Substanțele fitochimice sunt cele mai valoroase și cuprinzătoare structuri, care pot avea o gamă largă de beneficii protectoare, de la reducerea inflamației și accelerarea vindecării până la prevenirea infecțiilor și combaterea cancerului. Resveratrolul (RSV) este un compus fenolic natural din grupul stilbenoidelor oligomerice, care se găsește de obicei în dieta umană zilnică, cum ar fi strugurii, arahidele, fructele de pădure și cerealele. Prezintă efecte antiinflamatorii, neuroprotectoare, antioxidante și de prevenire și tratament a cancerului. Se crede că RSV are un rezultat impresionant în tratamentul cancerului colorectal (CRC) prin moleculele vitale și căile de semnalizare a cancerului, inclusiv SIRT1, P53, P21, AMPK, ROS, BMP7, COX-2, NO, caspaze, Wnt, TNF, NF -kB, EMT și calea pentozei fosfat. Prin urmare….
Primul studiu clinic concentrat pe eficacitatea RSV (singur și în contextul pulberii de struguri) la pacienții cu cancer de colon a fost efectuat în 2009. Rezultatele au arătat un efect de reglare în jos a RSV asupra CD133 și LGR5. Deși efectul observat nu a fost puternic, a arătat o valoare preventivă pentru aportul alimentar de RSV [117]. Un alt studiu [118]a evaluat siguranța și eficacitatea RSV la subiecții sănătoși, ceea ce a arătat că RSV până la 5 g/zi este sigur dacă este administrat oral. În plus, a fost observată o scădere a nivelurilor de IGF-1 și IGFBP-3 la unele tipuri de cancer. Într-un alt studiu, a fost analizată concentrația de resveratrol și metaboliții săi în țesutul colorectal la pacienții cu cancer colorectal. Evaluarea țesuturilor de biopsie a demonstrat că RSV și metaboliții săi au fost de preferință acumulați în partea dreaptă a colonului. Această selectivitate a site-ului este compara rabil la aspirină, care scade în mod favorabil riscul de tumori colorectale pe partea dreaptă. Acest studiu a demonstrat o inhibare de 5% a proliferării celulelor tumorale după aportul zilnic de RSV [119, 120]
Mai mult, un studiu care implică o nouă formulare micronizată (SRT501) a RSV a arătat o creștere substanțială a nivelurilor unui marker de apoptoză, numit caspază-3 scindată, în rândul pacienților cu cancer colorectal cu metastaze hepatic de asemenea, forma de dozare SRT501 oferă o biodisponibilitate mai bună pentru RSV și este bine tolerată de către pacienți [119]. În plus, există și alte studii clinice axate pe RVS și care implică pacienți cu cancer colorectal, care nu au fost publicate, dar înregistrate la http://www.clinicaltrials.gov [94]
Studiile clinice și preclinice menționate asupra RSV (în cancerul colorectal și alte tipuri de cancer) au fost revizuite în detaliu de Ko et al.[3].
Cancerul colorectal (CRC) este al treilea cel mai frecvent cancer și una dintre principalele cauze de deces prin cancer la nivel mondial. Factorii de mediu, alimentația și stilul de viață, inclusiv consumul de carne roșie, fumatul de țigară, consumul de alcool și antecedentele familiale sunt cei mai importanți factori de risc ai CCR. Mai multe căi, inclusiv inflamația, stresul oxidativ și apoptoza sunt implicate în incidența și progresia acesteia. Resveratrolul, un compus polifenolic, are diferite funcții farmacologice, inclusiv antiinflamație, prevenirea cancerului, efect de scădere a lipidelor și efect hipoglicemiant. Multe studii au demonstrat că resveratrolul ar putea reprezenta, de asemenea, un efect chimio-preventiv asupra CCR. Astfel, scopul revizuirii actuale este de a descrie rolul resveratrolului în tratamentul CCR într-o manieră moleculară.
Cancerul colorectal (CRC) este al treilea cel mai frecvent cancer la nivel mondial [ 1 ]. Metastazele hepatice sunt cauza principală de mortalitate în acest cancer [ 2 ]. CRC a provocat aproximativ 0,69 milioane de decese, iar rata mortalității este încă în creștere. Statistica internațională a cancerului a indicat că există 1,36 milioane de cazuri noi de CRC anual în întreaga lume [ 3 ]. În țările occidentale, în special în Statele Unite, există o scădere recentă a ratei de incidență a CRC, care se datorează screening-ului crescut, cum ar fi colonoscopie și test de sânge ocult în fecale [ 4 ]. Screening-ul îmbunătățit contribuie la detectarea precoce și tratamentul polipilor precancerosi [ 5 , 6 ]] ca și în țările cu venituri mari, prevalența bolii a fost stabilă din cauza screening-ului crescut [ 7 ]. Tratamentul CRC reprezintă încă o problemă clinică, în timp ce ratele de supraviețuire la 5 ani sunt de aproximativ 65%, ceea ce depinde de situația tumorii, stadiul de detectare a tumorii și terapie [ 8 ]. Studiile de tendințe seculare au sugerat că stilul de viață, dieta și factorii de mediu, inclusiv consumul de carne roșie, fumatul de țigară și consumul de alcool sunt cei mai importanți factori de risc ai CCR [ 5 , 9 ]. În plus, mai multe studii au arătat că antecedentele familiale de CCR au crescut riscul de îmbolnăvire. Pe baza observațiilor anterioare, ghidurile prezente propun ca persoanele cu antecedente familiale de acest cancer să fie testate pentru CCR mai devreme decât cele fără un astfel de cancer.10 ]. Recunoașterea noilor strategii terapeutice poate crește rata de supraviețuire a pacienților cu CCR [ 11 ].
Alături de diferite terapii, utilizarea compușilor naturali a apărut ca un nou orizont în tratamentul unei varietăți de boli precum cancerul [ 12 – 14 ]. În acest sens, mai multe investigații in vitro și in vivo au arătat că fitochimicele au rol potențial antioxidant, antiinflamator și anticancerigen prin modularea anumitor căi de semnalizare și biomarkeri moleculari pentru a opri incidența și progresia CRC [ 15 ]. Resveratrolul (trans-3, 4′, 5-trihidroxistilben) care derivă din familia stilbenelor de compuși fenolici, există în fructe de pădure, pin și nuci și în special în coaja strugurilor roșii [ 16 ].]. Acest compus are diferite funcții farmacologice, inclusiv antiinflamație, inhibare a cancerului, efect de scădere a lipidelor și efect hipoglicemiant. Resveratrolul inhibă peroxidarea lipidelor și agregarea trombocitelor, astfel joacă ca un antioxidant, antiinflamator și vasodilatator [ 17 , 18 ]. Recent, s-a descoperit că resveratrolul instigă autofagia în celulele canceroase [ 19 ]. S-a demonstrat că resveratrolul crește eliberarea diferitelor citokine proinflamatorii din celulele imune, care în cele din urmă promovează citotoxicitatea împotriva cancerului [ 20 ]. Acest agent natural sigur și multi-țintit a fost conectat cu suprimarea supraviețuirii și invazia celulelor canceroase [ 21 ].
