2015;45:247–254. doi: 10.1111/eci.12402. [PubMed] [CrossRef] [Google Scholar]
202. Schroecksnadel K., Grammer T.B., Boehm B.O., März W., Fuchs D. Total homocysteine in patients with angiographic coronary artery disease correlates with inflammation markers. Thromb. Haemost. 2010;103:926–935. doi: 10.1160/TH09-07-0422. [PubMed] [CrossRef] [Google Scholar]
203. Grammer T.B., Fuchs D., Boehm B.O., Winkelmann B.R., Maerz W. Neopterin as a predictor of total and cardiovascular mortality in individuals undergoing angiography in the Ludwigshafen Risk and Cardiovascular Health study. Clin. Chem. 2009;55:1135–1146. doi: 10.1373/clinchem.2008.118844. [PubMed] [CrossRef] [Google Scholar]
204. Lanser L., Pölzl G., Fuchs D., Weiss G., Kurz K. Neopterin is Associated with Disease Severity and Outcome in Patients with Non-Ischaemic Heart Failure. J. Clin. Med. 2019;8:2230. doi: 10.3390/jcm8122230. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
205. Murr C., Winklhofer-Roob B.M., Schroecksnadel K., Maritschnegg M., Mangge H., Böhm B.O., Winkelmann B.R., März W., Fuchs D. Inverse association between serum concentrations of neopterin and antioxidants in patients with and without angiographic coronary artery disease. Atherosclerosis. 2009;202:543–549. doi: 10.1016/j.atherosclerosis.2008.04.047. [PubMed] [CrossRef] [Google Scholar]
206. Murr C., Pilz S., Tanja B.G., Marcus E.K., Meinitzer A., Bernhard O.B., März W., Fuchs D. Vitamin D deficiency parallels inflammation and immune activation, the Ludwigshafen Risk and Cardiovascular Health (LURIC) study. Clin. Chem. Lab. Med. 2012;50:2205. doi: 10.1515/cclm-2012-0157. [PubMed] [CrossRef] [Google Scholar]
207. de Wilde M.C., Vellas B., Girault E., Yavuz A.C., Sijben J.W. Lower brain and blood nutrient status in Alzheimer’s disease: Results from meta-analyses. Alzheimers Dement. (N. Y.) 2017;3:416–431. doi: 10.1016/j.trci.2017.06.002. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
208. Landel V., Annweiler C., Millet P., Morello M., Féron F. Vitamin D, Cognition and Alzheimer’s Disease: The Therapeutic Benefit is in the D-Tails. J. Alzheimers Dis. 2016;53:419–444. doi: 10.3233/JAD-150943. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
209. Annweiler C., Dursun E., Féron F., Gezen-Ak D., Kalueff A.V., Littlejohns T., Llewellyn D.J., Millet P., Scott T., Tucker K.L., et al. Vitamin D and cognition in older adults’: Updated international recommendations. J. Intern. Med. 2015;277:45–57. doi: 10.1111/joim.12279. [PubMed] [CrossRef] [Google Scholar]
210. Aquilani R., Costa A., Maestri R., Ramusino M.C., Pierobon A., Dossena M., Solerte S.B., Condino A.M., Torlaschi V., Bini P., et al. Mini Nutritional Assessment May Identify a Dual Pattern of Perturbed Plasma Amino Acids in Patients with Alzheimer’s Disease: A Window to Metabolic and Physical Rehabilitation? Nutrients. 2020;12:1845. doi: 10.3390/nu12061845. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
211. Lourida I., Hannon E., Littlejohns T.J., Langa K.M., Hyppönen E., Kuźma E., Llewellyn D.J. Association of Lifestyle and Genetic Risk with Incidence of Dementia. JAMA. 2019;322:430–437. doi: 10.1001/jama.2019.9879. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
212. Stephen R., Hongisto K., Solomon A., Lönnroos E. Physical Activity and Alzheimer’s Disease: A Systematic Review. J. Gerontol. A Biol. Sci. Med. Sci. 2017;72:733–739. doi: 10.1093/gerona/glw251. [PubMed] [CrossRef] [Google Scholar]
213. Colcombe S.J., Erickson K.I., Scalf P.E., Kim J.S., Prakash R., McAuley E., Elavsky S., Marquez D.X., Hu L., Kramer A.F. Aerobic exercise training increases brain volume in aging humans. J. Gerontol. A Biol. Sci. Med. Sci. 2006;61:1166–1170. doi: 10.1093/gerona/61.11.1166. [PubMed] [CrossRef] [Google Scholar]
214. Rovio S., Kåreholt I., Helkala E.L., Viitanen M., Winblad B., Tuomilehto J., Soininen H., Nissinen A., Kivipelto M. Leisure-time physical activity at midlife and the risk of dementia and Alzheimer’s disease. Lancet Neurol. 2005;4:705–711. doi: 10.1016/S1474-4422(05)70198-8. [PubMed] [CrossRef] [Google Scholar]
215. Kim Y.J., Han K.-D., Baek M.S., Cho H., Lee E.J., Lyoo C.H. Association between physical activity and conversion from mild cognitive impairment to dementia. Alzheimer’s Res. Ther. 2020;12:136. doi: 10.1186/s13195-020-00707-1. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
216. Bressa C., Bailén-Andrino M., Pérez-Santiago J., González-Soltero R., Pérez M., Montalvo-Lominchar M.G., Maté-Muñoz J.L., Domínguez R., Moreno D., Larrosa M. Differences in gut microbiota profile between women with active lifestyle and sedentary women. PLoS ONE. 2017;12:e0171352. doi: 10.1371/journal.pone.0171352. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
217. Bubu O.M., Brannick M., Mortimer J., Umasabor-Bubu O., Sebastião Y.V., Wen Y., Schwartz S., Borenstein A.R., Wu Y., Morgan D., et al. Sleep, Cognitive impairment, and Alzheimer’s disease: A Systematic Review and Meta-Analysis. Sleep. 2017;40 doi: 10.1093/sleep/zsw032. [PubMed] [CrossRef] [Google Scholar]
219. Medic G., Wille M., Hemels M.E. Short- and long-term health consequences of sleep disruption. Nat. Sci. Sleep. 2017;9:151–161. doi: 10.2147/NSS.S134864. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
220. Shokri-Kojori E., Wang G.-J., Wiers C.E., Demiral S.B., Guo M., Kim S.W., Lindgren E., Ramirez V., Zehra A., Freeman C., et al. β-Amyloid accumulation in the human brain after one night of sleep deprivation. Proc. Natl. Acad. Sci. USA. 2018;115:4483–4488. doi: 10.1073/pnas.1721694115. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
221. Kreutzmann J.C., Havekes R., Abel T., Meerlo P. Sleep deprivation and hippocampal vulnerability: Changes in neuronal plasticity, neurogenesis and cognitive function. Neuroscience. 2015;309:173–190. doi: 10.1016/j.neuroscience.2015.04.053. [PubMed] [CrossRef] [Google Scholar]
222. Ming G.L., Song H. Adult neurogenesis in the mammalian brain: Significant answers and significant questions. Neuron. 2011;70:687–702. doi: 10.1016/j.neuron.2011.05.001. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
224. Mueller A.D., Meerlo P., McGinty D., Mistlberger R.E. Sleep and adult neurogenesis: Implications for cognition and mood. Curr. Top. Behav. Neurosci. 2015;25:151–181. doi: 10.1007/7854_2013_251. [PubMed] [CrossRef] [Google Scholar]
225. Fernandes C., Rocha N.B., Rocha S., Herrera-Solís A., Salas-Pacheco J., García-García F., Murillo-Rodríguez E., Yuan T.F., Machado S., Arias-Carrión O. Detrimental role of prolonged sleep deprivation on adult neurogenesis. Front. Cell Neurosci. 2015;9:140. doi: 10.3389/fncel.2015.00140. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
226. Liu B., Lin W., Chen S., Xiang T., Yang Y., Yin Y., Xu G., Liu Z., Liu L., Pan J., et al. Gut Microbiota as an Objective Measurement for Auxiliary Diagnosis of Insomnia Disorder. Front. Microbiol. 2019;10 doi: 10.3389/fmicb.2019.01770. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
227. Smith R.P., Easson C., Lyle S.M., Kapoor R., Donnelly C.P., Davidson E.J., Parikh E., Lopez J.V., Tartar J.L. Gut microbiome diversity is associated with sleep physiology in humans. PLoS ONE. 2019;14:e0222394. doi: 10.1371/journal.pone.0222394. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
228. Canet G., Hernandez C., Zussy C., Chevallier N., Desrumaux C., Givalois L. Is AD a Stress-Related Disorder? Focus on the HPA Axis and Its Promising Therapeutic Targets. Front. Aging Neurosci. 2019;11:269. doi: 10.3389/fnagi.2019.00269. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
229. Lutgendorff F., Akkermans L.M., Söderholm J.D. The role of microbiota and probiotics in stress-induced gastro-intestinal damage. Curr. Mol. Med. 2008;8:282–298. doi: 10.2174/156652408784533779. [PubMed] [CrossRef] [Google Scholar]
230. Ávila-Villanueva M., Gómez-Ramírez J., Maestú F., Venero C., Ávila J., Fernández-Blázquez M.A. The Role of Chronic Stress as a Trigger for the Alzheimer Disease Continuum. Front. Aging Neurosci. 2020;12:561504. doi: 10.3389/fnagi.2020.561504. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
231. Sotiropoulos I., Catania C., Pinto L.G., Silva R., Pollerberg G.E., Takashima A., Sousa N., Almeida O.F. Stress acts cumulatively to precipitate Alzheimer’s disease-like tau pathology and cognitive deficits. J. Neurosci. 2011;31:7840–7847. doi: 10.1523/JNEUROSCI.0730-11.2011. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
232. Carroll J.C., Iba M., Bangasser D.A., Valentino R.J., James M.J., Brunden K.R., Lee V.M., Trojanowski J.Q. Chronic stress exacerbates tau pathology, neurodegeneration, and cognitive performance through a corticotropin-releasing factor receptor-dependent mechanism in a transgenic mouse model of tauopathy. J. Neurosci. 2011;31:14436–14449. doi: 10.1523/JNEUROSCI.3836-11.2011. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
233. Justice N.J., Huang L., Tian J.B., Cole A., Pruski M., Hunt A.J., Jr., Flores R., Zhu M.X., Arenkiel B.R., Zheng H. Posttraumatic stress disorder-like induction elevates β-amyloid levels, which directly activates corticotropin-releasing factor neurons to exacerbate stress responses. J. Neurosci. 2015;35:2612–2623. doi: 10.1523/JNEUROSCI.3333-14.2015. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
234. Rapp M.A., Schnaider-Beeri M., Wysocki M., Guerrero-Berroa E., Grossman H.T., Heinz A., Haroutunian V. Cognitive decline in patients with dementia as a function of depression. Am. J. Geriatr. Psychiatry. 2011;19:357–363. doi: 10.1097/JGP.0b013e3181e898d0. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
235. Du X., Pang T.Y. Is dysregulation of the HPA-axis a core pathophysiology mediating co-morbid depression in neurodegenerative diseases? Front. Psychiatry. 2015;6:32. doi: 10.3389/fpsyt.2015.00032. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
236. Woolley C.S., Gould E., McEwen B.S. Exposure to excess glucocorticoids alters dendritic morphology of adult hippocampal pyramidal neurons. Brain Res. 1990;531:225–231. doi: 10.1016/0006-8993(90)90778-A. [PubMed] [CrossRef] [Google Scholar]
237. Sousa N., Madeira M.D., Paula-Barbosa M.M. Effects of corticosterone treatment and rehabilitation on the hippocampal formation of neonatal and adult rats. An unbiased stereological study. Brain Res. 1998;794:199–210. doi: 10.1016/S0006-8993(98)00218-2. [PubMed] [CrossRef] [Google Scholar]
238. Csernansky J.G., Dong H., Fagan A.M., Wang L., Xiong C., Holtzman D.M., Morris J.C. Plasma cortisol and progression of dementia in subjects with Alzheimer-type dementia. Am. J. Psychiatry. 2006;163:2164–2169. doi: 10.1176/ajp.2006.163.12.2164. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
239. Doecke J.D., Laws S.M., Faux N.G., Wilson W., Burnham S.C., Lam C.P., Mondal A., Bedo J., Bush A.I., Brown B., et al. Blood-based protein biomarkers for diagnosis of Alzheimer disease. Arch. Neurol. 2012;69:1318–1325. doi: 10.1001/archneurol.2012.1282. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
240. Popp J., Wolfsgruber S., Heuser I., Peters O., Hüll M., Schröder J., Möller H.J., Lewczuk P., Schneider A., Jahn H., et al. Cerebrospinal fluid cortisol and clinical disease progression in MCI and dementia of Alzheimer’s type. Neurobiol. Aging. 2015;36:601–607. doi: 10.1016/j.neurobiolaging.2014.10.031. [PubMed] [CrossRef] [Google Scholar]
241. Taverniers J., Van Ruysseveldt J., Smeets T., von Grumbkow J. High-intensity stress elicits robust cortisol increases, and impairs working memory and visuo-spatial declarative memory in Special Forces candidates: A field experiment. Stress. 2010;13:323–333. doi: 10.3109/10253891003642394. [PubMed] [CrossRef] [Google Scholar]
242. Fung T.C., Olson C.A., Hsiao E.Y. Interactions between the microbiota, immune and nervous systems in health and disease. Nat. Neurosci. 2017;20:145–155. doi: 10.1038/nn.4476. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
243. Bharwani A., Mian M.F., Foster J.A., Surette M.G., Bienenstock J., Forsythe P. Structural & functional consequences of chronic psychosocial stress on the microbiome & host. Psychoneuroendocrinology. 2016;63:217–227. doi: 10.1016/j.psyneuen.2015.10.001. [PubMed] [CrossRef] [Google Scholar]
244. Liang S., Wang T., Hu X., Luo J., Li W., Wu X., Duan Y., Jin F. Administration of Lactobacillus helveticus NS8 improves behavioral, cognitive, and biochemical aberrations caused by chronic restraint stress. Neuroscience. 2015;310:561–577. doi: 10.1016/j.neuroscience.2015.09.033. [PubMed] [CrossRef] [Google Scholar]
245. Mu Y., Gage F.H. Adult hippocampal neurogenesis and its role in Alzheimer’s disease. Mol. Neurodegener. 2011;6:85. doi: 10.1186/1750-1326-6-85. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
246. Nehls M. Unified theory of Alzheimer’s disease (UTAD): Implications for prevention and curative therapy. J. Mol. Psychiatry. 2016;4:3. doi: 10.1186/s40303-016-0018-8. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
247. Bailey M.T., Dowd S.E., Galley J.D., Hufnagle A.R., Allen R.G., Lyte M. Exposure to a social stressor alters the structure of the intestinal microbiota: Implications for stressor-induced immunomodulation. Brain Behav. Immun. 2011;25:397–407. doi: 10.1016/j.bbi.2010.10.023. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
248. De Palma G., Collins S.M., Bercik P., Verdu E.F. The microbiota-gut-brain axis in gastrointestinal disorders: Stressed bugs, stressed brain or both? J. Physiol. 2014;592:2989–2997. doi: 10.1113/jphysiol.2014.273995. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
249. Karl J.P., Hatch A.M., Arcidiacono S.M., Pearce S.C., Pantoja-Feliciano I.G., Doherty L.A., Soares J.W. Effects of Psychological, Environmental and Physical Stressors on the Gut Microbiota. Front. Microbiol. 2018;9 doi: 10.3389/fmicb.2018.02013. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
250. Galley J.D., Bailey M.T. Impact of stressor exposure on the interplay between commensal microbiota and host inflammation. Gut Microbes. 2014;5:390–396. doi: 10.4161/gmic.28683. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
251. Gubert C., Kong G., Renoir T., Hannan A.J. Exercise, diet and stress as modulators of gut microbiota: Implications for neurodegenerative diseases. Neurobiol. Dis. 2020;134:104621. doi: 10.1016/j.nbd.2019.104621. [PubMed] [CrossRef] [Google Scholar]
252. Benjamin J.L., Hedin C.R., Koutsoumpas A., Ng S.C., McCarthy N.E., Prescott N.J., Pessoa-Lopes P., Mathew C.G., Sanderson J., Hart A.L., et al. Smokers with active Crohn’s disease have a clinically relevant dysbiosis of the gastrointestinal microbiota. Inflamm. Bowel Dis. 2012;18:1092–1100. doi: 10.1002/ibd.21864. [PubMed] [CrossRef] [Google Scholar]
253. Chin-Chan M., Navarro-Yepes J., Quintanilla-Vega B. Environmental pollutants as risk factors for neurodegenerative disorders: Alzheimer and Parkinson diseases. Front. Cell. Neurosci. 2015;9 doi: 10.3389/fncel.2015.00124. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
254. Vasefi M., Ghaboolian-Zare E., Abedelwahab H., Osu A. Environmental toxins and Alzheimer’s disease progression. Neurochem. Int. 2020;141:104852. doi: 10.1016/j.neuint.2020.104852. [PubMed] [CrossRef] [Google Scholar]
255. Duan H., Yu L., Tian F., Zhai Q., Fan L., Chen W. Gut microbiota: A target for heavy metal toxicity and a probiotic protective strategy. Sci. Total Environ. 2020;742:140429. doi: 10.1016/j.scitotenv.2020.140429. [PubMed] [CrossRef] [Google Scholar]
256. Aitbali Y., Ba-M’hamed S., Elhidar N., Nafis A., Soraa N., Bennis M. Glyphosate based- herbicide exposure affects gut microbiota, anxiety and depression-like behaviors in mice. Neurotoxicol. Teratol. 2018;67:44–49. doi: 10.1016/j.ntt.2018.04.002. [PubMed] [CrossRef] [Google Scholar]
257. Ruuskanen S., Rainio M.J., Gómez-Gallego C., Selenius O., Salminen S., Collado M.C., Saikkonen K., Saloniemi I., Helander M. Glyphosate-based herbicides influence antioxidants, reproductive hormones and gut microbiome but not reproduction: A long-term experiment in an avian model. Environ. Pollut. 2020;266:115108. doi: 10.1016/j.envpol.2020.115108. [PubMed] [CrossRef] [Google Scholar]258.