Multe studii au demonstrat că resveratrolul ar putea reprezenta, de asemenea, un efect chimio-preventiv asupra CCR. Un studiu recent in vitro a demonstrat că resveratrolul în combinație cu 5-fluorouracil (5-FU), un medicament chimioterapeutic, a crescut efectele 5-FU prin efectele sale ca medicament anti-metastază asupra CRC. A fost evident faptul că celulele rezistente la 5-FU au fost mai susceptibile la resveratrol, ilustrând o potențială strategie terapeutică pentru CRC rezistent la 5-FU [ 21 ]. Studiile anterioare au raportat, de asemenea, că resveratrolul crește activitățile anticanceroase ale agentului chimioterapeutic, oxaliplatina, în modelul de cultură celulară al CRC. Resveratrolul poate crește sinergic efectele oxaliplatinei în suprimarea creșterii celulelor tumorale [ 22 ].
Având în vedere că resveratrolul nu este considerat ca terapie primară în tratamentul cancerului [ 23 ]. Dovezile din mai multe linii au indicat că resveratrolul ar putea fi folosit ca linie secundară de terapie și terapiile primare pentru cancer alături de resveratrol arată rezultate semnificative [ 23 , 24 ]. În acest sens, o gamă largă de studii au confirmat că resveratrolul cu abordări radioterapice sau chimioterapii ar putea fi oferit un regim terapeutic bun în tratamentul pacienților cu cancer [ 24 , 25 ].
Studiile cinetice metabolice au arătat că resveratrolul are o rată mare de clearance în circulație și o biodisponibilitate scăzută [ 26-28 ] , astfel încât aplicarea sa pentru inhibarea CRC a fost foarte contestată . Unele studii au recomandat că motivul posibil al cantității scăzute de resveratrol în circulație este că resveratrolul induce enzimele metabolice de fază II UGT și sulfotransferaza cât mai curând posibil pentru a genera glucuronide de resveratrol și sulfați de resveratrol [ 29 , 30 ].]. Un studiu clinic recent a clarificat faptul că nivelul a trei conjugați de sulfat de resveratrol a fost crescut la pacienții cu absorbție orală de 0,5-1,0 g resveratrol în fiecare zi și progresia celulelor tumorale a fost remarcabil inhibată la aceștia, propunând ca administrarea orală zilnică de 0,5-1,0 g. g resveratrolul ar putea avea un rol potențial în inhibarea CRC [ 31 ]. Astfel, scopul revizuirii actuale este de a descrie rolul resveratrolului în tratamentul CCR într-o manieră moleculară.
Inflamația cronică este unul dintre principalele mecanisme implicate în cancerul de colon. Prin urmare, compușii antiinflamatori pot fi benefici în tratamentul CRC. Citokinele sunt eliberate rapid de țesuturile deteriorate și sunt stimulatori ai răspunsului inflamator [ 32 , 33 ]. Foarte recent studii au indicat că expunerea celulelor intestinale la citokine poate mobiliza căi inflamatorii precum MAPK-urile, JAK-STAT și cascadele NF-kB, poate crește expresia enzimelor proinflamatorii, poate induce generarea de mediatori proinflamatori și, de asemenea, producerea de specii reactive de oxigen (ROS) [ 34 , 35]. Resveratrolul scade mediatorii proinflamatori, cum ar fi TNF-α și IL-1β, enzimele proinflamatorii precum iNOS și COX-2 și căile de semnalizare inflamatorii, cum ar fi NF-kB (Fig. 1) [ 36 ].
Reprezentare schematică în țintirea diferitelor căi de semnalizare folosind resveratrolul ca o nouă strategie terapeutică în tratamentul cancerului colorectal
Metastaza și invazia celulelor canceroase sunt procese interconectate care implică proliferarea celulelor, aderența celulară, migrarea celulelor și degenerarea proteolitică a barierelor tisulare, cum ar fi ECM și membrana bazală. Mai multe enzime proteolitice care includ metaloproteinaze de matrice (MMP-uri; în special MMP-2 și MMP-9) [ 37 , 38 ] și molecule de adeziune intracelulară (ICAM; în special ICAM-1), participă la degenerarea acestor bariere [ 39 , 40 ] . Unele dintre studiile în cancerul pancreatic, mamar, pulmonar și de colon au evidențiat supraexprimarea MMP-urilor în țesuturile virulente în comparație cu țesuturile aproape normale [ 41 , 42 ].]. Resveratrolul a diminuat migrarea și invazia celulelor canceroase pulmonare datorită suprimării activării NF-κB și a exprimării MMP-2 și MMP-9 [ 43 ]. Studiile anterioare au demonstrat că resveratrolul funcționează ca un factor multi-tasking și are funcții antiinflamatorii și anticancerigene în CRC [ 44 , 45 ]. Studii recente au demonstrat că resveratrolul inhibă mecanismele de semnalizare dependente de NF-κB [ 15 ]. Diferiți mediatori proinflamatori care s-au demonstrat că modifică micromediul tumoral, inclusiv membrii superfamiliei factorului de necroză tumorală (TNF), sunt modulați de NF-kB [ 9 , 10 ].]. TNF-α care este generat în micromediul tumoral, modulează celulele canceroase, celulele stromale din jur și ECM în mai multe tipuri de cancer și acționează ca un factor de creștere autocrin și paracrin [ 46 , 47 ]. Un studiu recent a raportat că resveratrolul a suprimat invazia și viabilitatea celulelor CRC induse de TNF-α și TNF-β [ 8 ]. Un alt studiu a raportat că resveratrolul a inhibat supraviețuirea îmbunătățită cu TNF-β și TNF-α în celulele CRC prin creșterea factorilor apoptotici, cum ar fi caspaza-3 scindată [ 48 ].]. Acest studiu a mai arătat că resveratrolul a scăzut activarea NF-kB, produsele genetice cancerigene dependente de NF-kB (MMP-9 și CXCR4), precum și factorii de semnalizare legați de EMT, inclusiv vimentina, melcul și E-cadherina în celulele CRC. Există, de asemenea, unele dovezi care au arătat că resveratrolul poate inhiba căile de semnalizare a MMP-9 și a factorului de creștere endotelial vascular (VEGF). Suprimarea MMP-9 și VEGF de către resveratrol previne metastazele și angiogeneza [ 49 , 50 ]. Oxidul nitric (NO) este produs de trei izoenzime ale NO sintazei. Unul dintre aceste izosime este iNOS, care este inductibil de citokine și mediatori proinflamatori și este independent de calciu, în timp ce altele sunt dependente de calciu/calmodulină și sunt produse în mod constitutiv [ 51 ].]. În modelele umane și animale de CRC, s-a indicat că activarea și expresia iNOS este crescută, sugerând rolul semnificativ al NO în tumorigeneza colonului [ 52 ]. Mai multe studii au raportat, de asemenea, că resveratrolul a scăzut expresia iNOS în celulele canceroase de colon, totuși mecanismul său principal este încă neclar [ 53 ]. Pterostilbenul (trans-3, 5-dimetoxi-4′-hidroxistilben), un analog structural al resveratrolului, exercită o acțiune antiinflamatoare pentru inhibarea carcinogenezei de colon care inhibă calea de semnalizare a p38 MAPK care duce la inducerea COX-2 și iNOS [ 35 ]]. În plus, resveratrolul este o suplimentare profitabilă, netoxică și o strategie alternativă pentru a reduce colita și cancerul de colon puternic legat de colită. Resveratrolul dezvoltă în mod remarcabil scorul de inflamație, scade cantitatea de neutrofile în ganglionii limfatici mezenterici și lamina propria și reglează celulele T CD3(+) care eliberează TNF-α și IFN-γ. P53, un marker al inflamației, este, de asemenea, diminuat de resveratrol [ 54 ].
În prezent, stresul oxidativ a crescut, presupus ca un amplificator principal al carcinogenezei [ 55 ]. Mai multe studii s-au concentrat pe exercitarea chimio-preventivă a resveratrolului ca antioxidant împotriva îmbătrânirii, a bolilor cardiovasculare și a cancerului. Activitatea antioxidantă a resveratrolului este asociată în principal cu capacitatea sa de a inhiba producția de radicali liberi, peroxidarea lipidelor și modularea enzimelor asociate cu antioxidanti [ 56 ]. Una dintre enzimele antioxidante importante este mangan superoxid dismutaza, care transformă radicalii liberi dăunători, superoxidul, în oxigen molecular și peroxid de hidrogen [ 49 ]. Resveratrolul suprimă expresia acestei enzime și, prin urmare, contribuie la inhibarea ROS [ 56]. Resveratrolul a reprezentat deja efecte supresoare asupra oxidării proteinelor și lipidelor mediate de peroxinitrit [ 57 ] prin reglarea în sus a SOD [ 58 ], catalazei (CAT), glutation peroxidazei (GPX) [ 59 ] și activarea căilor Sirt1/AMPK și Nrf2. [ 60 ].
Două molecule de semnalizare substanțiale, factorul de replicare STAT3 și protein kinaza AKT, au fost raportate a fi supra-exprimate sau activate în majoritatea tipurilor de cancere umane, prin urmare, acum este de obicei aprobat că ambele proteine prezintă obiective de încredere pentru noile antitumorale. proiectarea medicamentelor [ 61 , 62 ]. S-a descoperit că liniile celulare CRC reprezintă activarea fundamentală a proteinelor de creștere a supraviețuirii, inclusiv AKT și STAT3. Resveratrolul inhibă proteinele AKT și STAT3 despre care se recunoaște că au capacitate oncogenă în CRC [ 63 ]. Wallerath şi colab. [ 64] a demonstrat că resveratrolul ar putea crește rapid generarea de NO în celulele endoteliale cultivate. La niveluri fiziologice, NO, ca antioxidant, susține mucoasa gastrointestinală împotriva leziunilor, previne lipirea leucocitelor și protejează fluxul sanguin al mucoasei [ 65 , 66 ].
În carcinoame, s-a descoperit că semnalizarea mediată de integrină prin adeziune focală asigură reorganizarea citoscheletului și susține dezvoltarea tumorii, invazia și metastaza [ 67 ]. Unele studii au demonstrat că acumularea Focal Adhesion Kinaze (FAK) cu integrine și proteine citoscheletice în contactele focale cauzează o migrație crescută a celulelor, precum și o potențială modulare a creșterii și supraviețuirii celulelor [ 68 , 69 ]. De asemenea, în tumori, s-a demonstrat că FAK este implicat în migrarea celulelor, supraviețuirea, invazia și metastaza, expresia genelor și auto-reînnoirea celulelor stem tumorale [ 70 ].]. Resveratrolul suprimă proliferarea și invazia celulelor CRC prin inducerea activității Sirt1, reducerea căii inflamatorii mediate de NF-κB și inhibarea activității kinazei de adeziune focală (FAK), ceea ce duce la scăderea moleculelor de aderență focală și la o creștere a apoptozei [ 71 ]. În plus, resveratrolul a inhibat capacitatea de invazie și formare de colonii, creșterea celulelor, expresia β1-integrinei și activarea FAK a celulelor în micromediul cancerului de alginat. Resveratrolul a suprimat activarea NF-kB și, prin urmare, a inhibat produsele finale ale genei dependente de NF-kB care sunt implicate în apoptoză, metastază și invazie [ 71 ].
Un subgrup de superfamilie a factorului de creștere transformantă (TGF)-β, proteinele morfogenetice osoase (BMP), joacă un rol important în reglarea mai multor faze cheie ale dezvoltării embrionare, creșterii, diferențierii și apoptozei diferitelor celule [ 72 ]. Dovezi recente au indicat că BMP-urile participă la progresia cancerului, cum ar fi cancerul de colon [ 73 ]. BMP își începe semnalizarea prin calea fosfatidil inozitol 3-kinaza (PI3K)/Akt [ 74 , 75 ]. Semnalizarea P13K/Akt joacă un rol serios în reglarea supraviețuirii celulare și a apoptozei [ 76 ].] care a fost detectat supraactivat în multe tumori pentru a scădea apoptoza și a promova proliferarea. Inhibitorul tumoral fosfataza și omologul tensinei (PTEN) reglează în jos semnalizarea PI3K/Akt și este adesea inactivat de mutații în mai multe tipuri de cancer. Resveratrolul reduce fosforilarea Akt1/2/3 în mod semnificativ în celulele CRC [ 77 ].