Iqubal A., Ahmed M., Ahmad S., Sahoo CR, Iqubal MK, Haque SE Environmental neurotoxic pollutants: Review. Mediul. Sci. Poluarea. Res. Int. 2020; 27 :41175–41198. doi: 10.1007/s11356-020-10539-z. [ PubMed ] [ CrossRef ] [ Google Scholar ]259.
Claus SP, Guillou H., Ellero-Simatos S. Microbiota intestinală: un jucător major în toxicitatea poluanților de mediu? NPJ Biofilms Microbiomi. 2016; 2 :16003. doi: 10.1038/npjbiofilms.2016.3. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]260.
Panza F., Lozupone M., Solfrizzi V., Watling M., Imbimbo BP Timpul de testare a terapiei antibacteriene în boala Alzheimer. Creier. 2019; 142 :2905–2929. doi: 10.1093/brain/awz244. [ PubMed ] [ CrossRef ] [ Google Scholar ]261.
Westman G., Berglund D., Widén J., Ingelsson M., Korsgren O., Lannfelt L., Sehlin D., Lidehall A.-K., Eriksson B.-M. Răspuns inflamator crescut la pacienții seropozitivi cu citomegalovirus cu boala Alzheimer. Plus unu. 2014; 9 :e96779. doi: 10.1371/journal.pone.0096779. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]263.
Zhang N., Ju Z., Zuo T. Time for food: The impact of diet on gut microbiota and human health. Nutriție. 2018; 51–52 :80–85. doi: 10.1016/j.nut.2017.12.005. [ PubMed ] [ CrossRef ] [ Google Scholar ]264.
Hills RD, Jr., Pontefract BA, Mishcon HR, Black CA, Sutton SC, Theberge CR Gut Microbiome: Profound Implications for Diet and Disease. Nutrienți. 2019; 11 :1613. doi: 10.3390/nu11071613. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]265.
Suez J., Zmora N., Segal E., Elinav E. Pro, contra și multe necunoscute ale probioticelor. Nat. Med. 2019; 25 :716–729. doi: 10.1038/s41591-019-0439-x. [ PubMed ] [ CrossRef ] [ Google Scholar ]266.
Verhoog S., Taneri PE, Roa Díaz ZM, Marques-Vidal P., Troup JP, Bally L., Franco OH, Glisic M., Muka T. Dietary Factors and Modulation of Bacteria Strains of Akkermansia muciniphila and Faecalibacterium prausnitzii: O revizuire sistematică. Nutrienți. 2019; 11 :1565. doi: 10.3390/nu11071565. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]267.
Siddarth P., Li Z., Miller KJ, Ercoli LM, Merril DA, Henning SM, Heber D., Small GW Studiu randomizat controlat cu placebo al efectelor de memorie ale sucului de rodie la adulții de vârstă mijlocie și în vârstă. A.m. J. Clin. Nutr. 2020; 111 :170–177. doi: 10.1093/ajcn/nqz241. [ PubMed ] [ CrossRef ] [ Google Scholar ]268.
Smirnov KS, Maier TV, Walker A., Heinzmann SS, Forcisi S., Martinez I., Walter J., Schmitt-Kopplin P. Challenges of metabolomics in human gut microbiota research. Int. J. Med. Microbiol. 2016; 306 :266–279. doi: 10.1016/j.ijmm.2016.03.006. [ PubMed ] [ CrossRef ] [ Google Scholar ]269.
Ford AH, Almeida OP Efectul suplimentării vitaminei B asupra funcției cognitive la vârstnici: o revizuire sistematică și meta-analiză. Îmbătrânirea drogurilor. 2019; 36 :419–434. doi: 10.1007/s40266-019-00649-w. [ PubMed ] [ CrossRef ] [ Google Scholar ]270.
Sharma V., Rodionov DA, Leyn SA, Tran D., Iablokov SN, Ding H., Peterson DA, Osterman AL, Peterson SN B-Vitamin Sharing Promotes Stability of Gut Microbial Communities. Față. Microbiol. 2019; 10 :1485. doi: 10.3389/fmicb.2019.01485. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]271.
Moreira SC, Jansen AK, Silva FM Intervenții dietetice și cunoaștere a pacienților cu boala Alzheimer: O revizuire sistematică a studiului controlat randomizat. Dement. Neuropsicol. 2020; 14 :258–282. doi: 10.1590/1980-57642020dn14-030008. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]272.
Costantini L., Molinari R., Farinon B., Merendino N. Impact of Omega-3 Fatty Acids on the Gut Microbiota. Int. J. Mol. Sci. 2017; 18 :2645. doi: 10.3390/ijms18122645. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]273.
Farina N., Llewellyn D., Isaac M., Tabet N. Vitamina E pentru demența Alzheimer și afectarea cognitivă ușoară. Cochrane Database Syst. Rev. 2017; 18 :4. doi: 10.1002/14651858.CD002854.pub5. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]274.
Stevens AJ, Purcell RV, Darling KA, Eggleston MJF, Kennedy MA, Rucklidge JJ Microbiomul intestinal uman se modifică în timpul unui studiu de control randomizat de 10 săptămâni pentru suplimentarea cu micronutrienți la copiii cu tulburare de hiperactivitate cu deficit de atenție. Sci. Rep. 2019; 9 :10128. doi: 10.1038/s41598-019-46146-3. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]275.
Zhou S.-S., Xu J., Zhu H., Wu J., Xu J.-D., Yan R., Li X.-Y., Liu H.-H., Duan S.- M., Wang Z., şi colab. Mecanismele implicate de microbiota intestinală în creșterea expunerii sistemice la ginsenozide prin polizaharidele coexistente în decoctul de ginseng. Sci. Rep. 2016; 6 :22474. doi: 10.1038/srep22474. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]276.
Caputo M., Bona E., Leone I., Samà MT, Nuzzo A., Ferrero A., Aimaretti G., Marzullo P., Prodam F. Inositols and metabolic disorders: From farm to bedside. J. Tradit. Completa. Med. 2020; 10 :252–259. doi: 10.1016/j.jtcme.2020.03.005. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]277.
Dias GP, Cavegn N., Nix A., do Nascimento Bevilaqua MC, Stangl D., Zainuddin MS, Nardi AE, Gardino PF, Thuret S. The role of dietary polyphenols on adult hippocampal neurogenesis: Molecular mechanisms and behavioral effects on depresie și anxietate. Oxid Med. Cell Longev. 2012; 2012 :541971. doi: 10.1155/2012/541971. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]278.
Stefani M., Rigacci S. Protein pliere și agregare în amiloid: interferența prin compuși fenolici naturali. Int. J. Mol. Sci. 2013; 14 :12411–12457. doi: 10.3390/ijms140612411. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]279.
Porzoor A., Alford B., Hügel HM, Grando D., Caine J., Macreadie I. Proprietăți anti-amiloidogenice ale unor compuși fenolici. Biomolecule. 2015; 5 :505–527. doi: 10.3390/biom5020505. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]280.
Small GW, Siddarth P., Li Z., Miller KJ, Ercoli L., Emerson ND, Martinez J., Wong KP, Liu J., Merrill DA și colab. Efectele memoriei și creierului amiloid și Tau ale unei forme biodisponibile de curcumină la adulții nedementați: un studiu dublu-orb, controlat cu placebo, de 18 luni. A.m. J. Geriatr. Psihiatrie. 2018; 26 :266–277. doi: 10.1016/j.jagp.2017.10.010. [ PubMed ] [ CrossRef ] [ Google Scholar ]281.
Voulgaropoulou SD, van Amelsvoort T., Prickaerts J., Vingerhoets C. Efectul curcuminei asupra cogniției în boala Alzheimer și îmbătrânirea sănătoasă: o revizuire sistematică a studiilor preclinice și clinice. Brain Res. 2019; 1725 :146476. doi: 10.1016/j.brainres.2019.146476. [ PubMed ] [ CrossRef ] [ Google Scholar ]282.
Chaplin A., Carpéné C., Mercader J. Resveratrol, sindrom metabolic și microbiota intestinală. Nutrienți. 2018; 10 :1651. doi: 10.3390/nu10111651. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]283.
Zam W. Microbiota intestinală ca țintă terapeutică prospectivă pentru curcumină: o revizuire a influenței reciproce. J. Nutr. Metab. 2018; 2018 :1367984. doi: 10.1155/2018/1367984. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]284.
Nady B., Bat-Erdene J., Anne P., Tharusha J., Hussein M., Mohammad NS, Perminder S., Ross G. Resveratrol ca potențial candidat terapeutic pentru tratamentul și managementul bolii Alzheimer. Curr. Top. Med. Chim. 2016; 16 :1951–1960. doi: 10.2174/1568026616666160204121431. [ PubMed ] [ CrossRef ] [ Google Scholar ]285.
Wu A., Noble EE, Tyagi E., Ying Z., Zhuang Y., Gomez-Pinilla F. Curcumin stimulează DHA în creier: Implicații pentru prevenirea tulburărilor de anxietate. Biochim. Biophys. Acta. 2015; 1852 :951–961. doi: 10.1016/j.bbadis.2014.12.005. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]287.
Arbo BD, André-Miral C., Nasre-Nasser RG, Schimith LE, Santos MG, Costa-Silva D., Muccillo-Baisch AL, Hort MA Resveratrol Derivatives as Potential Treatments for Alzheimer and Parkinson’s Disease. Față. Îmbătrânirea Neurosci. 2020; 12 :103. doi: 10.3389/fnagi.2020.00103. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]288.
Hayden KM, Beavers DP, Steck SE, Hebert JR, Tabung FK, Shivappa N., Casanova R., Manson JE, Padula CB, Salmoirago-Blotcher E., et al. Asocierea dintre o dietă inflamatorie și funcția cognitivă globală și demența incidentă la femeile în vârstă: Studiul de memorie a inițiativei pentru sănătatea femeilor. Dementul Alzheimer. 2017; 13 :1187–1196. doi: 10.1016/j.jalz.2017.04.004. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]289.
Omar SH Dietele mediteraneene și MIND care conțin biofenoli de măsline reduce prevalența bolii Alzheimer. Int. J. Mol. Sci. 2019; 20 :2797. doi: 10.3390/ijms20112797. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]290.
Holland TM, Agarwal P., Wang Y., Leurgans SE, Bennett DA, Booth SL, Morris MC Dietary flavonols and risk of Alzheimer dementa. Neurologie. 2020; 94 :e1749–e1756. doi: 10.1212/WNL.0000000000008981. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]291.
Pei R., Liu X., Bolling B. Flavonoide și sănătatea intestinului. Curr. Opinează. Biotehnologia. 2020; 61 :153–159. doi: 10.1016/j.copbio.2019.12.018. [ PubMed ] [ CrossRef ] [ Google Scholar ]292.
Barbagallo M., Marotta F., Dominguez LJ Stresul oxidativ la pacienții cu boala Alzheimer: Efectul extractelor de pudră de papaya fermentată. Mediat. Inflamm. 2015; 2015 :624801. doi: 10.1155/2015/624801. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]293.
Gutiérrez-Sarmiento W., Sáyago-Ayerdi SG, Goñi I., Gutiérrez-Miceli FA, Abud-Archila M., Rejón-Orantes JDC, Rincón-Rosales R., Peña-Ocaña BA Modificări ale microbiotei intestinale și metabolismului prezis căile în timpul fermentației colonice a fracțiunii indigerabile de bar pe bază de Mango (Mangifera indica L.). Nutrienți. 2020; 12 :683. doi: 10.3390/nu12030683. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]294.
Henning SM, Summanen PH, Lee R.-P., Yang J., Finegold SM, Heber D., Li Z. Elagitaninii de rodie stimulează creșterea Akkermansia muciniphila in vivo. Anaerob. 2017; 43 :56–60. doi: 10.1016/j.anaerobe.2016.12.003. [ PubMed ] [ CrossRef ] [ Google Scholar ]295.
Freund-Levi Y., Eriksdotter-Jönhagen M., Cederholm T., Basun H., Faxén-Irving G., Garlind A., Vedin I., Vessby B., Wahlund L.-O., Palmblad J. Tratamentul cu acizi grași ω-3 la 174 de pacienți cu boală Alzheimer uşoară până la moderată: studiu OmegaAD: un studiu randomizat dublu-orb. Arc. Neurol. 2006; 63 :1402–1408. doi: 10.1001/archneur.63.10.1402. [ PubMed ] [ CrossRef ] [ Google Scholar ]296.
Quinn JF, Raman R., Thomas RG, Yurko-Mauro K., Nelson EB, Van Dyck C., Galvin JE, Emond J., Jack CR, Weiner M., et al. Suplimentarea cu acid docosahexaenoic și declinul cognitiv în boala Alzheimer: un studiu randomizat. JAMA. 2010; 304 :1903–1911. doi: 10.1001/jama.2010.1510. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]297.
Shinto L., Quinn J., Montine T., Dodge HH, Woodward W., Baldauf-Wagner S., Waichunas D., Bumgarner L., Bourdette D., Silbert L. și colab. Un studiu pilot randomizat, controlat cu placebo, a acizilor grași Omega-3 și a acidului alfa lipoic în boala Alzheimer. J. Alzheimer Dis. 2014; 38 :111–120. doi: 10.3233/JAD-130722. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]298.
Lee S.-T., Chu K., Sim J.-Y., Heo J.-H., Kim M. Panax Ginseng Enhances Cognitive Performance in Alzheimer Disease. Alzheimer Dis. conf. univ. dezordine. 2008; 22 :222–226. doi: 10.1097/WAD.0b013e31816c92e6. [ PubMed ] [ CrossRef ] [ Google Scholar ]299.
Heo J.-H., Lee S.-T., Chu K., Oh MJ, Park H.-J., Shim J.-Y., Kim M. Ginsengul procesat la căldură îmbunătățește funcția cognitivă la pacienți cu boala Alzheimer moderat severă. Nutr. Neurosci. 2012; 15 :278–282. doi: 10.1179/1476830512Y.0000000027. [ PubMed ] [ CrossRef ] [ Google Scholar ]300.
Barak Y., Levine J., Glasman A., Elizur A., Belmaker RH Tratamentul cu inozitol al bolii Alzheimer: un studiu dublu, încrucișat, controlat cu placebo. Prog. Neuro-Psihofarmacol. Biol. Psihiatrie. 1996; 20 :729–735. doi: 10.1016/0278-5846(96)00043-7. [ PubMed ] [ CrossRef ] [ Google Scholar ]301.
Mishra S., Palanivelu K. Efectul curcuminei (turmeric) asupra bolii Alzheimer: O prezentare generală. Ann. Indian Acad. Neurol. 2008; 11 :13–19. doi: 10.4103/0972-2327.40220. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]302.
An Y., Feng L., Zhang X., Wang Y., Wang Y., Tao L., Qin Z., Xiao R. Pot fi asociate aporturile alimentare și modelele biomarkerilor de folat, vitamina B6 și vitamina B12 cu afectare cognitivă prin hipermetilarea genelor NUDT15 și TXNRD1 legate de redox. Clin. Epigenetica. 2019; 11 :139. doi: 10.1186/s13148-019-0741-y. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]303.
Lefèvre-Arbogast S., Féart C., Dartigues J.-F., Helmer C., Letenneur L., Samieri C. Dietary B Vitamins and a 10-Year Risk of Dementa in Older Persons. Nutrienți. 2016; 8 :761. doi: 10.3390/nu8120761. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]304.
Akbari E., Asemi Z., Kakhaki RD, Bahmani F., Kouchaki E., Tamtaji OR, Hamidi GA, Salami M. Efectul suplimentării probiotice asupra funcției cognitive și a statusului metabolic în boala Alzheimer: un randomizat, dublu-orb și Trial controlat. Față. Îmbătrânirea Neurosci. 2016; 8 :256. doi: 10.3389/fnagi.2016.00256. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]305.
Tamtaji OR, Taghizadeh M., Kakhaki RD, Kouchaki E., Bahmani F., Borzabadi S., Oryan S., Mafi A., Asemi Z. Răspunsul clinic și metabolic la administrarea de probiotice la persoanele cu boala Parkinson: A randomizat , studiu dublu-orb, controlat cu placebo. Clin. Nutr. 2019; 38 :1031–1035. doi: 10.1016/j.clnu.2018.05.018. [ PubMed ] [ CrossRef ] [ Google Scholar ]306.
Barros-Santos T., Silva KSO, Libarino-Santos M., Gouveia C.-PE, Reis HS, Tamura EK, de Oliveira-Lima AJ, Berro LF, Uetanabaro APT, Marinho EAV Efectele tratamentului cronic cu tulpini noi de Lactobacillus plantarum asupra comportamentelor cognitive, anxioase și depresive la șoarecii masculi. Plus unu. 2020; 15 :e0234037. doi: 10.1371/journal.pone.0234037. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]307.
Wang Q.-J., Shen Y.-E., Wang X., Fu S., Zhang X., Zhang Y.-N., Wang R.-T. Tratamentul concomitent cu memantină și Lactobacillus plantarum atenuează tulburările cognitive la șoarecii APP/PS1. Aging (Albany NY) 2020; 12 :628–649. doi: 10.18632/aging.102645. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]308.
Xiao J., Katsumata N., Bernier F., Ohno K., Yamauchi Y., Odamaki T., Yoshikawa K., Ito K., Kaneko T. Probiotic Bifidobacterium breve în îmbunătățirea funcțiilor cognitive ale adulților în vârstă cu suspiciune ușoară Tulburări cognitive: un studiu randomizat, dublu-orb, controlat cu placebo. J. Alzheimer Dis. 2020; 77 :139–147. doi: 10.3233/JAD-200488. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]309.
Den H., Dong X., Chen M., Zou Z. Eficacitatea probioticelor asupra cogniției și biomarkeri ai inflamației și stresului oxidativ la adulții cu boala Alzheimer sau tulburări cognitive ușoare – O meta-analiză a studiilor controlate randomizate. Aging (Albany NY) 2020; 12 :4010–4039. doi: 10.18632/aging.102810. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]310.
Bagga D., Reichert JL, Koschutnig K., Aigner CS, Holzer P., Koskinen K., Moissl-Eichinger C., Schöpf V. Probioticele conduc microbiomul intestinal care declanșează semnăturile emoționale ale creierului. Microbii intestinali. 2018; 9 :486–496. doi: 10.1080/19490976.2018.1460015. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]312.
Distrutti E., O’Reilly J.-A., McDonald C., Cipriani S., Renga B., Lynch MA, Fiorucci S. Modulation of intestinal microbiota by the probiotic VSL#3 resets brain gene expression and ameliorates the deficit legat de vârstă în LTP. Plus unu. 2014; 9 :e106503. doi: 10.1371/journal.pone.0106503. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]313.
Moon M., Choi JG, Nam DW, Hong HS, Choi YJ, Oh MS, Mook-Jung I. Ghrelin ameliorează disfuncția cognitivă și neurodegenerarea la șoarecii injectați cu oligomer amiloid-β1-42 intrahipocampal. J. Alzheimer Dis. 2011; 23 :147–159. doi: 10.3233/JAD-2010-101263. [ PubMed ] [ CrossRef ] [ Google Scholar ]314.
Gomes S., Martins I., Fonseca AC, Oliveira CR, Resende R., Pereira CM Efectul protector al leptinei și grelinei împotriva toxicității induse de oligomerii amiloid-β într-o linie celulară hipotalamică. J. Neuroendocrinol. 2014; 26 :176–185. doi: 10.1111/jne.12138. [ PubMed ] [ CrossRef ] [ Google Scholar ]315.
Messaoudi M., Lalonde R., Violle N., Javelot H., Desor D., Nejdi A., Bisson JF, Rougeot C., Pichelin M., Cazaubiel M., et al. Evaluarea proprietăților psihotrope ale unei formulări probiotice (Lactobacillus helveticus R0052 și Bifidobacterium longum R0175) la șobolani și subiecți umani. Br. J. Nutr. 2011; 105 :755–764. doi: 10.1017/S0007114510004319. [ PubMed ] [ CrossRef ] [ Google Scholar ]316.
Agahi A., Hamidi GA, Daneshvar R., Hamdieh M., Soheili M., Alinaghipour A., Taba SME, Salami M. Contribuie severitatea bolii Alzheimer la sensibilitatea sa la modificarea microbiotei intestinale? Un studiu clinic dublu-orb. Față. Neurol. 2018; 9 :662. doi: 10.3389/fneur.2018.00662. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]317.
Rogers GB, Bruce KD Provocări și oportunități pentru terapia de transplant de microbiotă fecală. Epidemiol. Infecta. 2013; 141 :2235–2242. doi: 10.1017/S0950268813001362. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]318.
Parker A., Fonseca S., Carding SR Gut microbis and metabolites as modulators of hemato-brain barier integrity and creier health. Microbii intestinali. 2020; 11 :135–157. doi: 10.1080/19490976.2019.1638722. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]319.
Ngandu T., Lehtisalo J., Solomon A., Levälahti E., Ahtiluoto S., Antikainen R., Bäckman L., Hänninen T., Jula A., Laatikainen T., et al. O intervenție pe mai multe domenii de 2 ani de dietă, exerciții fizice, antrenament cognitiv și monitorizare a riscului vascular versus control pentru a preveni declinul cognitiv la persoanele în vârstă cu risc (FINGER): un studiu controlat randomizat. Lancet. 2015; 385 :2255–2263. doi: 10.1016/S0140-6736(15)60461-5. [ PubMed ] [ CrossRef ] [ Google Scholar ]320.
Kivipelto M., Mangialasche F., Snyder HM, Allegri R., Andrieu S., Arai H., Baker L., Belleville S., Brodaty H., Brucki SM, et al. World-Wide FINGERS Network: O abordare globală a reducerii riscurilor și prevenirii demenței. Dementul Alzheimer. 2020; 16 :1078–1094. doi: 10.1002/alz.12123. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]321.
Coley N., Ngandu T., Lehtisalo J., Soininen H., Vellas B., Richard E., Kivipelto M., Andrieu S. Adherence to multidomain interventions for dementa prevention: Data from the FINGER and MAPT trials. Dementul Alzheimer. 2019; 15 :729–741. doi: 10.1016/j.jalz.2019.03.005. [ PubMed ] [ CrossRef ] [ Google Scholar ]322.
Bredesen DE, John V. Therapeutics de generație următoare pentru boala Alzheimer. EMBO Mol. Med. 2013; 5 :795–798. doi: 10.1002/emmm.201202307. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]323.
Bredesen DE Inversarea declinului cognitiv: un nou program terapeutic. Aging (Albany NY) 2014; 6 :707–717. doi: 10.18632/aging.100690. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]324.
Bredesen DE, Amos EC, Canick J., Ackerley M., Raji C., Fiala M., Ahdidan J. Inversarea declinului cognitiv în boala Alzheimer. Aging (Albany NY) 2016; 8 :1250–1258. doi: 10.18632/aging.100981. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]325.
Bredesen D., Sharlin K., Jenkins D., Okuno M., Youngberg W., Cohen S., Stefani A., Brown R., Conger S., Tanio C. și colab. Inversarea declinului cognitiv: 100 de pacienți. J. Alzheimer Dis. Parc. 2018; 8 doi: 10.4172/2161-0460.1000450. [ CrossRef ] [ Google Scholar ]326.
Peng X., Xing P., Li X., Qian Y., Song F., Bai Z., Han G., Lei H. Towards Personalized Intervention for Alzheimer’s Disease. Genom. Proteom. Bioinform. 2016; 14 :289–297. doi: 10.1016/j.gpb.2016.01.006. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
Articolele de la
Nutrients sunt furnizate aici prin amabilitatea
Institutului Multidisciplinar de Publicare Digitală (MDPI)
Acest studiu pilot a urmărit să studieze fezabilitatea și efectele asupra calității vieții (QOL) și a bunăstării a postului pe termen scurt (STF) în timpul chimioterapiei la pacienții cu cancer ginecologic.
metode
Într-un studiu individual, randomizat, încrucișat cu pacienți cu cancer ginecologic, au fost incluse 4 până la 6 cicluri de chimioterapie planificate.Treizeci și patru de pacienți au fost randomizați la post termen scurt STF în prima jumătate a chimioterapiei urmată de dieta normocalorică (grupa A; n = 18) sau invers (grupa B; n = 16).Postul a început 36 de ore înainte și s-a încheiat la 24 de ore după chimioterapie (perioadă de 60 de ore).
QOL calitatea vietii a fost evaluata prin sistemul de măsurare FACIT.
Rezultate
Reducerea indusă de chimioterapie a calitatii vietii QOL a fost mai mică decât diferența minimă importantă (MID; FACT-G = 5) cu post pe termen scurt STF, dar mai mare decât MID pentru perioadele fără repetare.
Deteriorarea indusă de chimioterapie a FACIT-F totală a fost de 10,4 ± 5,3 pentru pacienți postitori și 27,0 ± 6,3 pentru cicluri fără repaus alimentar în grupa A și 14,1 ± 5,6 pentru pacienți ne postitori și 11,0 ± 5,6 pentru cicluri de repaus alimentar/post în grupa B.
NU au existat efecte adverse grave.
Concluzie
postul pe termen scurt STF în timpul chimioterapiei este bine tolerat și pare să îmbunătățească calitatea vietii QOL și oboseala în timpul chimioterapiei.Studiile mai mari ar trebui să demonstreze efectul postului termen scurt STF ca adjuvant al chimioterapiei.
The effects of short-term fasting on quality of life and tolerance to chemotherapy in patients with breast and ovarian cancer: a randomized cross-over pilot study
Acest studiu a fost înregistrat la clinicaltrials.gov: NCT01954836 .
fundal
Experimental, starea de repaus alimentar pe termen scurt (STF) protejeaza celulele sanatoase impotriva efectelor adverse ale chimioterapiei, in timp ce celulele tumorale sunt mai vulnerabile la aceasta [ 1 ].Un număr tot mai mare de dovezi din cercetările de bază indică efectele benefice ale postului intermitent și periodic în bolile cronice [ 2 – 7 ].Postul promovează schimbări pronunțate în căile metabolice și în procesele celulare, cum ar fi răspunsul la stres (hormoză), autofagia și scade IGF-1 care afectează alți factori cum ar fi Akt, Ras și țintă de rapamicină (mTOR) la mamifere pentru a regla scăderea creșterii și proliferării celulare.La primate și la rozătoare restricțiile calorice și postul intermitent sunt asociate cu risc redus de cancer [9 , 10].Chimioterapia este un pilon principal în tratamentul tumorilor maligne, dar este frecvent limitat de toxicitatea sa.Recent, s-a demonstrat că efectele postului asupra susceptibilității la chimioterapie diferă între celulele normale și celulele canceroase, un fenomen descris ca rezistență la stres diferențial (DSR) [ 11 , 12 ].Datele experimentale indică faptul că starea de repaus alimentar poate promova protecția celulelor normale, dar nu a celulelor canceroase în timpul chimioterapiei, deoarece căile oncogene inhibă rezistența la stres.În celulele sănătoase, modificările expresiei metabolice și ale genei sunt induse de post, incluzând creșterea reglementării căilor de reparare a ADN-ului și a antioxidanților mediate parțial de închiderea căilor ca IGF-1 / Akt și mTOR [ 13 ].
Deoarece scăderea în greutate poate afecta negativ prognosticul pacienților cu cancer,postul pe termen scurt (STF), care NU are legătură cu pierderea în greutate pe termen lung și efectele sale adverse au fost introduse în cercetarea de bază ca potențial add-on tratament în timpul chimioterapie [ 14 ].Până în prezent, datele experimentale arată în mod constant că , combinația de cicluri de repaus alimentar pe termen scurt cu chimioterapie este eficientă în creșterea tolerabilității și eficacității chimioterapeutice și are astfel un potențial ridicat de translație [1 ].
Într-o primă serie de 10 pacienți cu diferite tipuri de cancer, STF sa dovedit a fi fezabil și a redus severitatea efectelor secundare induse de chimioterapie.[ 8 ].
Într-un studiu clinic uman pilot randomizat recent, 13 femei cu cancer de sân negativ HER2 care au primit chimioterapie neoadjuvante au fost randomizate la STF în timpul chimioterapiei sau pentru a mânca o dietă sănătoasă.Postul a redus toxicitatea hematologică a chimioterapiei și a indus o recuperare mai rapidă a deteriorării ADN [ 12 ].
Studiul-pilot prezent a fost conceput pentru a evalua efectul unui 60 h-STF asupra calității vieții (QOL) la pacienții cu cancer ginecologic sub chimioterapie.Pe baza dovezilor experimentale am presupus că postul crește QOL și reduce oboseala în timpul chimioterapiei comparativ cu alimentația standard [ 15 ].Prin intermediul unui proiect de explorare cross-over am comparat QOL, bunăstarea generală și oboseală în toate ciclurile de chimioterapie la care am fost supuși bolnavii cancer față de toate ciclurile de chimioterapie cu dietă normocalorică.
metode
Acest studiu a fost conceput ca un studiu randomizat, controlat individual, încrucișat.Proiectul de studiu a fost ales pe fondul heterogenității anticipate a populației de studiu și a protocoalelor de chimioterapie.Toți participanții la studiu și-au dat consimțământul în scris.Protocolul de studiu a fost revizuit și aprobat de Comitetul de etică al Centrului Medical Charité din Berlin (EA4 / 088/13).Pacienții au fost înscriși în perioada noiembrie 2013 – martie 2015;intervențiile și urmărirea au fost finalizate până în august 2015. Procedurile de studiu și colectarea datelor au fost efectuate la departamentul de ambulatoriu al Universității Charité din Berlin, la Immanuel Krankenhaus Berlin.
Proceduri de studiu
Pacienții au fost trimiși de trei departamente de spitale ginecologice, două centre de îngrijire a cancerului de sân și Centrul european de caritate ovarian Charité.Participanții potențiali au fost examinați pentru a fi eligibili în timpul unei întâlniri la centrul de studiu, iar candidații eligibili au fost programați pentru o întâlnire de înscriere.Fiecare participant eligibil a fost repartizat aleatoriu în două secvențe diferite de nutriție în timpul chimioterapiei programate.Grupa A a fost randomizată la un STF de 60 de ore în primele trei din cele 6 chimioterapii programate (36 de ore înainte până la 24 de ore după chimioterapie), urmată de o alimentație normocalorică în timpul următoarelor 3 chimioterapii.Grupa B a fost alocată unei secvențe de alimentație inversă.Între ciclurile de chimioterapie toți pacienții au fost sfătuiți să urmeze dieta lor obișnuită.Toți pacienții au primit o consiliere de aproximativ o oră de către dietiții cu experiență în tratamentele de repaus alimentar.