Resveratrolul a reprezentat, de asemenea, suprimarea dependentă de doză a căii de semnalizare Wnt, care este una dintre căile importante în mai multe boli grave, cum ar fi cancerul. Resveratrolul scade expresia genelor țintă Wnt, inclusiv ciclina D1 și conducție, și suprimă dezvoltarea celulelor induse de Wnt și a celulelor CRC conduse de Wnt [ 78 ]. Alte studii au raportat că resveratrolul scade expresia ciclinelor D1 și D2, care reglează direct progresia ciclului celular [ 79 ]. Aceste molecule sunt de obicei induse în timpul malignității [ 80 ] și inhibate de fitochimicale anticancer [ 81 ]. De asemenea, oprirea ciclului celular de către resveratrol într-o varietate de tipuri de linii celulare canceroase este bine înregistrată [ 82 ].]. În plus, resveratrolul suprimă activitățile diferitelor enzime legate de replicarea ADN-ului și proliferarea celulară [ 83 ]; aceste efecte colective ar putea fi motivul acceptabil pentru efectele resveratrolului asupra modulării proliferării celulare [ 84 ].
În ciuda tuturor caracteristicilor anticanceroase documentate ale resveratrolului, majoritatea investigațiilor au fost efectuate în metode preclinice și de cultură celulară. Deoarece există diferențe în profilul metabolismului și genetica oamenilor și, de asemenea, pentru că proprietățile potențiale ale resveratrolului în modelele animale nu pot fi presupuse egale cu oamenii, efectele fiziologice ale resveratrolului au fost studiate și la oameni.
Toxicitatea, metabolismul și farmacocinetica resveratrolului au fost evaluate la pacienții cu cancer și la subiecții sănătoși [ 85 , 86 ]. Resveratrolul este metabolizat rapid, mai ales în sulfat conjugați și glucuronide care sunt excretate prin urină. Datorită biodisponibilității scăzute din cauza metabolismului său extins, doze mari de resveratrol (până la 5 g/zi) au fost utilizate de către investigatori. Cercetările menționate au arătat că acest compus polifenolic pare să fie sigur și bine tolerat. Cu toate acestea, efecte adverse, cum ar fi dureri abdominale, greață și diaree, au fost observate la persoanele care consumă mai mult de 1 g resveratrol pe zi [ 86 ]. Astfel, studiile clinice studiază această limitare a dozei [ 86 , 87]. Biodisponibilitatea slabă a resveratrolului este o problemă importantă în ceea ce privește extrapolarea impactului său asupra oamenilor și au fost dezvoltate diferite metode pentru a crește biodisponibilitatea acestuia, cum ar fi administrarea acestuia cu diferite alimente, utilizarea în combinație cu o piperină fitochimică suplimentară, precum și utilizarea acestuia. o formulă nanotehnologică, pulberi micronizate sau abordare promedicament [ 88 – 93 ]. Masa 1arată lista de studii clinice pe resveratrol și diferite tipuri de cancer, inclusiv CRC.
tabelul 1
Studii clinice despre resveratrol și cancer
Cancer
Doza de resveratrol
Durata studiului
Nr pacienti
Rezultat
Refs
CRC
5 g/zi
14 zile
6
Creșteți caspaza-3 scindată în țesutul hepatic malign.
Profil de siguranță inacceptabil și eficacitate minimă la pacienții cu mielom multiplu recidivat/refractar, evidențiind riscurile dezvoltării de noi medicamente în astfel de populații
Cancer colorectal CRC , cancer de prostată PC , mielom multiplu MM , cancer de sân BC
O investigație efectuată asupra patogenezei cancerului de prostată a arătat că resveratrolul este capabil să întârzie reapariția cancerului. Aproximativ 33-50% dintre pacienții cu cancer de prostată prezintă reapariția bolii biochimice după terapia primară. Creșterea concentrației de antigen prostatic specific (PSA) este cea mai precoce indicație de recidivă a bolii. MPX, piele de struguri de muscadină pulverizată care conține resveratrol, a întârziat dezvoltarea recurenței prin prelungirea timpului de dublare a PSA (PSADT) cu 5,3 luni. Cu toate acestea, aceste constatări nu au fost semnificative [ 94]. Această investigație se desfășoară în studiul clinic de fază II, așa că rămâne de observat dacă MPX este o opțiune viabilă de terapie. În mod contradictoriu, un al doilea studiu clinic care implică resveratrolul și cancerul de prostată a concluzionat cu siguranță că nu ar fi o terapie viabilă. În ciuda datelor preclinice conform cărora resveratrolul reglează activitatea receptorului de androgeni și scade generarea de androgeni, Kjaer și colab. [ 95 ] a indicat că, deoarece nu a avut niciun impact asupra nivelurilor de PSA sau a volumului prostatei, resveratrolul nu ar putea trata cancerul de prostată. Pe baza celor discutate, pare improbabil ca resveratrolul să demonstreze că este eficient pentru terapia cancerului de prostată, dar sunt necesare studii clinice suplimentare pentru a confirma acest lucru.
Există și alte cercetări care arată că resveratrolul este o terapie slabă, inclusiv în anumite tipuri de mielom multiplu (MM). S-a observat că resveratrolul suprimă STAT3, AKT și NF-kB și exercită efecte citotoxice în liniile celulare MM [ 96 ]. SRT501 a fost examinat la subiecții cu MM refractar sau recidivat. În ciuda dovezilor convingătoare ale studiilor preclinice că resveratrolul contribuie la terapia pacienților cu MM, studiul clinic a arătat că a provocat numeroase efecte adverse severe, dintre care cea mai proeminentă a fost insuficiența renală [ 97 ].]. Deoarece SRT501 nu a avut efecte nefrotoxice într-un studiu de fază I și a fost documentat că este sigur într-un studiu de fază II pentru pacienții cu cancer colorectal, aceste efecte adverse părea să fie specifice MM. Aceste descoperiri arată că SRT501 și probabil orice alte versiuni de resveratrol nu ar putea fi o terapie potențială pentru MM.
De asemenea, în studiile clinice efectuate la pacienții cu cancer colorectal, constatările par pline de speranță, dar rămân nedeterminate în ceea ce privește dacă resveratrolul ar putea fi o terapie adecvată. S-a descoperit că resveratrolul suprimă creșterea tumorii și declanșează apoptoza în celulele colonului uman in vitro, iar modelele murine au indicat că resveratrolul suprimă carcinogeneza și inflamația colorectale [ 98 , 99 ].]. Astfel, două studii clinice au urmărit să distingă farmacocinetica resveratrolului în metastazele hepatice sau în țesutul colorectal. După o terapie de 2 săptămâni cu resveratrol sau SRT501, în țesuturile de cancer colorectal ale pacienților, resveratrolul parental măsurat, precum și nivelurile principale ale metaboliților săi au fost similare cu nivelurile eficiente de resveratrol utilizate în investigațiile preclinice. Cu toate acestea, funcția anticanceroasă a metaboliților resveratrolului nu a fost încă validată, prin urmare nu este clar dacă acest rezultat oferă o justificare suplimentară pentru urmărirea resveratrolului ca potențială terapie pentru cancerul de colon. În plus, nivelurile de caspază-3, un biomarker apoptotic și nivelurile de Ki-67, un biomarker al proliferării, au fost ușor influențate în probele de țesut [ 31 , 88 ].]. Deși este dovedit că resveratrolul posedă unele efecte farmacologice, este obscur dacă aceste efecte sunt suficient de considerabile pentru a face din el un material terapeutic benefic pentru tratamentul cancerului de colon.