Toate măsurătorile au fost efectuate la momentul inițial și la 8 zile după fiecare ciclu de chimioterapie.Subiecții înălțime / greutate corporală au fost măsurate după un protocol standardizat.Pentru evaluarea QOL, s-au folosit inventare valide generale și stare de oboseală.Efectele adverse au fost evaluate prin două interviuri în timpul și la sfârșitul postului și prin intermediul unui jurnal.
pacienţii
Toate femeile au avut un diagnostic confirmat de cancer mamar sau cancer ovarian și o chimioterapie programată.Criteriile de eligibilitate au inclus vârsta ≥ 18 ani;IMC ≥ 19 kg / m 2 ;Starea de performanță OMS 0-2;speranța de viață anticipată de peste 3 luni;Criteriile de excludere includ: diabetul de tip 1 sau tratamentul intensificat cu insulină;infarctul miocardic, accident vascular cerebral sau embolie pulmonară în ultimele 3 luni;boală cardiacă instabilă;insuficiență renală, istoric al tulburărilor de alimentație;dementa, psihoza, afectarea mobilității fizice.
S-au examinat o sută douăzeci și unu de pacienți, 50 de pacienți au îndeplinit criteriile de includere și au fost recrutați la studiu.Diagrama fluxului pacientului este prezentată în figura 1 .Un pacient a anulat inițial tratamentele chimioterapice complete.Deși STF a fost în general bine tolerată, patru pacienți s-au retras din STF după ce au prezentat efecte secundare minore ale durerii de cap STF (două cazuri), hiperventilație în timpul primei chimioterapii (un caz) și slăbiciune generală subiectivă (un caz).Un pacient a raportat o aversiune față de substanțele nutritive pe bază de post.Astfel, în total 5 pacienți (10%) au renunțat la intervenția de repaus alimentar.Doi pacienți s-au retras din studiu din motive personale (probleme familiale), fiecare în faza de dietă.Opt subiecți nu au putut fi evaluați pentru urmăririle ulterioare ca urmare a neaderenței la studiu din cauza restricțiilor de timp și a refuzului complet de a completa alte documente de studiu (trei în fazele de repaus, cinci în fazele de dietă).Aceste abandonări nu au legătură cu efectele negative ale intervențiilor de studiu, evaluate prin interviuri telefonice.Au fost analizați un total de 34 de pacienți cu BC primar ( n = 29), BC avansați ( n = 1) și OC ( n = 4).
Toți pacienții au primit asistență oncologică standard, determinată de cerințele lor individuale.Pacienții din ambele grupuri au fost sfătuiți să-și mențină activitatea fizică regulată și să se abțină de la alte tratamente integrative sau de susținere noi în timpul perioadei de studiu.
Medicamente chimioterapeutice și terapie standard
Medicamentele chimioterapeutice utilizate în acest studiu au fost taxani (docetaxel (D), paclitaxel (T)), agenți de platină (carboplatină (P), ciclofosfamidă (C), antracicline (Epirubicină E, doxorubicină A metotrexatată fluorouracil (F), anticorp IgG1 bevacizumab (Avastin) și pentru pacienții cu supraexprimare HER2 / neu cu pertuzumab sau trastuzumab, care au fost administrate în diferite combinații standard, în conformitate cu protocoalele de tratament bazate pe indicații. EC, FEC-D, FEC-D + trastuzumab, AC-T, EC-T, TAC și D + pertuzumab + trastuzumab. + bevacizumab. S-au administrat antiemetice standard și medicamente pentru prevenirea reacțiilor de hipersensibilitate, inclusiv inhibitori de dexametazonă și 5HT3.
Intervenția postului
Toți pacienții au primit consiliere individuală în funcție de nevoile lor individuale, pentru a realiza corect programul de post și dieta normocalorică.Programul postului a fost adaptat la abordarea stabilită a postului modificat utilizat în spitalul nostru timp de mulți ani și evaluată în studiile efectuate la pacienții cu afecțiuni reumatismale și dureri cronice [ 16 ].Perioada de repaus a început 36 de ore înainte de chimioterapie și sa încheiat la 24 de ore după terminarea chimioterapiei, rezultând o perioadă totală de repaus de 60 de ore.În timpul perioadei de repaus subiecții au primit cantități nelimitate de apă, ceai din plante, suc de legume 2x100cl și mici cantități standardizate de bulion de legume ușoare, cu un aport total de energie totală de 350 kcal.Conformitatea cu regimul de repaus alimentar a fost evaluată prin apeluri telefonice și interviuri personale la sfârșitul ciclurilor de chimioterapie.
Perioada de hrană standard
Toți pacienții au fost sfătuiți să urmeze o dietă mediocră normocalorică pe parcursul acestei faze de studiu, inclusiv zilele de chimioterapie.
randomizare
Pacienții au fost repartizați aleatoriu în grupuri de tratament (bloc-randomizare cu variații aleatorie ale blocurilor (software statistic SAS / Base® (software statistic SAS / Base®)). nu a fost orbită.
Acest studiu randomizat, încrucișat, a avut ca scop să ofere o primă perspectivă asupra QOL și tolerabilității la chimioterapie la pacienții cu cancer de sân sau ovarieni cu STF versus dieta normocalorică în timpul chimioterapiei.Ca studiu pilot, scopul său a fost de a permite proiectarea unui alt proces de confirmare anticipat.
Mărimea eșantionului a fost calculată pentru grupa A începând cu STF, presupunând o marjă de echivalență de 5, adică MID de FACT-G, o diferență reală de 7 și o deviație standard de 3. O mărime de eșantion de 16 atinge o putere de 82% cu un nivel de semnificație de 0,05.Presupunând aceeași condiție pentru grupul B, obținem minim 32 de pacienți.
Variabilele continue cu o distribuție normală s-au exprimat ca valoare medie și deviație standard.Normalitatea a fost testată prin testul Shapiro-Wilks.Comparația statistică a caracteristicilor de bază și a rezultatelor a fost efectuată utilizând testul × 2 cu corecție Yates sau testul exact Fisher, atunci când este cazul, pentru variabilele categorice.Pentru variabilele continue am utilizat t-testul Studentului cu două ochiuri, respectiv testul independent Mann-Whitney U, pentru a testa ipoteza nulă că distribuția unei variabile este aceeași pe grupurile A și B.
Au fost efectuate calcule cu NCSS (versiunea 10), R (versiunea 3.1.0) și IBM SPSS Statistici pentru Windows (versiunea 22.0, Armonk, NY: IBM Corp).
Rezultate
Vârsta medie a celor 34 de pacienți a fost de 51y (intervalul 28-69y).La prima jumătate a ciclului de chimioterapie (grupul A), 18 pacienți au început cu postul, în timp ce ceilalți 16 pacienți (grupul B) au început cu dietă normală (Tabelul 1 ).
tabelul 1
Legendă: Caracteristici de bază pentru ambele grupuri.Grupa A: Postul pentru prima jumătate a ciclului de chimioterapie și consumul unei diete normocalorice pentru a doua jumătate a ciclului;Grupa B: Alimentația unei diete normocalorice pentru prima jumătate a ciclului de chimioterapie și de repaus pentru a doua jumătate a ciclului
Postul a fost în siguranță și toate reacțiile adverse raportate au fost de grad scăzut și la un nivel care nu a interferat cu activitățile zilnice.Efectele adverse minore pe parcursul tuturor ciclurilor, și în principal în timpul primului ciclu STF, au inclus cefaleea (5 ×), foamea (5 ×), ușurința greață după consumul de bulion sau sucuri (11 ×) și o reacție ortostatică.A fost o infecție a rănii, fără legătură cu postul.În ceea ce privește tolerabilitatea chimioterapiei, nu au fost documentate evenimente adverse de grad III / IV de toxicitate comună.
Modificări ale greutății corporale / indicele de masă corporală
Creșterea în greutate este o problemă obișnuită pentru pacienții cu cancer mamar / ovarian tratați cu chimioterapie și în special cu antracicline + taxani [ 18 ].În acest studiu nu au existat modificări semnificative ( p > 0,3) în greutate.Valoarea medie a greutății corporale (IMC) a pacienților din grupa A a fost de 73 kg (26,1) la început și 72,3 kg (25,8) la sfârșitul studiului.Pentru grupul B valorile corespunzătoare sunt 67,9 kg (23,7) și 68,5 kg (24,2).
Pe parcursul tuturor ciclurilor de chimioterapie a celor 34 de pacienți am documentat 102 cicluri de repaus și 74 de cicluri pe regim alimentar normocaloric.Diferența numerică între ciclurile de repaus și cele fără repaus este rezultatul a 5 pacienți care nu au dorit să treacă la regimul alimentar normocaloric după ce au postit primele 3 cicluri.Conformitatea cu protocolul postului, așa cum a fost verificată prin jurnale, apeluri telefonice și interviuri, părea bună.
De bază
Mijloacele diferitelor scale și subscale au fost mai slabe pentru pacienții cu cancer ovarian comparativ cu pacienții cu cancer mamar la momentul inițial (ziua 0, ciclul 1) (OC vs BC: FACT-G 66,8 (± 21,7) vs 80,5 (± 15,3); FACIT-F 33,5 (± 10,3) vs 39,7 (± 10,2); Total FACIT-F 100,3 (± 31,8) vs 120,2 (± 21,7)).Consistențele interne au fost excelente pentru diferitele chestionare: (alpha Cronbach: α> 0,9).FACT-G (α = 0,91), FACIT-F (α = 0,96), TOI FACIT-F (α = 0,97) și FACIT-F total (α = 0,96).Nu au existat statistic diferențe semnificative între FACT-G și subscalele între cele două grupuri la momentul inițial, cu excepția cazurilor de bunăstare socială / familială p = 0,042.
Cursuri de chimioterapie
Am utilizat un crossover AB × BA (A = SFT; B = dieta normocalorică) cu cross-over la ciclul 4. Cinci pacienți au avut un total de doar patru cicluri de chimioterapie, așa că am aliniat primele două cicluri la c1 și c2 și cele două cicluri secunde la c4 și c5.
Valorile medii ale tuturor scalelor în ziua 0 (înainte de chimioterapie) nu au fost semnificativ diferite pentru grupul A și B în cele 6 cicluri, adică pacienții s-au recuperat în 3 săptămâni de la fiecare ciclu de chimioterapie în ceea ce privește QOL (Fig.2a /b) Prin urmare, nu s-au evidențiat efecte de transmisie de la SFT, respectiv regim alimentar normocaloric, în primele trei cicluri până la a doua perioadă.
a : Valori pretestare la cicluri (C1-C6) în FACIT-F (FS), FACIT-F, FACIT-F TOI, FACT-G și Total FACIT-F, FACIT-F (FS), FACIT-F, FACIT-F TOI, FACT-G și Total FACIT-F Grupa B, Ziua 0
Modificarea QOL după chimioterapie
În cadrul grupului A am descoperit un efect benefic statistic și clinic al STF în timpul chimioterapiei (cicluri c1-c3) față de dieta normocalorică (cicluri c4-c6) asupra QOL și a oboselii.La ciclurile cu grupuri B (c4-c6) nu au fost asociate cu o reducere semnificativă a QOL indusă de chimioterapie și a oboselii comparativ cu dieta regulată (c1-c3) (Tabelul 2 ).
tabel 2
Deviațiile medii și standard pentru parametrii de rezultat cu diferențe medii de grup și CI 95% pentru FACT-G, FACIT-F, TOI FACIT-F și Total FACIT-F între ciclurile cu regim alimentar și regimul regulat din grupurile A și B în ziua 8 după chimioterapie
La analiza modificărilor de QOL în perioadele de repaus alimentar sau fără repaus prin aplicarea diferențelor minime importante (MID pentru FACT-G = 5) am descoperit pentru pacienții din grupul A, în primele trei cicluri posterioare, o scădere a FACT-G (medie = 3,0 ) care a fost mai mică decât IDM.Astfel, pacienții aflați sub STF nu au perceput schimbarea între FACT-G în ziua 0 și ziua 8 după chemoterapia ca fiind importantă.Dimpotrivă, schimbarea QOL pentru pacienții din grupul A în ciclurile de dietă normo-calorică c4-c6 a fost mai mare decât MID (media 12,8); acești pacienți au perceput că reducerea indusă de chimioterapie a QOL este importantă (Fig.3a , dd ].
FACT-G Parcela pădurii de diferență medie.a ) Grupa A (pretest – ziua 8, ciclurile c1-c3 = postul).b ) grupa B (pretest – ziua 8 (ciclurile c1 – c3 = normocalorice) c ) grupa B (pretest (c4) – ziua 8, ciclurile c4-c6 = 8, cicluri c4-c6 = normocalorice)
În consecință, diferențele medii (zi 0 – ziua 8) ale TOI FACIT-F din grupa A au fost 10,5 pentru primele trei cicluri și 25,1 pentru ciclurile 4-6 .Pacienții care au postit în primele trei cicluri au avut o diferență clinic semnificativ mai mică de TOI FACIT-F comparativ cu aceiași pacienți în timpul ciclurilor 4-6 pe o dietă regulată.
Diferențele medii ale TOI FACIT-F din grupa B au fost 13,1 pentru primele trei cicluri și 10,8 pentru ciclurile 4-6 .Pacienții care au postit în primele trei cicluri au avut o diferență clinic semnificativ mai mică de TOI FACIT-F comparativ cu aceiași pacienți în timpul ciclurilor 4-6 pe o dietă regulată.Nu a existat o diferență semnificativă între diferențele medii ale TOI FACIT-F pentru grupa A și grupul B în timpul ciclurilor de post (10,5 vs 10,8).
În grupul B nu am găsit diferențe semnificative statistic între primele 3 cicluri și celelalte trei cicluri.Cu toate acestea, la analiza modificărilor QOL în perioadele de repaus sau fără repaus alimentar, prin aplicarea MID, am constatat o medie a diferenței medii de FACT-G pentru ciclurile cu STF de 4,6 și cu dietă normocalorică de 8,1 [Fig. [ Fig.3b ,3b,,c].c ].Astfel, pacienții cu regim alimentar normocaloric au avut o diferență medie mai mare decât MID în acea fază, în timp ce cu STF diferența medie a fost de 4,6 adică sub MID.Fig.3a3a – -dd rezumă parcelele forestiere ale modificărilor induse de chimioterapie ale FACT-G în fiecare din cele două faze ale dietei în ambele grupuri.
În general, STF a fost foarte bine acceptat de către pacienți.În cadrul consultării finale, majoritatea pacienților au raportat o mai bună toleranță la chimioterapie cu STF, evaluarea generală a pacientului privind eficacitatea STF a fost „bună” sau „foarte bună” pentru 28 de pacienți, „moderată” la 5 pacienți și „fără efect” la 1 pacient.Treizeci și unu de pacienți au declarat că vor repeta din nou în timpul chimioterapiei, 3 pacienți au declarat că nu doresc să se repede în timpul chimioterapiei.
Discuţie
Acesta este primul studiu clinic pentru a explora efectele postirii pe termen scurt STF asupra calitatii vietii QOL, oboseală și bunăstare în timpul chimioterapiei.În mod experimental, STF postul pe termen scurt a fost documentat pentru a induce modificări profunde în expresia genelor și în metabolismul celular, care fac celulele normale mai rezistente la stresul oxidativ și astfel pot conferi beneficii în situația tratamentului cancerului prin chimioterapie [ 11 ].
Studiul prezent a inclus femeile cu cancer de sân și cancer ovarian și a utilizat un studiu randomizat încrucișat intra-individual pentru a echilibra heterogenitatea în stările de boală și protocoalele de chimioterapie.STF cu o perioadă de 60 de ore nu a determinat scăderea în greutate și a fost asociată doar cu efecte adverse minore care nu au fost evaluate de către pacienți și nu au afectat activitățile zilnice.
În general,postul pe termen scurt STF a condus la o toleranță mai bună la chimioterapie, cu o QOL mai puțin compromisa și oboseală redusă în termen de 8 zile după chimioterapie.
Până în prezent, există doar trei rapoarte anterioare în eșantioane mai mici care au evaluat efectele postul pe termen scurt STF în timpul chimioterapiei.Într-o primă serie de cazuri, Safdie et al.au descris 10 cazuri în care pacienții cu malignități diferite au postit voluntar înaintea și după chimioterapia cu diferite perioade ale perioadelor de repaus [ 8 ].Șase pacienți care au suferit chimioterapii cu și fără post, au raportat o reducere a oboselii, a slăbiciunii și a unor efecte secundare gastro-intestinale prin post.
Într-un studiu pilot recent, De Groot et al.a investigat o postire de 48 ore cu zero calorii în timpul chimioterapiei TAC la 13 femei cu cancer de sân confirmat cu HER-2-negativ în stadiul II și III.Pe lângă parametrii metabolici, endocrini și hematologici, a fost evaluată și deteriorarea ADN la PBMC, iar efectele secundare au fost evaluate în conformitate cu criteriile terminologice comune pentru evenimentele adverse (CTCAE).Doi pacienți s-au retras de la post după cea de-a treia chimioterapie din cauza unei deteriorări clinice care nu a avut legătură cu postul.Postul a fost sigur și a avut efecte benefice asupra toxicității hematologice și, eventual, asupra deteriorării ADN în celulele sănătoase (limfocite și celule mieloide) [ 19].
Rezultatele noastre confirmă fezabilitatea și tolerabilitatea postirii pe termen scurt STF care însoțesc chimioterapia și se extind asupra acestor rezultate indicând un potențial efect benefic asupra calitatii vietii QOL, obosealii și bunăstarii în timpul tratamentului pentru cancer.Deoarece calitatea vietii QOL este un rezultat al tratamentului din ce în ce mai apreciat, rezultatele prezente par a fi relevante clinic.
Nu am găsit nici o dovadă de malnutriție.
De remarcat că siguranța postului înainte de chimioterapie poate fi extrapolată numai la populația selectată de pacienți oncologi, deoarece excludem cei cu pierdere în greutate recentă sau un IMC <19,0 kg / m 2 .
Studiul nostru are câteva limitări. Limita cea mai evidentă este dimensiunea eșantionului mic în contextul unui studiu pilot experimental, care limitează puterea studiului și împiedică concluziile statistice ferme. A doua limitare este designul încrucișat, cu influența cunoscută a efectelor de transfer. Cu toate acestea, am ales acest design pe fondul dimensiunii mici a eșantionului definit și a probabilității respective de grupări eterogene care exclude compararea corectă a ciclurilor de chimioterapie cu randomizare comună. O altă limitare se datorează reputației pozitive a curelor de repaus în Germania. Ca atare, participanții la studiu ar putea fi predispuși la post, inducând astfel un efect nespecific. Acest lucru ar putea fi abordat doar cu un studiu randomizat dublu-orb, care, cu toate acestea, nu este fezabil.
Cu toate acestea, ne-am propus să includem mai mulți pacienți cu cancer ovarian, deoarece acești pacienți au fost deseori slăbiți prin intervenție chirurgicală, au prezentat mai frecvent pierdere în greutate anterioară și au fost mai frecvent incluși în alte studii clinice, opțiunile pentru recrutarea studiilor fiind limitate. Eterogenitatea populației studiate și tratamentele sale este o limitare pentru interpretarea rezultatelor, însă reflectă complexitatea actuală a chimioterapiilor aplicate și reflectă validitatea externă a rezultatelor. În cele din urmă, există o posibilitate de subreport și de nerespectare a postului, în ciuda încurajării tuturor subiecților de studiu să dezvăluie sincer toate alimentele și băuturile consumate în timpul perioadelor de post și monitorizarea de către nutriționiști. Inca,deoarece postul este stabilit pe scară largă și reputația pozitivă în Germania și toți participanții au primit informații detaliate înainte de includerea studiului despre schema postului credem că respectarea conformității raportată reflectă adevărata conformare cu protocolul de post.
De notat, efectul benefic al postului asupra calității vieții a fost un grup mai pronunțat în A comparativ cu grupul B. Există mai multe explicații posibile pentru acest efect diferențial. În primul rând, în ciuda randomizării, grupul A a prezentat statistic o importanță statistică nesemnificativă, dar semnificativă din punct de vedere clinic, a unei calități mai scăzute a vieții decât grupul B la momentul inițial. Acest lucru poate să fi influențat răspunsul la tratament al postului. În al doilea rând, ar putea fi faptul că postul este mai eficient dacă împiedică efectele negative înainte de a se produce mai degrabă decât după ce acestea apar. În al treilea rând, pacienții din grupa A ar fi putut experimenta mai pronunțat efecte nespecifice deoarece au postit mai întâi. În contrast, pacienții din grupul B si-ar fi putut reduce așteptările negative ale efectelor adverse ale chimioterapiei în primele cicluri, reducând astfel efectele nespecifice ale următoarei intervenții de repaus alimentar.
Concluzie
Studiul nostru arată că postirea pe termen scurt STF în timpul chimioterapiei este fezabilă și are efecte benefice asupra calitatii vietii QOL, bunăstării și oboselii. Sunt necesare studii mai mari, randomizate, cu studii de confirmare pentru a evalua în continuare această abordare inovatoare a tratamentului.
Finanțarea
Studiul a fost finanțat printr-un grant acordat de Günter und Regine Kelm Stiftung, Zürich, Elveția.
Disponibilitatea datelor și a materialelor
Seturile de date utilizate și / sau analizate în timpul studiului actual sunt disponibile de la autorul corespunzător, la cerere.
AM și SB au conceput studiul și au contribuit la achiziția de date. AM, AJ, RS, CK, BK, BB, JS au coordonat studiul. Analiza statistică a fost efectuată de MW și NS. AM, SB, CK, NS și MW au interpretat rezultatele. SB, AM și MW au scris și revizuit manuscrisul. Toți autori au citit și au aprobat versiunea finală a manuscrisului.
Protocolul de studiu a fost revizuit și aprobat de Comitetul de etică al Centrului Medical Charite, Berlin EA4 / 088/13. Toți participanții la studiu și-au dat consimțământul în scris.
1. Lee C, Raffaghello L, Brandhorst S, Safdie FM, Bianchi G, Martin-Montalvo A, Pistoia V, Wei M, Hwang S, Merlino A, și colab. Posturile de repaus retardă creșterea tumorilor și sensibilizează o gamă largă de tipuri de celule canceroase la chimioterapie. Sci Transl Med.2012;4 (124): 124ra127. doi: 10.1126 / scitranslmed.3003293.[ Articolul gratuit PMC ][ PubMed ][ Ref.cross ]
3. Tannenbaum A, Silverstone H. Influența gradului de restricție calorică asupra formării tumorilor pielii și a hepatoamelor la șoareci.Cancer Res.1949; 9 (12): 724-727.[ PubMed ]
4. Michalsen A, Schneider S, Rodenbeck A, Lüdtke R, Huether G, Dobos GJ. Efectele pe termen scurt ale postului asupra sistemului neuroendocrin la pacienții cu sindroame dureroase cronice. Nutr Neurosci.2003;6 (1): 11-18. doi: 10.1080 / 1028415021000042811.[ PubMed ][ Cross Ref ]
5. Michalsen A, Li C, Kaiser K, Lüdtke R, Meier L, Stange R, Kessler C. Tratamentul intern al fibromialgiei: o comparație neordonată a medicamentelor convenționale versus medicina integrativă, inclusiv terapia de repaus. Comportament bazat pe evidente Alternat Med.2013;2013 : 908610. doi: 10.1155 / 2013/908610.[ Articolul gratuit PMC ][ PubMed ][ Ref.cross ]
6. Choi IY, Piccio L, Childress P, Bollman B, Ghosh A, Brandhorst S, Suarez J, Michalsen A, Cross AH, Morgan TE și colab. Dietul imitând postul favorizează regenerarea și reduce simptomele autoimunității și sclerozei multiple. Replica celulară 2016; 15 (10): 2136-2146. doi: 10.1016 / j.celrep.2016.05.009.[ Articolul gratuit PMC ][ PubMed ][ Ref.cross ]
8. Safdie FM, Dorff T, Quinn D, Fontana L, Wei M, Lee C, Cohen P, Longo VD. Postul și tratamentul cancerului la om: un raport de caz. Îmbătrânire.2009;1 (12): 988-1007. doi: 10.18632 / aging.100114.[ Articolul gratuit PMC ][ PubMed ][ Ref.cross ]
9. Fontana L, Partridge L, Longo VD. Extinderea duratei de viață sănătoase – de la drojdie la om.Ştiinţă.2010;328 (5976): 321-326. doi: 10.1126 / science.1172539.[ Articolul gratuit PMC ][ PubMed ][ Ref.cross ]
10. Colman RJ, Anderson RM, Johnson SC, Kastman EK, Kosmatka KJ, Beasley TM, Allison DB, Cruzen C, Simmons HA, Kemnitz JW și colab. Restricția calorică întârzie declanșarea bolii și mortalitatea la maimuțele rhesus.Ştiinţă.2009;325 (5937): 201-204. doi: 10.1126 / science.1173635.[ Articolul gratuit PMC ][ PubMed ][ Ref.cross ]
11. Lee C, Longo VD. Postul vs. restricția dietetică în protecția celulară și tratamentul cancerului: de la organisme model la pacienți.Oncogene.2011;30 (30): 3305-3316. doi: 10.1038 / onc.2011.91.[ PubMed ][ Cross Ref ]
12. Raffaghello L, Lee C, Safdie FM, Wei M, Madia F, Bianchi G, Longo VD. Rezistența la tensiune diferențială dependentă de îngrijiri protejează celulele normale, dar nu și cele împotriva cancerului, împotriva chimioterapiei cu doze mari. Proc Natl Acad Sci SUA A. 2008; 105 (24): 8215-8220. doi: 10.1073 / pnas.0708100105.[ Articolul gratuit PMC ][ PubMed ][ Ref.cross ]
13. Lee C, Safdie FM, Raffaghello L, Wei M, Madia F, Parrella E, Hwang D, Cohen P, Bianchi G, Longo VD. Nivelurile reduse de IGF-I mediază protecția diferențială a celulelor normale și a cancerului ca răspuns la postul și îmbunătățirea indicelui chimioterapeutic.Cancer Res.2010;70 (4): 1564-1572. doi: 10.1158 / 0008-5472.CAN-09-3228.[ Articolul gratuit PMC ][ PubMed ][ Ref.cross ]
14. Michalsen A, Kuhlmann MK, Lüdtke R, Bäcker M, Langhorst J, Dobos GJ. Leziunile prelungite la pacienții cu sindroame dureroase cronice conduc la întârzierea imbunatatirii starii de spirit, care nu este legata de pierderea in greutate si de epuizarea leptinei induse de repaus. Nutr Neurosci.2006;9 (5-6): 195-200. doi: 10.1080 / 10284150600929656.[ PubMed ][ Cross Ref ]
15. Boschmann M, Michalsen A. Terapia prin posturi – perspective vechi și noi. Forschende Komplementärmedizin Res Complement Med.2013;20 (6): 410-411. doi: 10.1159 / 000357828.[ PubMed ][ Cross Ref ]
16. Michalsen A, Li C. Terapia de post pentru tratarea și prevenirea bolilor – stadiul actual al probelor. Forschende Komplementarmedizin (2006) 2013; 20 (6): 444-453. doi: 10.1159 / 000357765.[ PubMed ][ Cross Ref ]
17. Schunemann HJ, Akl EA, Guyatt GH. Interpretarea rezultatelor măsurilor rezultatelor raportate de pacienți în studiile clinice: perspectiva clinicianului. Rezultate privind calitatea vieții pentru sănătate.2006;4 : 62. doi: 10.1186 / 1477-7525-4-62.[ Articolul gratuit PMC ][ PubMed ][ Ref.cross ]
18. Winkels RM, Beijer S, van Lieshout R, van Barneveld D, Hofstede J, Kuiper J, Vreugdenhil A, van Warmerdam LJC, Schep G, Blaisse R, și colab. Modificări ale greutății corporale în timpul diferitelor tipuri de chimioterapie la pacienții cu cancer mamar. e-SPEN J. 2013; 9 (1): e39-44.
19. De Groot S, Vreeswijk MP, Welters MJ, Gravesteijn G, Boei JJ, Jochems A, Houtsma D, Putter H, van der Hoeven JJ, Nortier JW, și colab. Efectele postului pe termen scurt asupra toleranței la chimioterapia (neo) adjuvantă la pacienții cu cancer de sân HER2-negativ: un studiu pilot randomizat. BMC Cancer.2015;15 : 652. doi: 10.1186 / s12885-015-1663-5.[ Articolul gratuit PMC ][ PubMed ][ Ref.cross ]
20. Dorff TB, Groshen S, Garcia A, Shah M, Tsao-Wei D, Pham H, Cheng CW, Brandhorst S, Cohen P, Wei M, și colab. Siguranța și fezabilitatea postului în asociere cu chimioterapia pe bază de platină. BMC Cancer.2016;16 (1): 360. doi: 10.1186 / s12885-016-2370-6.[ Articolul gratuit PMC ][ PubMed ][ Ref.cross ]
Articolele de la BMC Cancer sunt oferite aici prin amabilitatea BioMed Central
Colostrul oral GcMAF este un aliment produs din colostru care conține celule ale sistemului imunitar și mulți anticorpi și ajută la stimularea sistemului imunitar.
Introducerea de „Saisei Pharma”
Saisei Pharma este de la Universitatea Tokushima în 2014, ce dezvolta Colostrul oral GcMAF si cerceteaza senzitizeri SDT și terapie câmp electric.Și are scopul de a trata infecția acută, infecție cronică, cancer, autism, sindromul oboselii cronice, scleroza multiplă, artrita reumatoidă, boala Alzheimer, dementa si multe alte boli.
Beneficiile ale colostru oral GcMAF
Ce este colostrul?
Colostrul este o formă de lapte produs de glandele mamare primele cateva zile dupa ce a dat naștere înainte să apară lapte adevărat.Colostrul conține anticorpi pentru a proteja nou-născuților împotriva bacteriilor, virusurilor și diferite tipuri de boli.
Anticorpii din colostrul
colostru de vaca conține imunoglobulină, cum ar fi IgG, IgA și IgM.Și este, de asemenea, ridicat în lactoferina, factor de creștere, minerale și vitamine.
Cercetarea asupra colostru de vacă – câteva rapoarte cu privire la eficacitatea asupra organismului uman.
1. stimularea sistemului imunitar
2. prevenirea bolilor infecțioase, cum ar fi rotavirusului raceala si gripa, si
3. reglementarea funcțiilor intestinelor
4. îmbunătățirea rezistenței musculare și fizice
5. anti-îmbătrânire
colostru bovine ca alimente
1. Aceasta este hrana din pulbere de colostru de bovine
2. colostru este similar cu componentele sanguine
3. Se poate lua fie sublingual, fie oral
4. Se activează macrofagele și ajută la stimularea sistemului imunitar
Colostru -efecte anti-îmbătrânire si asupra sistemului imunitar
Sistemul imunitar este un sistem facut pentru a menține corpul sănătos.Aceasta vă protejează de bacterii și viruși, precum și, de asemenea, va poate scapa de celulele canceroase.Cu toate acestea, funcția biologică și performanțele fizice ating maximul la varsta de 20 ani fiind în scădere la jumătate la 40 de ani.Pe masura ce sistemul imunitar slăbește cu vârsta, devine mai ușor să ne îmbolnăvim.Deci, este important să păstrați stimularea sistemului imunitar și sa-l păstrați ‘tânăr’,
Rolul macrofagelor
1. Funcții similare NK-Cells
Funcția macrofagelor ca și celulele NK care devoreaza si digeră celulele canceroase, virusi si microbi. Macrofagele sunt produse prin diferențierea monocitelor în țesuturi, care reprezintă 5 % din celulele albe din sânge.Aceasta este o celulă amoeboid și joacă un rol important în devorarea si digestia resturilor celulare, substanțelor străine, microbilor și celulele canceroase.se credea că au existat două feluri de diferențieri;macrofage care încurajează inflamatia numite macrofage M1 și macrofage care scad si incurajeaza repararea tesuturilor numite macrofage M2.
Dar, recent s-a crezut că există mai multe forme activate de macrofage, cum ar fi macrofage clasice activate, macrofagele de vindecare a rănilor, macrofage de supresie sau forme intermediare activate de macrofage.Astfel încât diversitatea de activare a macrofage rămâne încă necunoscuta.
Există mai multe rapoarte de caz de care au tratat cu macrofage activate HIV, cancer de san, cancerul de colon și cancerul de prostată .Astfel încât evaluarea și recunoașterea despre macrofage activate au început să fie în centrul atenției în tratarea multor tipuri de boli.
Acesta este procesul de modul în care macrofagele activate distrug celulele canceroase:
1. Macrofagele activate sunt combinate direct cu celulele canceroase.
2. macrofagele activate distrug celulele canceroase prin eliberarea substanțelor chimice, cum ar fi peroxidul de hidrogen.