CRC este un cancer predominant și una dintre principalele cauze de mortalitate prin cancer în întreaga lume. Mai mulți factori de la genetică la dietă sunt implicați în incidența acestei malignități. Fiziopatologia sa este eterogenă, în care moleculele multiple și diferite căi de semnalizare, inclusiv inflamația, stresul oxidativ și apoptoza sunt implicate în incidența și progresia sa. O serie de studii au susținut efectele potențiale ale resveratrolului în tratamentul CRC. Acest compus polifenol prezintă diferite proprietăți, inclusiv eficacitate antioxidantă, antiinflamatoare, inductor de apoptoză și anti-angiogeneză. Datorită acestor efecte semnificative, resveratrolul este sugerat ca un nou agent terapeutic pentru cancer. În plus, unele studii au raportat că consumul de resveratrol în combinație cu alte medicamente anticancerigene le poate crește efectele și, de asemenea, le poate reduce efectele secundare. Astfel, acest compus multi-tasking poate fi un nou candidat în tratamentul CRC, cu toate acestea, sunt necesare mai multe studii umane.
ZA a contribuit la conceperea, proiectarea și redactarea manuscrisului. MH, RS, HM și RR. J a contribuit la colectarea datelor și la redactarea manuscrisului. ZA a supravegheat studiul. Toți autorii au citit și au aprobat manuscrisul final.
Reijonen P, Osterlund P, Isoniemi H, Arola J, Nordin A. Invazia biliară verificată histologic a fost asociată cu supraviețuirea fără recurență hepatică afectată în metastazele hepatice ale cancerului colorectal rezecat. Scand J Surg. 2018 doi: 10.1177/1457496918812237. [ PubMed ] [ CrossRef ] [ Google Scholar ]2.
Helling TS, Martin M. Cauza morții prin metastaze hepatice în cancerul colorectal. Ann Surg Oncol. 2014; 21 :501–506. doi: 10.1245/s10434-013-3297-7. [ PubMed ] [ CrossRef ] [ Google Scholar ]3.
Gan Y, Li Y, Li T, Shu G, Yin G. CCNA2 acționează ca un biomarker nou în reglarea creșterii și apoptozei cancerului colorectal. Cancer Manag Res. 2018; 10 :5113–5124. doi: 10.2147/CMAR.S176833. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]4.
Fajardo AM, Piazza GA. Chimioprevenția în fiziologia și bolile gastrointestinale. Abordări antiinflamatorii pentru chimioprevenția cancerului colorectal. Am J Physiol Gastrointest Ficat Physiol. 2015; 309 :G59–G70. doi: 10.1152/ajpgi.00101.2014. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]5.
Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011; 61 :69–90. doi: 10.3322/caac.20107. [ PubMed ] [ CrossRef ] [ Google Scholar ]6.
Siegel R, Naishadham D, Jemal A. Statistica cancerului, 2013. CA Cancer J Clin. 2013; 63 :11–30. doi: 10.3322/caac.21166. [ PubMed ] [ CrossRef ] [ Google Scholar ]7.
Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015; 65 :87–108. doi: 10.3322/caac.21262. [ PubMed ] [ CrossRef ] [ Google Scholar ]8.
Siegel RL, Miller KD, Fedewa SA, Ahnen DJ, Meester RGS, Barzi A, et al. Statistica cancerului colorectal, 2017. CA Cancer J Clin. 2017; 67 :177–193. doi: 10.3322/caac.21395. [ PubMed ] [ CrossRef ] [ Google Scholar ]9.
Aune D, Chan DS, Lau R, Vieira R, Greenwood DC, Kampman E, Norat T. Fibre alimentare, cereale integrale și risc de cancer colorectal: revizuire sistematică și meta-analiză doză-răspuns a studiilor prospective. BMJ. 2011; 343 :d6617. doi: 10.1136/bmj.d6617. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]10.
Park CH, Kim NH, Park JH, Park DI, Sohn CI, Jung YS. Impactul antecedentelor familiale de cancer colorectal asupra prevalenței specifice vârstei a neoplaziei colorectale. J Gastroenterol Hepatol. 2019; 34 :537–543. [ PubMed ] [ Google Scholar ]11.
Kim DU, Kwak B, Kim SW. Fosfodiesteraza 4B este o țintă terapeutică eficientă în cancerul colorectal. Biochem Biophys Res Commun. 2019; 508 :825–831. doi: 10.1016/j.bbrc.2018.12.004. [ PubMed ] [ CrossRef ] [ Google Scholar ]12.
Muhammad N, Steele R, Isbell TS, Philips N, Ray RB. Extractul de pepene amar inhibă creșterea cancerului de sân în modelul preclinic prin inducerea morții celulelor autofagice. Oncotarget. 2017; 8 (39):66226–66236. doi: 10.18632/oncotarget.19887. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]13.
Bhattacharya S, Muhammad N, Steele R, Kornbluth J, Ray RB. Pepenele galben crește toxicitatea mediată de ucigașul natural împotriva celulelor canceroase ale capului și gâtului. Cancer Prev Res. 2017; 10 :337–344. doi: 10.1158/1940-6207.CAPR-17-0046. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]14.
Bhattacharya S, Muhammad N, Steele R, Peng G, Ray RB. Rolul imunomodulator al extractului de pepene amar în inhibarea creșterii carcinomului cu celule scuamoase la cap și gât. Oncotarget. 2016; 7 :33202–33209. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]15.
Yin TF, Wang M, Qing Y, Lin YM, Wu D. Progresul cercetării privind efectele chimiopreventive ale fitochimicelor asupra cancerului colorectal și a mecanismelor acestora. World J Gastroenterol. 2016; 22 :7058–7068. doi: 10.3748/wjg.v22.i31.7058. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]16.
Rotelli MT, Bocale D, De Fazio M, Ancona P, Scalera I, Memeo R, et al. Dovezi IN-VITRO pentru proprietățile protectoare ale principalelor componente ale dietei mediteraneene împotriva cancerului colorectal: o revizuire sistematică. Surg Oncol. 2015; 24 :145–152. doi: 10.1016/j.suronc.2015.08.001. [ PubMed ] [ CrossRef ] [ Google Scholar ]17.
Pace-Asciak CR, Hahn S, Diamandis EP, Soleas G, Goldberg DM. Fenolicii din vinul roșu trans-resveratrolul și quercetina blochează agregarea trombocitelor umane și sinteza eicosanoidelor: implicații pentru protecția împotriva bolii coronariene. Clin Chim Acta. 1995; 235 :207–219. doi: 10.1016/0009-8981(95)06045-1. [ PubMed ] [ CrossRef ] [ Google Scholar ]18.