Odată ce macrofagele ingereaza antigen, se elibereaza citokine si activeaza celule T specifice.Fragmentează substanțele străine pe care macrofagele le-au ingerat și descompus și le combină cu MHC-Ⅱ în celule și le prezintă pe suprafața celulei.Aceasta se numește prezentarea antigenului de către macrofage.
▲ macrofage distrug celulele canceroase
3. Efectul Antiangiogenic
Conform experimentelor pe animale, acesta este identificat că GcMAF are efect antiangiogenic(impiedica dezvoltarea vaselor de sange tumorale) asupra cancerului.
Semnalul prezentării antigenului de către macrofage este transmis in limfocite numite celule T Helper.
Celulele T helper mature exprima proteina de suprafață CD4 și proteina receptor al celulelor T – receptor.
Și fiecare proteină receptor a celulelor T helper se combină cu MHC-Ⅱ a macrofagelor și antigenul prezentat de macrofage, astfel încât macrofagelor încep să fie activate.
Structura receptorului proteina a celulei T este diferit în funcție de fiecare celulă T helper.Doar o celula T helper, care se potriveste cu un fragment de antigen prezentat de macrofage va fi activata.
Celulele T helper activate activează macrofagele prin producerea citokinelor, cum ar fi interleukina si limfokine si activeaza de asemenea si celulele B care recunosc același antigen(vezi celule B T NK STEM LAK etc)
Celulele B activate se diferențiază în celule de producere a anticorpului și apoi produc anticorp împotriva antigenului și apoi il elibereaza(pe anticorp impotriva antigenului).Și anticorpul specific se combină cu antigenul și apoi il înghite complex.Virusul si microbii pot fi înghițiti în mod eficient pentru macrofage.
Intre timp celulele T ajuta activarea macrofagelor , creșterea și diferențierea celulelor B prin eliberarea de limfokine.
Prin urmare, tratamentul de activare a macrofagelor e un tratament care face cel mai bun posibil din sistemului imunitar pe care fiecare persoană il are la origine.Mai mult decât atât, se poate spune că acest lucru este tratamentul care combină atât tratamentul cu celule NK cat si tratamentul cu celule dendritice(iata ,aveti si varianta pentru acasa a tratamentelor cancer scumpe de la clinici)
Ce este GcMAF?
Cercetatorii care au dezvoltat GcMAF
Acestea sunt fotografiile Dr. Yamamoto, care a dezvoltat prima generatie GcMAF, si Dr. Hitoshi Hori și Dr. Yoshihiro Uto de la Universitatea Tokushima, centrul de dezvoltare GcMAF în Japonia, în ultimii 20 de ani.
Istoria dezvoltării GcMaf
1991
Dr.Yamamoto descopera GcMAF.
1992
Dr.Yamamoto a vizitat centrulTokushima universitar.
1998
Dr.Uto sa alaturat echipei de dezvoltare.
2002
Prima lucrare de cercetare a fost publicata. (Prima generație GcMaf)
2010
Tokushima University si Saisei Mirai au început să facă cercetări și să dezvolte cea de a doua generație GcMaf împreună.
2011
Cea de a doua generație GcMaf a fost produs la Saisei Mirai Cell Culture Center ca studiu clinic, iar validarea clinică a început.
2012
Au trecut 20 de ani de când cercetarea GcMaf a început la Universitatea Tokushima.
2013
Peste 1000 de cazuri de a doua utilizare generație GcMaf au fost aplicate la Saisei Mirai.
2014
Lucrarea 3 a fost publicata pe Anticancer Research.
2015
centrul Tokushima universitar și Saisei Mirai au reușit în dezvoltarea Colostrul GcMAF pentru prima dată în lume.
Subtipul în funcție de diferența de lanț zahăr GcMaf
Există trei genotipuri, cum ar fi Gc1f, Gc1s și GC2 în Gc Subtip.Acolo devin șase tipuri în total, deoarece cei 2 dimeri ca homo și hetero sunt combinate ca fenotip. Aceasta este Gc1f1f, G1s1s, Gc22, Gc1f1s, Gc1f2 și Gc1s2.
Transformarea proteinei Gc în GcMAF
N.Yamamoto, și colab., J. Immunol., 151,2794-2802, 1993.
GcProtein este transformata în GcMaf ca răspuns la tăierea lanțului de zahăr activat β-galactozidaza din celule B activate și Sialidase din celulele T.
Activitatea fiziologică a GcMaf
Există șase activități importante: accelerarea activării fagocitare, creșterea producției de superoxid, inhibarea vascularizării, promovarea activității antitumorale și promovarea maturarii celulelor dendritică.Nu s-au raportat creșterea numărului de mononucleare din sânge și promovarea maturarii celulelor dendritice.Slide-urile de mai jos prezintă aceste cazuri.
Prima generație de teste de activitate GcMAF anti-tumorale
Nonaka K., Onizuka S., Uto Y., și colab., J. Surgical Res., 172, 116-22, 2012.
Aceasta sunt testele utilizate de șoareci.Grupul tratat cu prima generație GcMAF prezintă efectul inhibitor tumoral puternic comparativ cu cel netratat.Tumora nu s-a marit.
A doua generație Testele de activitate GcMaf anti-tumorale
Kuchiike D., Uto Y., și colab., Anticancer Res., 33, 2881-5, 2013. Uto Y., și colab., PCT / JP2012 / 072884
De asemenea, testele utilizate de șoareci.Linia neagră arată grupul netratat, iar linia punct verde arată cel tratat cu prima generație GcMAF.Aceasta arată în mod clar că rata de supraviețuire a fost prelungită a grupului tratat cu prima generație GcMAF. comparativ cu cea fără tratare
Ele sunt similare.Primul colostru de câteva ore după ce a dat naștere este deosebit de similar cu serul.Se spune că laptele de culoare roșie este produsă la început, după ce o vacă a dat o naștere.Dar, după 24 de ore, componenta a fost schimbat foarte mult și devine lapte regulat.
țesutului limfoid asociat intestine
patch-uri Peyer în cazul în care există o mulțime de macrofage
GALT sau țesutului limfoid asociat stomac, se spune ca este cel mai mare fond de macrofage din organism.Și este cunoscut faptul că o mulțime de macrofage există în patch-uri Payer.Colostrul MAF își propune să activeze în mod direct aceste macrofage(ATENTIE:chimioterapia distruge si stomacul-posbil sa nu mai avem cu ce lucra)Profesor Uto, Universitatea Tokushima, a început experimentul de bază al MAF colostru și GALT.Și noi așteaptăm cu nerăbdare rezultatele.
Activitatea fagocitară a Colostrul GcMAF
Acesta arată că prima generație 10ng GcMAF și 100ng colostrul MAF sunt aproape la fel ca activare.
in curs de brevetare: Nu 2013-257888
Acesta arată că 100ng Colostrul MAF ingerat oral și 10ng din prima generație GcMAF sunt aproape de aceeași activare.Prin urmare, este de așteptat ca utilizarea de 10 ori mai mult colostru MAA va aduce același efect ca și prima generație GcMAF. Colostrul oral MAF conține 200ng per capsulă.Așa că, dacă o capsula pe zi este luată, este de așteptat să câștige același efect ca și prima generație GcMAF.Dar este încă în curs de investigare.Ar trebui să fie posibil să conțină 1500ng per capsulă.
Sistemul limfatic și GcMAF
Prin stimularea directă a sistemului limfatic lângă partea lezată, macrofagele vor fi activate.
variante sublinguale, prin nebulizator, prin capsule sau prin supozitoare sunt luate în considerare.Se crede că ar trebui să fie mai eficiente pentru a stimula in mod direct sistemul limfatic aproape de partea lezată.O clinică din Elveția a folosit nebulizator cu ultrasunete pentru a lua GcMAF produs de ser.Dar atunci noi credem că va fi necesar pentru a experimenta pentru a vă asigura că în cazul în care structura GcMAF nu va fi modificata prin nebulizator cu ultrasunete.
Produse și experimente orale colostrul GcMAF
Imaginea produsului
Acest lucru se face în capsule și ambalate.
Experimentul de activitate fagocitara de macrofage
Activitatea fagocitară macrofaga este in curs de testare.Acest test este cel mai important test pentru a vedea funcția macrofagelor.
Cazuri de adaptare /aplicare colostru oral MAF
Cancer, autism, sindromul de oboseala cronica (CFS), boala lyme, boli infecțioase, cum ar fi gripa, virusul Noro, malaria, febra dengue, HIV, hepatita de tip B și de tip C, tuberculoza, boala autoimună etc
Se pare ca liderii in GcMaf cu rezultate in boli sunt acestia:
Printre altele au furnizat GcMaf la peste 11.000 de oameni, prin clinici si medici, precum și în interiorul propriilor clinici.
Au tratat cu succes 33 de tipuri diferite de cancer; GcMAF trateaza toate tipurile de cancer tumorale, ceea ce reprezintă 90% din ele. Și mai mult de 50 de alte boli majore.
Puteti vedea declarațiile persoanelor care s-au recuperat de la boala cu GcMAF al lor la adresa:
gcmaf.se/participants-results/ și gcmaf.se/news/
Și înregistrări video a zeci de oameni recuperati de boli majore la gcmaf.se/videos.
O companie israeliana Efranat face teste cu GcMAF. Ofera o doza de GcMAF cu $1,000, dar multi care au utilizat aceasta spun ca nu cred ca este eficient.
Dieta Ketogenica
Practic se mănâncă verdeturi, o mulțime de grăsimi permise in cancer -omega 3 si alte grasimi si uleiuri permise in cancer (inclusiv avocado, ulei de nucă de cocos, ulei de măsline, etc.) și peste 20 tablete de proteine MAP® opțional cu 100 de grame de ouă / pește / carne păsări de curte pe zi.
măsuraţi cetoza si glucoza din sânge. Sunteți în intervalul nutrițional cetoza la 0.5-3.0 mmol / l.
Niveluri de până la 6.0 pot avea loc de la infometare.
Daca mancati prea multi carbohidrati sau proteine prea multe (cu excepția MAP®) pe zi veti scadea din cetoza.
De asemenea, măsurati nivelurile de glucoză. Acestea ar trebui să scadă în mod progresiv și să rămână stabile la cea mai mică valoare posibilă și NU va crește după mese. Cu toate acestea, sa NU pierdeti in greutate – cresteti comprimate MAP până la 50 pe zi și să măncati mai multe nuci de macadamia si avocado pentru a opri pierderea in greutate. Suplimente de dieta
Vitamina D3 – 20,000IU pe zi. Nivelurile sanguine de vitamina D3, trebuie monitorizată și menținută între 150 și 250 nmol / l.
Ulei de măsline – 1 lingura de ulei de masline extra virgin de zi cu zi. Furnizează acidul oleic și Omega 3.
concentrat de tomate – 1 lingura amestecat cu ulei de masline pe zi ofera licopen si e deosebit de util pentru cancer de prostată. Opțional, dar de ajutor, în alte patologii..
Aspirina – 75mg pe zi. Încetează în caz de sângerare.
Apa curata alcalina- beti cel puțin doi litri de apă sau ceai de plante în fiecare zi. apa alcalina este recomandată.
iod din solutia Lugol – 2 picaturi pe zi, în cazul în care noduli tiroidieni sunt evidenți
Miscare moderata (prooxidativ)
exercitii fizice moderate(plimbare in parc, lucru in gradina) sau hipertermie pentru a stabili și menține un statut pro-oxidativ.
Produs prin fermentarea naturala cca 490ng OA-GcMAF în fiecare litru.
Conține lactoferinele și imunoglobuline.
Conține 42 de tulpini microbiene.
Conține Swiss colostru bio.
Este bogat în minerale, vitamine si molecule bio-active.
Folosite în clinicile de imunoterapie alături de injectabile GcMAF. suficient de puternic pentru a activa complet sistemul imunitar în termen de 3 săptămâni de utilizare.
sau capsule orale Colostru-MAF :
1000mcg colostru MAF ™-per capsulă.
Factorul de activare Macrofag – 100ng per capsulă.
Bogat în imunoglobuline și alți factori imunitari.
Capsulele rezistente la acizi emit MAF in intestin.
GcMAF intrarectal – Bravo Supozitoare
dozare minimă – 1 pentru a se introduce, de două ori pe săptămână, noaptea, înainte de un tratament sonoporation.
cancer pancreas, ficat sau cancer de colon – pentru un efect maxim, 1 in fiecare noapte dupa ce sa retras.
GcMAF în sistemul limfatic – in clinica
GcMAF injectat în apropierea ganglionilor limfatici situate din apropiere de masa tumorala Injecție este ghidata de imagistica cu ultrasunete. Dozare să fie determinată de către un medic, dar este de obicei 1500 + ng pe săptămână.
atentie la Rerum despre care se pretinde a fi inlocuitor de GcMaf .Iata ce spun unii despre Rerum
Rerum® oferă factorului de activare a macrofagelor asumat anterior GcMAF.
Rerum® este de dimensiuni nanoparticula(spre deosebire de GcMAF), astfel încât să poată ajunge la un număr mult mai mare de obiective în corpul uman.
Rerum® este extrem de biodisponibil în contrast cu proteine lungi, cum ar fi GcMAF.
Rerum® nu este derivat din sânge și, prin urmare, nu este afectat de nici una dintre problemele care afectează proteine derivate din sânge .
Rerum® nu are nevoie de nici o grijă deosebită pentru depozitare și transport, cum ar fi de refrigerare.
Rerum® este un supliment alimentar de bună credință.
Rerum® este fabricat într-o instalație de stat-of-domeniu certificate GMP în Germania.
si cum sta treaba de fapt :https://gcmaftruth.st/
GcMAF nebulizat in plămânii (prin nebulizer) – în clinica
GcMAF care urmează să fie livrate la plămâni într-un nebulizator cu abur. Dozare determinată de un medic, dar este de obicei 1500 + ng pe săptămână. Dozajul se dizolvă în 5 ml de ser fiziologic de fiecare dată. Sonoporatie – în clinica
Crește permeabilitătea membranei tumorale si intracelulare la droguri
Daca tratamente alternative cancer ce curata sangele de microbi si usureaza munca sistemului imunitar(suprapotenteaza sistemul imunitar) precum cel al Dr Bob Beck sau Dr Royal Rife s-au dovedit vindecatoare si de boli precum SIDA( intr-ucat si vitamina C IntraVenos, terapiile ozon/oxigen /apa ozonata/apa ionizata alcalina functioneaza pe acelasi principiu al curatarii sangelui de microorganisme – inclusiv de virusi- cu oxigen sau vitamina C si suprapotenteaza imunitatea se prea poate ca si acestea sa vindece SIDA), astazi voi discuta terapii nutritionale care sa ajute la vindecarea HIV/ SIDA.
urmatorul articol este scris de de Harold D. Foster phD (doctorat)
Universitatea din Victoria.British Columbia.Canada
(Retipărit cu permisiunea autorului)
De asemenea, vă recomandăm să descărcați, gratuit, publicarea Dr. Foster ,2006 , tratarea cu succes Orthomolecular de SIDA: Acumularea Dovezi din Africa:
SIDA este un deficit de boală cauzată de HIV (Virusul Imunodeficientei Umane).HIV-1 conține o genă care este practic identica cu cel care permite ca oamenii să producă enzima, glutation peroxidaza.Cand virusul este replicat, începe să concureze serios cu gazda sa pentru cele patru elemente nutritive necesare pentru a face aceasta enzima, în special elementul seleniu și trei aminoacizi, glutamina, cisteina si triptofan.Pe măsură ce crește infecția, deficiențe grave ale acestor substante nutritive se dezvolta.Seleniu inadecvat determină sistemul imunitar să se prăbușească, funcționarea defectuoasă a tiroidei și depresia se dezvolta.Deficit Glutamina duce la irosirea masei musculare si diaree.Lipsa de cisteină duce la probleme ale pielii cum ar fi psoriazisul și o mai mare sensibilitate la infectii.O lipsă de triptofan provoacă diaree, dermatita, dementa si in final moartea.Devine ușor pentru alti agenti patogeni de a infecta pacientul(cu un sitem imuntar slabit, evident) .Pe scurt, persoana infectata a dezvoltat tulburarea numita SIDA.Tratamentul HIV / SIDA, prin urmare, ar trebui să includă întotdeauna dietele ridicate în aceste patru elemente nutritive pentru a inversa astfel de deficiențe (PLUS-compeltez eu-eliminarea cauzei-a VIRUSULUI-cu una din metodele mentionate la inceputul articolului).Detalii privind modul în care acest lucru ar putea fi realizat sunt disponibile în discuție mai detaliată care urmează și în cartea „Care este cauza SIDA Într-adevăr”, care este disponibil gratuit pentru descărcare la http://www.hdfoster.com/publications(Nota editorului: I îndeamnă toți cititorii să citească aceasta carte importantă Dr. Foster.)
Dacă HIV-1 provoaca SIDA prin consumarea din organism de seleniu, cisteina, glutamina si triptofanul apoi modul de a ajuta tratarea aceastei tulburari este, evident, diete îmbogățite în aceste substante nutritive 1-2.) (IN PARALEL CU-completez eu- eliminarea virusului prin metodele mentionate la inceputul articolului-ca si in cancer-elimini si cauza dar in acelasi timp te asiguri ca supravietuiesti pe durata tratamentului prin corectarea deficitelor induse de cauza-de acest virus)
Dar cât de mult din fiecare este necesar?Care sunt nivelurile ideale de seleniu, cisteina, glutamina, și triptofanul în corpul uman?
SIDA și seleniu
Un studiu realizat de Shamberger și Willis 3 a descoperit ca indivizi sănătosi S.U.A., între 50 și 71 de ani, au în medie 21,7 micrograme de seleniu per 100 mililitri de sange.Aceasta in comparatie cu pacientii cu cancer de aceeasi varsta, care au fost gasiti pentru a avea doar 16,2 micrograme per 100 ml.În plus, într-un document ulterior, aceiași autori 4 descris o comparație între nivelurile de seleniu in sange de oameni din 19orașe S.U.A..Acestea au fost mai mari înRapid City.South Dakotași mai mici înLima.Ohio;25.6 și 15.7 micrograme per 100 ml. Interesant, Shamberger și Willis au găsit o relație inversă între ratele de deces clare datorita cancer în aceste orașe și niveluri de seleniu din sange.Cu atât mai mare seleniu, cu atat mai mică mortalitatea de cancer. Pentru a ilustra, înRapid City, În cazul în care nivelurile de seleniu din sange au fost cele mai mari, rata de deces de cancer anual la 100.000 de oameni, în timpul perioadei de 1962-1966, a fost 94,0.Prin contrast, înLima.Ohiocancer rata mortalitatii pentru această perioadă de timp a fost exact de două ori mai mare-188.0.Acest lucru sugereaza ca este mult mai bine pentru a avea un nivel de seleniu sânge de 25,6 micrograme per 100 ml decât una de 15,7.Astfel deS.U.A.diferențele regionale în nivelurile de seleniu sange au fost aproape sigur o reflectare a variațiilor în conținutul sol local a acestui mineral.
Braverman și Pfeiffer 5 au publicat date cu privire la diferite niveluri de aminoacizi din sange in ambele grupe de barbati adulți sănătoși și femei.Pentru cisteină astfel de niveluri variază de la 6 până la 14 micromoli per 100 mililitri pentru bărbați și 5 la 13 pentru femei.Nivelurile sanguine de glutamină date de acești autori variază de la 45 la 105 micromoli per 100 ml de bărbați adulți și 40-90 pentru femei adulte.În mod similar, cifrele citate de triptofan in sange pentru barbati si femei variază de la 4 la 25 micromoli per 100 ml.S-ar părea logic să încerce să atingă capetele înalte ale acestor intervale.
Alimente
Firește, o modalitate de a ridica nivelul corpului de seleniu și cei trei aminoacizi este de a mânca alimente în care apar cel mai frecvent.
„Care este cauza SIDA într-adevăr”http://www.hdfoster.com/publications conține anexe ce clasifica/coteaza / ranking alimente în funcție de conținutul lor de acid seleniu și amino.(Conținut de seleniu depinde în mare măsură solul în care a fost cultivat mâncarea.) Mai jos sunt câteva exemple de alimente care sunt în mod normal crescute în cel puțin unul dintre elementele nutritive esențiale: Seleniu, Cisteina, Glutamina, și triptofan.
Seleniu: nuci de Brazilia, usturoi, ciuperci, ficat, grau integral, orz, ou, pește și crustacee, „.
Cisteină: Brazilia nuci, usturoi, ceapa, broccoli,Bruxellesvarza, carne de pasăre, galbenusul de ou, iaurt, germeni de grau.
Glutamina: nuci de Brazilia, vânat sălbatic, germeni de grau, ovaz, granola, brânză de vaci, ricotta.
Triptofan: Migdale, caju, păsări de curte, hamsii, ouă, brânză de vaci și alte brânzeturi necoapte, ovăz, fulgi de ovaz, banane, ananas, iaurt.
Au fost dezbateri considerabile pe care forma de seleniu ar trebui să fie luate ca un supliment.Seleniu organic legat, de exemplu, în drojdia de bere, este considerabil cea mai eficientă în creșterea concentrațiilor sanguine ale acestui oligoelement decât este sodiu-selenit.(Passwater RA (1980). Seleniul ca produse alimentare și medicamente.New Canaan.CT:. Keats Publishing.pp 183-196) Cât de mult seleniu ar trebui să fie luat pe zi de indivizi HIV-pozitivi este subiect de discuție.Poate comentariile cele mai logice au venit de la dr Will Taylor 6
„(C) ercetarea a arătat că există probleme în absorbția nutrienților, chiar și în persoane infectate HIV + asimptomatice, sugestia a fost făcută ca pacientii cu HIV, trebuie să ia DOZE mai mari de vitamine decat persoanelor neinfectate pentru a atinge aceleași niveluri de sange. Din afirmatiile USDA cf care suplimentele alimentare în intervalul de 50-200 micrograme (mcg) de seleniu (SE) de zi cu zi este sigur si eficient pentru persoanele sanatoase, o doză de 400 mcg pare rezonabil pentru persoanele infectate cu HIV, în cazul în care nu au absorbția nutrientului afectată. Pentru un pacient SIDA care este demonstrabil deficitar in Se, o doză mai mare de zi cu zi (de până la 800 mcg) pentru o scurtă perioadă de timp (spun mai multe săptămâni) pentru a obtine nivelul lor de sange, urmată de o scădere la 400 mcg este o strategie eficientă, care a fost utilizata într-un studiu clinic publicat , ce a implicat pacientii cu SIDA. Această întrebare asupra niveluui de doză trezește în mod natural preocupări pentru că în trecut atât de mult au fost spuse ref toxicitatea potențială a Se. Cred că pericolul de toxicitate grave cu suplimentarea Se a fost exagerată.Amenințarea de toxicitate acută gravă cu suplimentarea este, în opinia mea inexistenta la doze mai mici de 1,000 mcg pe zi, în unele persoane.Astfel, dozele din intervalul 400 mcg sunt, fără îndoială siguranță.În orice caz, semnele de toxicitate cronică Se – mirosul de usturoi de respiratie si transpiratie, gust metalic în gură, păr fragil și unghiile – sunt foarte distinctive, și ușor de inversat prin scăderea dozei „.
Nu am nimic de adăugat.Dr. Taylor pare a fi corect, ca de obicei.
Glutamina
Suplimentele glutamină, în mod normal, sub formă de tablete de 500 miligrame, sunt disponibile în magazinele alimentare de sănătate.Pacienții cu SIDA sunt cunoscuti a fi foarte deficitari în glutamină.Într-un studiu la Harvard 7 persoanelor HIV-seropozitivi care au fost în mare parte asimptomatice, nivelurile serice glutamină s-au dovedit a fi foarte deprimate/scazute, chiar dacă acestea nu au aratat nici un semn de SIDA.În ciuda faptului că unui subgrup i-a fost dat 20 g (20.000 miligrame, adică 5 lingurițe) de glutamină pe zi în doze mici la 24 de ore timp de o luna, nivelurile sanguine de glutamină au rămas deprimate.Pacientiilor 8 le-au fost apoi date 40 de grame de glutamină pe zi, o cantitate de obicei rezervat pentru pacientii cu transplant de maduva osoasa lupta impotriva infectiei.
Pressman și Buff 9 au furnizat probabil cel mai bun sfat cu privire la utilizarea terapeutică a glutamina ca un supliment, atunci când ei au scris:
„Glutamina poate ajuta, de asemenea tratamentul diareei grave cauzate de SIDA sau de alte probleme intestinale, cum ar fi colita ulceroasa. Doze destul de mari de glutamina, fel de mare ca 40.000 de miligrame, pot fi necesare.Glutamina imbunatateste absorbtia apei prin colon, care ajuta scutirea diareei.Doze care mare ar trebui luate sub supraveghere, și numai de cei cu diaree cauzata de o problema medicala grava.Nu tratai diaree minora de indigestie sau un virus la stomac 24 de ore cu glutamină. „
Ca Shabert și Ehrlich 10 puncte din:
„Există cazuri foarte specifice în care da glutamină la un individ bolnav nu ar fi indicata. Persoanele care au ciroza severă a ficatului, sindromul Reye, sau o altă tulburare metabolică care poate duce la o acumulare de amoniac în sânge au risc crescut pentru encefalopatia sau comă. Problema de bază este o incapacitate de a goli corpul de azot în exces, care se transformă în amoniac și în final provoacă umflarea creierului și moartea cerebrala cu celule. Atunci cand ficatul este grav deteriorat sau când coma hepatică este iminentă, glutamină nu este eficientă și ar provoca numaideteriorarea in continuare la creier. „
CISTEINĂ
Suplimente de cisteină au fost folosite de mai mulți ani de către medici orthomolecular pentru a trata boli diferite, precum accident vascular cerebral, depresie maniacale, astm, si psihoza schizofrenică.Cisteina, totuși, este un aminoacid slab absorbite și trebuie să fie administrat în doze destul de mari.Pentru a cita Braverman și Pfeiffer 5:
„Când am stabilit că suplimentarea cisteina este necesara, vom începe, de obicei, cu o doză de 500 mg / zi. (Începând cu o cantitate mai mare poate duce la indigestie). Treptat, am putea crește doza la 3 sau 4 g pe zi. Intre timp , vom păstra un ochi pe valorile serice de cistina. Considerăm că, ca nivelurile de cistina vor reveni la valori plasmatice normale, scazute de zinc, acid folic și taurină, de asemenea, vor reveni la normal. Unii cercetători au folosit la fel de mult ca 7 g pe zi de cisteină. Trebuie remarcat faptul că dozele foarte mari de cisteină, probabil mai mare decât 7 g pe zi, pot fi dăunătoare. Pacienții cu cistinurie, o tulburare ereditară caracterizată prin excreția unor cantități mari de cistină și alți aminoacizi în urină, au un risc crescut de formarea calculilor biliari datorita cistina. Ne-ar sugera o limită de 500 mg de două ori pe zi, cisteină, cu excepția cazurilor sub supraveghere medicală. Vitamina C poate preveni toxicitate cisteină. „
Într-adevăr, cisteină trebuie întotdeauna luata cu doze mari de vitamina C si vitaminele B1, B6, E și care îmbunătățesc eficacitatea 9.Alte suplimente recomandate sunt de magneziu și zinc, din care deficiențele sunt în detrimentul metabolismul glutationului.In deficit de magneziu, de exemplu, unul dintre enzimele care este necesară în sinteza glutatione, gamma glutamil transpeptidaza, este coborâta.Suplimente de zinc și magneziu, prin urmare, pot spori sinteza glutationului, în condiții specifice 5.Cu toate acestea, este cunoscut faptul că diabeticii ar trebui sa evite suplimentarea cisteină deoarece poate bloca efectele insulinei prin modificarea structurii sale chimice.Asta este, cisteină rupe unele obligațiuni eco-link SS, schimbarea formei moleculare a insulinei.Unii medici orthomoleculari preferă să descrie N-acetil-cisteină pentru tratamentul SIDA.Acest lucru se datorează faptului că pacienții cu SIDA au, de obicei probleme de absorbție digestive.Este mai ușor pentru ei să ia N-acetil-cisteină decât fie cisteină sau glutation.Medicii care tratează bolnavi de SIDA, în general, recomandă doze zilnice destul de ridicate de N-acetil-cisteină, în intervalul de la 1.800 la 2.400 de miligrame, luate la intervale regulate, în trei sau patru doze divizate 9.
Triptofan
Triptofanul este cel mai puțin abundent aminoacid esențial în produsele alimentare.Deficiențele de aceasta sunt cunoscute a fi legate cu o gamă largă de probleme de sanatate, inclusiv boala Hartnup lui, pelagra, depresie, hipertensiune arteriala, anorexie, insomnie, și comportamentul prea agresiv.Suplimentele de triptofan de până la 3 grame pe zi au fost folosite, de asemenea, pentru a controla durerea greu de rezolvat.Acest aminoacid este utilizat de organism pentru biosinteza de niacina, serotonina, și diferite proteine.Ca urmare, deficiente de triptofan, văzute la persoanele care sunt HIV-seropozitivi, par să rezulte într-o varietate de simptome, inclusiv cele asociate cu pelagra;și anume dermatita, diaree, și demență 5,11
L-triptofan este forma supliment mai de dorit, deoarece toti ceilalti metaboliții triptofan, cu excepția niacina, au efecte secundare semnificative.Braverman și Pfeiffer 5 explică faptul că:
„Infuzii de triptofan pot ridica nivelul triptofan ser de șase până la zece ori la persoanele normale, fara efecte secundare aparente. Încărcare orală (4 g) a controalelor normale poate crește concentrațiile plasmatice de până la patru ori normale în decurs de două ore. Douăsprezece grame zilnic la pacientii maniaci pot menține concentrațiile plasmatice la de trei ori normal „.
Ei descriu în continuare avertizarea : șapte pacienți au luat 2 grame de triptofan pe zi, timp de 6 săptămâni.Nivelurile lor triptofan plasmatice au fost de atunci aproape duble de cele ale unui grup de control de 96 de pacienti.
Spre deosebire de seleniu, cisteina si glutamina, triptofan nu este ușor disponibil în magazine alimentare de sănătate.În toamna anului 1989, FDA a reamintit toate L-triptofan, precizând că a provocat afectiune rara si moartea eozinofilie-mialgie Sindromul (EMS) 12.La 22 martie 1990, FDA a interzis complet vânzarea publică a-L-triptofan.
Adevărul pare a fi că un lot defect de triptofan, probabil, a provocat moartea a 37 de persoane, și permanent cu handicap psete 1.500.Este clar, totuși, că acest lucru s-a datorat nu amino acidului in sine ci contaminarii sale, produs ca urmare a utilizării bacteriilor de inginerie genetica la producerea sa-GMO 13.Interzicerea vânzării de triptofan, din cauza primului dezastru ingineriei genetice din lume, a dus la interzicerea vânzării de whisky din cauza decese cauzate de un lot rau GMO.Cu toate acestea, este încă posibil să cumperati 5-hidroxi triptofan (5-HTP), derivat din semințe de Griffonia Simplicifolia, o plantă medicinală utilizată în mod tradițional în Ghana.CamerunȘi Côte d’Ivoire 14.Acest supliment nu ar trebui să fie luate de către orice persoană care utilizează ISRS (recaptarii serotoninei) sau MAO (monoaminooxidazei) inhibitor medicamentele prescrise de 15.5-hidroxi triptofan este formată prin adăugarea unei grupări hidroxil (OH) la triptofan, de enzimă hidroxilază triptofan și este intermediarul în sinteza naturală de triptofan în serotonină 16.În plus, un alt metabolit triptofan, niacina, este disponibil în orice magazin de produse alimentare de sănătate.De asemenea, trebuie reamintit faptul că există 400 de miligrame de triptofan se gasesc într-o ceașcă de germeni de grâu, în timp ce branza de vaci de grasime redusa conține 300 de miligrame pe ceasca.Există, de asemenea circa 600 de miligrame de triptofan în 1/2 kg de curcan sau de pui.În majoritatea țărilor, în cazul în care nu este disponibil în magazine alimentare de sănătate, triptofan poate fi prescris de către un medic.Într-adevăr, se sugerează cu tărie că oricine încearcă să inverseze deficitele de seleniu, cisteina, glutamina, și triptofan cauzate de infectia cu HIV ar trebui să facă acest lucru sub supravegherea unui medic orthomolecular (nutriționale) .(Nota editorului: Nu menține o bază de date de astfel de medici si recomand o căutare pe internet..-de exemplu Dr Rath are sediu si in Romania)
NU uitati de tratarea CAUZEI(NU doar a simptomelor).Am mentionbat solutii la inceput si intre timp am gasit si ceva interesant ref Vitamina C si HIV/SIDA:
2. Foster, H.D. (2004). How HIV-1 causes AIDS: Implications for prevention and treatment. Medical Hypotheses, 62, 549-553.