Shankar S, Singh G, Srivastava RK. Chimioprevenirea prin resveratrol: mecanisme moleculare și potențial terapeutic. Front Biosci. 2007; 12 :4839–4854. doi: 10.2741/2432. [ PubMed ] [ CrossRef ] [ Google Scholar ]19.
Wang J, Li J, Cao N, Li Z, Han J, Li L. Resveratrolul, un activator al SIRT1, induce autofagie protectoare în cancerul pulmonar fără celule mici prin inhibarea Akt/mTOR și activarea p38-MAPK. Onco vizează acolo. 2018; 11 :7777–7786. doi: 10.2147/OTT.S159095. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]20.
Trung LQ, An DTT. Este resveratrolul o moleculă imunomodulatoare a cancerului? Front Pharmacol. 2018; 9 :1255. doi: 10.3389/fphar.2018.01255. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]21.
Buhrmann C, Yazdi M, Popper B, Shayan P, Goel A, Aggarwal BB și colab. Resveratrolul chemosensibilizează supraviețuirea indusă de tnf-beta a celulelor de cancer colorectal tratate cu 5-FU. Nutrienți. 2018 doi: 10.3390/nu10070888. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]22.
Kaminski BM, Weigert A, Scherzberg MC, Ley S, Gilbert B, Brecht K, et al. Potențarea indusă de resveratrol a efectelor antitumorale ale oxaliplatinei este însoțită de un profil de citokine modificat al macrofagelor umane derivate din monocite. Apoptoza. 2014; 19 :1136–1147. doi: 10.1007/s10495-014-0988-x. [ PubMed ] [ CrossRef ] [ Google Scholar ]23.
Carter LG, D’Orazio JA, Pearson KJ. Resveratrol și cancer: concentrare pe dovezi in vivo. Endocr Relat Cancer. 2014; 21 (3):R209–R225. doi: 10.1530/ERC-13-0171. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]24.
Jiang Z, Chen K, Cheng L, Yan B, Qian W, Cao J și colab. Resveratrol și tratamentul cancerului: actualizări. Ann NY Acad Sci. 2017; 1403 :59–69. doi: 10.1111/nyas.13466. [ PubMed ] [ CrossRef ] [ Google Scholar ]25.
Kma L. Efectul sinergic al resveratrolului și radioterapiei în controlul cancerelor. Asian Pac J Cancer Prev. 2013; 14 :6197–6208. doi: 10.7314/APJCP.2013.14.11.6197. [ PubMed ] [ CrossRef ] [ Google Scholar ]26.
Baur JA, Sinclair DA. Potențialul terapeutic al resveratrolului: dovezile in vivo. Nat Rev Drug Discov. 2006; 5 :493–506. doi: 10.1038/nrd2060. [ PubMed ] [ CrossRef ] [ Google Scholar ]27.
Gescher AJ, Steward WP. Relația dintre mecanisme, biodisponibilitate și eficacitatea chimiopreventivă preclinică a resveratrolului: o enigmă. Biomarkeri de epidemiol de cancer Prev. 2003; 12 :953–957. [ PubMed ] [ Google Scholar ]28.
Delmas D, Lin HY. Rolul proceselor de dinamică membranară și al moleculelor exogene în absorbția celulară a resveratrolului: consecințe în biodisponibilitate și activități. Mol Nutr Food Res. 2011; 55 :1142–1153. doi: 10.1002/mnfr.201100065. [ PubMed ] [ CrossRef ] [ Google Scholar ]29.
Lancon A, Hanet N, Jannin B, Delmas D, Heydel JM, Lizard G, et al. Resveratrolul în celulele HepG2 de hepatom uman: metabolismul și inductibilitatea enzimelor de detoxifiere. Drug Metab Dispos. 2007; 35 :699–703. doi: 10.1124/dmd.106.013664. [ PubMed ] [ CrossRef ] [ Google Scholar ]30.
Boocock DJ, Faust GE, Patel KR, Schinas AM, Brown VA, Ducharme MP, et al. Studiu farmacocinetic de escaladare a dozei de fază I la voluntari sănătoși cu resveratrol, un potențial agent chimiopreventiv al cancerului. Biomarkeri de epidemiol de cancer Prev. 2007; 16 :1246–1252. doi: 10.1158/1055-9965.EPI-07-0022. [ PubMed ] [ CrossRef ] [ Google Scholar ]31.
Patel KR, Brown VA, Jones DJ, Britton RG, Hemingway D, Miller AS, et al. Farmacologia clinică a resveratrolului și a metaboliților săi la pacienții cu cancer colorectal. Cancer Res. 2010; 70 :7392–7399. doi: 10.1158/0008-5472.CAN-10-2027. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]32.
Feghali CA, Wright TM. Citokinele în inflamația acută și cronică. Front Biosci. 1997; 2 :d12–d26. doi: 10.2741/A171. [ PubMed ] [ CrossRef ] [ Google Scholar ]33.
Kolios G, Brown Z, Robson RL, Robertson DA, Westwick J. Activitatea și expresia sintazei de oxid nitric inductibile într-o linie de celule epiteliale colonice umane, HT-29. Brit J Pharmacol. 1995; 116 :2866–2872. doi: 10.1111/j.1476-5381.1995.tb15938.x. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]35.
Paul S, Rimando AM, Lee HJ, Ji Y, Reddy BS, Suh N. Acțiunea antiinflamatoare a pterostilbenului este mediată prin calea protein kinazei activate de mitogen p38 în celulele cancerului de colon. Cancer Prev Res. 2009; 2 :650–657. doi: 10.1158/1940-6207.CAPR-08-0224. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]36.
Liang YC, Huang YT, Tsai SH, Lin-Shiau SY, Chen CF, Lin JK. Suprimarea ciclooxigenazei inductibile și a oxidului nitric sintetazei inductibile de către apigenină și flavonoide înrudite în macrofagele de șoarece. Carcinogeneza. 1999; 20 :1945–1952. doi: 10.1093/carcin/20.10.1945. [ PubMed ] [ CrossRef ] [ Google Scholar ]37.
Sternlicht MD, Werb Z. Cum metaloproteinazele matricei reglează comportamentul celular. Annu Rev Cell Dev Biol. 2001; 17 :463–516. doi: 10.1146/annurev.cellbio.17.1.463. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]38.
Jiang MC, Liao CF, Lee PH. Aspirina inhibă activitatea metaloproteinazei-2 a matricei, crește producția de E-cadherină și inhibă invazia in vitro a celulelor tumorale. Biochem Biophys Res Commun. 2001; 282 :671–677. doi: 10.1006/bbrc.2001.4637. [ PubMed ] [ CrossRef ] [ Google Scholar ]39.
Aimes RT, Quigley JP. Metaloproteinaza-2 a matricei este o colagenază interstițială. Enzima fără inhibitori catalizează scindarea fibrilelor de colagen și a colagenului nativ solubil de tip I generând fragmentele specifice de 3/4 și 1/4 de lungime. J Biol Chem. 1995; 270 (11):5872–5876. doi: 10.1074/jbc.270.11.5872. [ PubMed ] [ CrossRef ] [ Google Scholar ]40.
Kleiner DE, Jr, Stetler-Stevenson WG. Biochimia structurală și activarea metaloproteazelor matriceale. Curr Opin Cell Biol. 1993; 5 (5):891–897. doi: 10.1016/0955-0674(93)90040-W. [ PubMed ] [ CrossRef ] [ Google Scholar ]41.
Lochter A, Bissell MJ. O odisee de la sân la os: controlul în mai multe etape al metastazelor mamare și al osteolizei prin metaloproteinaze ale matricei. APMIS. 1999; 107 :128–136. doi: 10.1111/j.1699-0463.1999.tb01535.x. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]42.
Bramhall SR. Metaloproteinazele matriceale și inhibitorii lor în cancerul pancreatic. De la știința moleculară la o aplicație clinică. Int J Pancreatol. 1997; 21 (1):1–12. [ PubMed ] [ Google Scholar ]43.
Liu PL, Tsai JR, Charles AL, Hwang JJ, Chou SH, Ping YH și colab. Resveratrolul inhibă metastaza celulelor de adenocarcinom pulmonar uman prin suprimarea căii factorului nuclear-kappaB mediată de hemoxigenaza 1 și, ulterior, prin reglarea în jos a expresiei metaloproteinazelor matriceale. Mol Nutr Food Res. 2010; 54 :S196–S204. doi: 10.1002/mnfr.200900550. [ PubMed ] [ CrossRef ] [ Google Scholar ]44.
Chan JY, Phoo MS, Clement MV, Pervaiz S, Lee SC. Resveratrolul prezintă efecte inverse legate de doză asupra apoptozei celulelor canceroase de colon evocate de 5-fluorouracil: rolurile caspazei-6 și p53. Cancer Biol Ther. 2008; 7 :1305–1312. doi: 10.4161/cbt.7.8.6302. [ PubMed ] [ CrossRef ] [ Google Scholar ]45.
Temraz S, Mukherji D, Shamseddine A. Ținte potențiale pentru prevenirea cancerului colorectal. Int J Mol Sci. 2013; 14 :17279–17303. doi: 10.3390/ijms140917279. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]46.
Balkwill F. Factor de necroză tumorală sau factor de promovare a tumorii? Cytokine Growth Factor Rev. 2002; 13 :135–141. doi: 10.1016/S1359-6101(01)00020-X. [ PubMed ] [ CrossRef ] [ Google Scholar ]47.
Balkwill F, Mantovani A. Inflamație și cancer: înapoi la Virchow? Lancet (Londra, Anglia) 2001; 357 :539–545. doi: 10.1016/S0140-6736(00)04046-0. [ PubMed ] [ CrossRef ] [ Google Scholar ]48.
Albini A, Cesana E, Noonan DM. Celulele stem canceroase și micromediul tumoral: soliști sau cântăreți corali. Curr Pharm Biotechnol. 2011; 12 :171–181. doi: 10.2174/138920111794295756. [ PubMed ] [ CrossRef ] [ Google Scholar ]49.
Boghossian S, Hawash A. Chemoprevention în cancerul colorectal — unde ne aflăm și ce am învățat din experiența de douăzeci de ani. Chirurg. 2012; 10 :43–52. doi: 10.1016/j.surge.2011.07.003. [ PubMed ] [ CrossRef ] [ Google Scholar ]50.
Kimura Y, Sumiyoshi M, Baba K. Activitățile antitumorale ale stilbenelor sintetice și naturale prin acțiune antiangiogenă. Cancer Sci. 2008; 99 :2083–2096. doi: 10.1111/j.1349-7006.2008.00938.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]51.
Cianchi F, Cortesini C, Fantappie O, Messerini L, Schiavone N, Vannacci A, et al. Expresia inductibilă a oxidului nitric sintazei în cancerul colorectal uman: corelație cu angiogeneza tumorii. Sunt J Pathol. 2003; 162 :793–801. doi: 10.1016/S0002-9440(10)63876-X. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]52.
Pandurangan AK, Esa NM. Factori dietetici non-nutritivi în țintirea moleculelor de reglementare în cancerul colorectal: o actualizare. Asian Pac J Cancer Prev. 2013; 14 :5543–5552. doi: 10.7314/APJCP.2013.14.10.5543. [ PubMed ] [ CrossRef ] [ Google Scholar ]53.
Panaro MA, Carofiglio V, Acquafredda A, Cavallo P, Cianciulli A. Efectele antiinflamatorii ale resveratrolului apar prin inhibarea activării NF-kappaB indusă de lipopolizaharide în celulele canceroase de colon umane Caco-2 și SW480. Br J Nutr. 2012; 108 :1623–1632. doi: 10.1017/S0007114511007227. [ PubMed ] [ CrossRef ] [ Google Scholar ]54.
Cui X, Jin Y, Hofseth AB, Pena E, Habiger J, Chumanevich A, et al. Resveratrolul suprimă colita și cancerul de colon asociate cu colita. Cancer Prev Res. 2010; 3 :549–559. doi: 10.1158/1940-6207.CAPR-09-0117. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]55.
Sengottuvelan M, Deeptha K, Nalini N. Resveratrolul ameliorează deteriorarea ADN-ului, dezechilibrul prooxidant și antioxidant în carcinogeneza de colon la șobolan indusă de 1,2-dimetilhidrazină. Chem Biol Interact. 2009; 181 :193–201. doi: 10.1016/j.cbi.2009.06.004. [ PubMed ] [ CrossRef ] [ Google Scholar ]56.
Wang T, Wang G, Zhang Y, Zhang J, Cao W, Chen X. Efectul supraexprimării mediate de lentivirus sau tăcere a MnSOD asupra apoptozei sinoviocitelor asemănătoare fibroblastelor tratate cu resveratrol în artrita reumatoidă. Eur J Pharmacol. 2018; 844 :65–72. doi: 10.1016/j.ejphar.2018.12.001. [ PubMed ] [ CrossRef ] [ Google Scholar ]57.