3. Shamberger, R., and Willis, C. (1980). Journal of the National Cancer Institute, 44, 931. Cited by Passwater, R.A. (1980). Selenium as food and medicine. New Canaan, CT: Keats, p.18.
4. Shamberger, R. and Willis, C. (1971). CRC critical reviews in clinical laboratory sciences, 211-221. Cited by Passwater, op.cit., pp.21-22.
5. Braverman, E.R. (with Pfeiffer, C.C.)(1987). The healing nutrients within: Facts, findings and new research on amino acids. New Canaan: Keats Publishing.
6. Taylor, E.W. (1997). Selenium and viral diseases: Facts and hypotheses. Journal of Orthomolecular Medicine, 12(4), 227-239.
7. Young, L.C., Gatzen, C., Wilmore, K., and Wilmore, D.W. (1992). Glutamine (Gln) supplementation fails to increase plasma Gln levels in asymptomatic HIV+ individuals. Journal of the American Medical Association, 92 (Suppl), A-88.
8. Robinson, M.K., Hong, R.W., and Wilmore, D.W. (1992). Glutathione deficiency and HIV infection. Letter to the editor, The Lancet, 339, 1603-1604.
9. Pressman, A.H. (with Buff, S.)(1997). The GSH phenomenon: Nature’s most powerful oxidant and healing agent glutathione. New York: St. Martin’s Press, p. 135.
10. Shabert, J., and Ehrlich, N. (1994). The ultimate nutrient glutamine: The essential nonessential amino acid. Garden City Park, NY: Avery Publishing Group, p.46.
11. Fuchs, D., Moller, A.A., Reibnegger, G., Stockle, E., Werner, E.R., and Wachter, H. (1990). Decreased serum tryptophan in patients with HIV-1 infection correlates with increased serum neopterin with neurologic/psychiatric symptoms. Journal of Acquired Immunodeficiency Syndrome, 3(9), 873-876.
12. Manders, D.W. (1995). The FDA ban of L-tryptophan: Politics, profits and Prozac. Science Policy, 26(2). Reprinted on Cognitive Enhancement Research Institute website:http://www.ceri.com/trypto.htm .
13. Boyens, I. (1999). The skeleton in the GMO closet: Did genetic engineering cause the tryptophan-EMS disaster of 1989? From Unnatural harvest: How corporate science is secretly altering our food. Toronto: Doubleday Canada. (Seehttp://www.aquarianonline.com/Eco/UnnaturalHarvest.html)
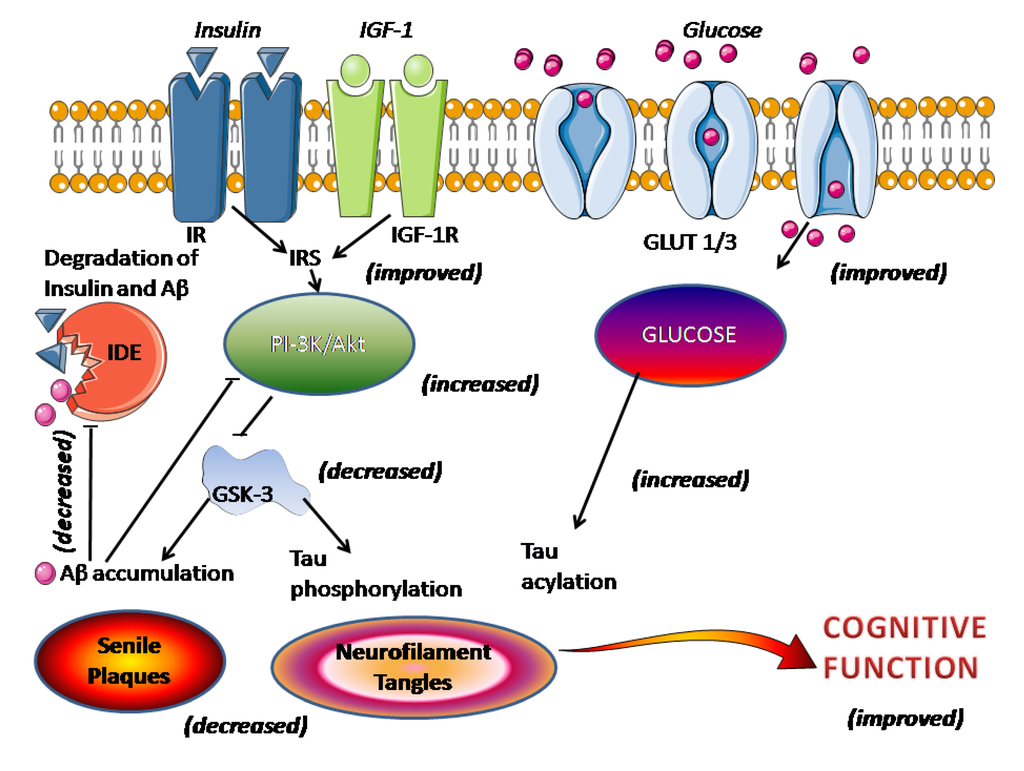
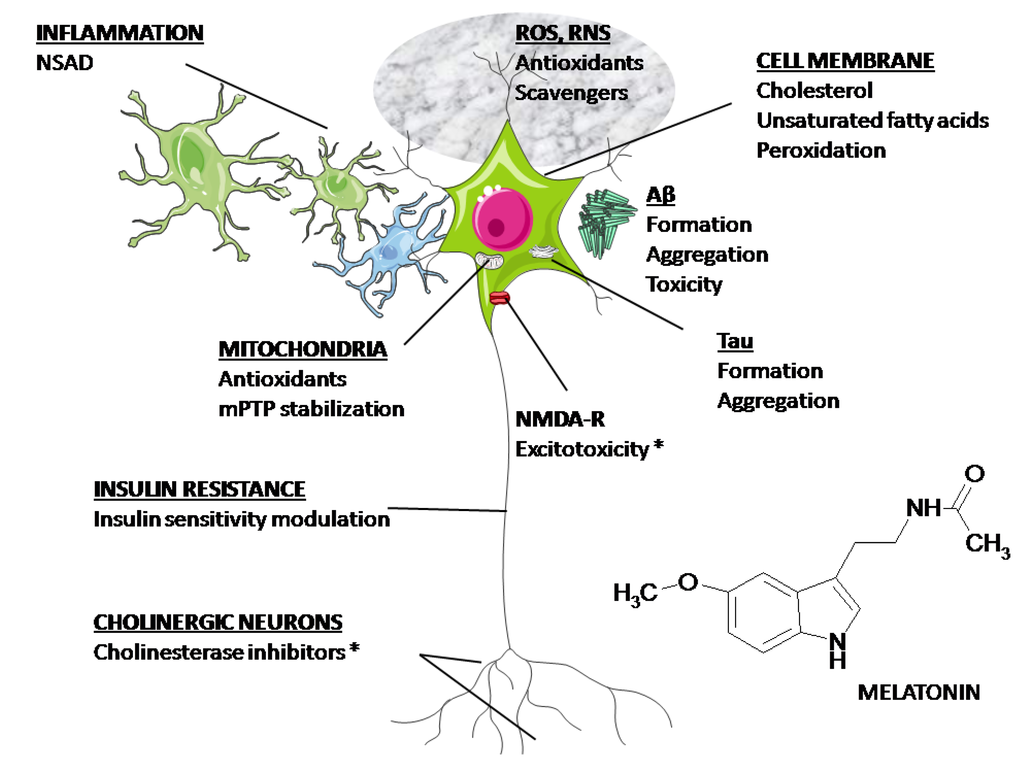














 1,2
1,2


















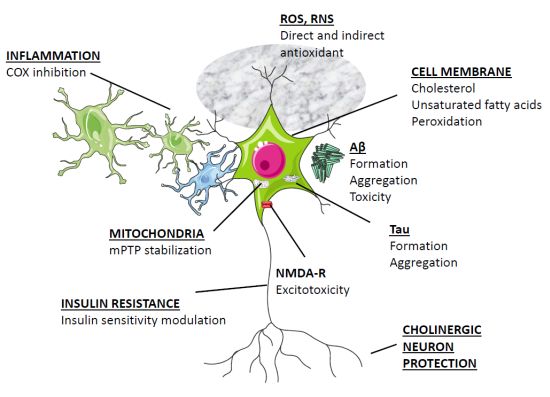{kind=link}
{kind=link}
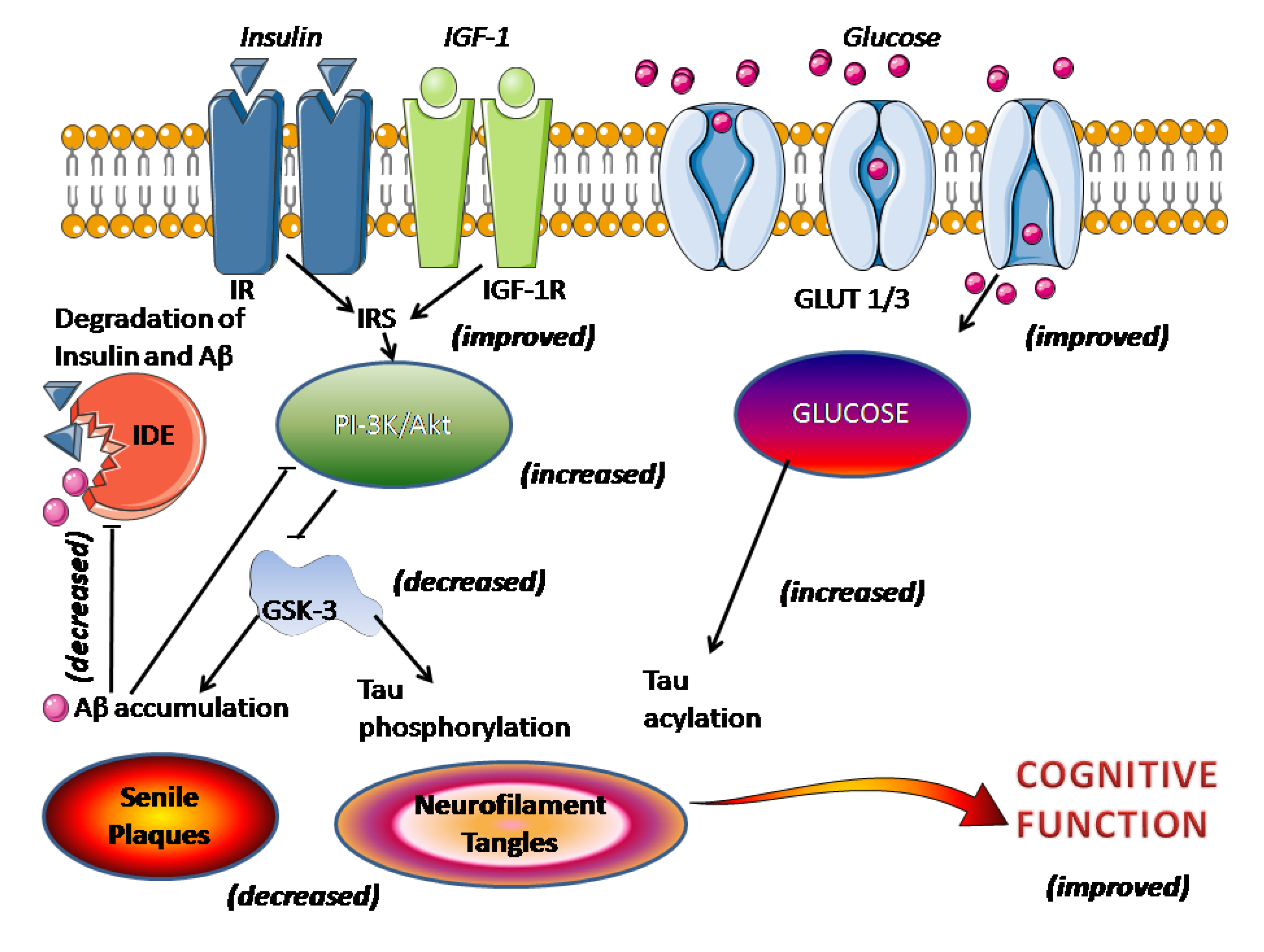{kind=link}
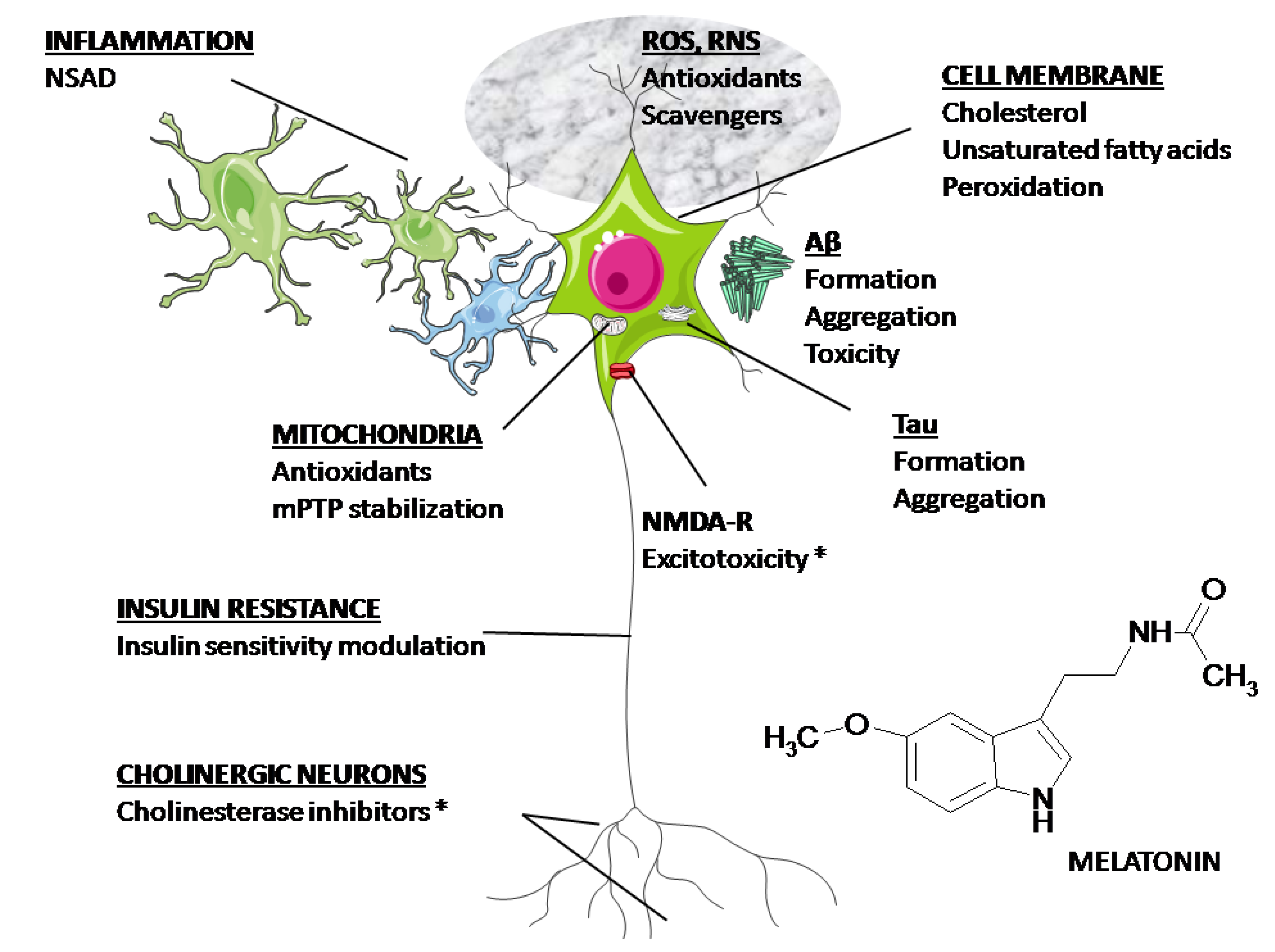{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}