Olas B, Nowak P, Kolodziejczyk J, Ponczek M, Wachowicz B. Efectele protectoare ale resveratrolului împotriva modificărilor oxidative/nitrative ale proteinelor plasmatice și lipidelor expuse la peroxinitrit. J Nutr Biochem. 2006; 17 :96–102. doi: 10.1016/j.jnutbio.2005.05.010. [ PubMed ] [ CrossRef ] [ Google Scholar ]58.
Xia N, Daiber A, Habermeier A, Closs EI, Thum T, Spanier G, et al. Resveratrolul inversează decuplarea endotelială a oxidului nitric sintazei la șoarecii knockout de apolipoproteina E. J Pharmacol Exp Ther. 2010; 335 :149–154. doi: 10.1124/jpet.110.168724. [ PubMed ] [ CrossRef ] [ Google Scholar ]59.
Chiou YS, Tsai ML, Nagabhushanam K, Wang YJ, Wu CH, Ho CT și colab. Pterostilbenul este mai puternic decât resveratrolul în prevenirea tumorigenezei colonului indusă de azoximetan (AOM) prin activarea căii de semnalizare antioxidantă mediată de factorul 2 (Nrf2) legat de NF-E2. J Agric Food Chim. 2011; 59 :2725–2733. doi: 10.1021/jf2000103. [ PubMed ] [ CrossRef ] [ Google Scholar ]60.
Tamaki N, Cristina Orihuela-Campos R, Inagaki Y, Fukui M, Nagata T, Ito HO. Resveratrolul îmbunătățește stresul oxidativ și previne progresia parodontitei prin activarea căilor de apărare Sirt1/AMPK și Nrf2/antioxidant într-un model de parodontită de șobolan. Free Radic Biol Med. 2014; 75 :222–229. doi: 10.1016/j.freeradbiomed.2014.07.034. [ PubMed ] [ CrossRef ] [ Google Scholar ]61.
Van Meter TE, Broaddus WC, Cash D, Fillmore H. Cotratamentul cu un nou inhibitor de analog fosfoinozitid și carmustină îmbunătățește eficacitatea chimioterapeutică prin atenuarea activității AKT în gliom. Cancer. 2006; 107 :2446–2454. doi: 10.1002/cncr.22248. [ PubMed ] [ CrossRef ] [ Google Scholar ]62.
Al Zaid Siddiquee K, Turkson J. STAT3 ca țintă pentru inducerea apoptozei în tumorile solide și hematologice. Cell Res. 2008; 18 (2):254–267. doi: 10.1038/cr.2008.18. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]63.
Santandreu FM, Valle A, Oliver J, Roca P. Resveratrolul potențează stresul oxidativ citotoxic indus de chimioterapie în celulele cancerului de colon uman. Cell Physiol Biochim. 2011; 28 :219–228. doi: 10.1159/000331733. [ PubMed ] [ CrossRef ] [ Google Scholar ]64.
Wallerath T, Deckert G, Ternes T, Anderson H, Li H, Witte K și colab. Resveratrolul, o fitoalexină polifenolică prezentă în vinul roșu, îmbunătățește expresia și activitatea oxidului nitric sintazei endoteliale. Circulaţie. 2002; 106 :1652–1658. doi: 10.1161/01.CIR.0000029925.18593.5C. [ PubMed ] [ CrossRef ] [ Google Scholar ]65.
Russell J, Okayama N, Alexander JS, Granger DN, Hsia CJ. Pretratamentul cu polinitroxil albumină (PNA) inhibă aderența leucocitelor-celule endoteliale indusă de ischemie-reperfuzie. Free Radic Biol Med. 1998; 25 (2):153–159. doi: 10.1016/S0891-5849(98)00022-7. [ PubMed ] [ CrossRef ] [ Google Scholar ]66.
Thom SR, Bhopale VM, Milovanova TN, Yang M, Bogush M, Buerk DG. Legătura oxidului azot sintaza-2 la kinaza de adeziune focală în neutrofile influențează activitatea enzimatică și funcția integrinei beta2. J Biol Chem. 2013; 288 :4810–4818. doi: 10.1074/jbc.M112.426353. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]67.
Guan JL. Rolul kinazei de adeziune focală în semnalizarea integrinei. Int J Biochem Cell Biol. 1997; 29 :1085–1096. doi: 10.1016/S1357-2725(97)00051-4. [ PubMed ] [ CrossRef ] [ Google Scholar ]69.
Cary LA, Guan JL. Kinaza de adeziune focală în semnalizarea mediată de integrină. Front Biosci. 1999; 4 :D102–D113. doi: 10.2741/Cary. [ PubMed ] [ CrossRef ] [ Google Scholar ]70.
Kong D, Chen F, Sima NI. Inhibarea kinazei de adeziune focală induce apoptoza în celulele canceroase ale vezicii urinare prin Src și calea fosfatidilinozitol 3-kinaze/Akt. Exp Ther Med. 2015; 10 :1725–1731. doi: 10.3892/etm.2015.2745. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]71.
Buhrmann C, Shayan P, Goel A, Shakibaei M. Resveratrolul reglează invazia celulelor canceroase colorectale prin modularea moleculelor de adeziune focală. Nutrienți. 2017; 9 (10): E1073. doi: 10.3390/nu9101073. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]72.
Bragdon B, Moseychuk O, Saldanha S, King D, Julian J, Nohe A. Proteine morfogenetice osoase: o revizuire critică. Semnal celular. 2011; 23 :609–620. doi: 10.1016/j.cellsig.2010.10.003. [ PubMed ] [ CrossRef ] [ Google Scholar ]73.
Zhang L, Ye Y, Long X, Xiao P, Ren X, Yu J. Semnalizarea BMP și efectele sale paradoxale în tumorigeneză și diseminare. Oncotarget. 2016; 7 :78206–78218. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]74.
Cassar L, Nicholls C, Pinto AR, Chen R, Wang L, Li H, et al. Inhibarea telomerazei mediată de receptorul TGF-beta, scurtarea telomerilor și senescența celulelor canceroase de sân. Celula proteică. 2017; 8 :39–54. doi: 10.1007/s13238-016-0322-1. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]75.
Leinhauser I, Richter A, Lee M, Hofig I, Anastasov N, Fend F, et al. Caracteristicile oncogene ale proteinei morfogenice osoase 7 (BMP7) în feocromocitom. Oncotarget. 2015; 6 :39111–39126. doi: 10.18632/oncotarget.4912. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]76.
Fresno Vara JA, Casado E, de Castro J, Cejas P, Belda-Iniesta C, Gonzalez-Baron M. PI3K/Akt signaling pathway and cancer. Cancer Treat Rev. 2004; 30 (2):193–204. doi: 10.1016/j.ctrv.2003.07.007. [ PubMed ] [ CrossRef ] [ Google Scholar ]77.
Zeng YH, Zhou LY, Chen QZ, Li Y, Shao Y, Ren WY și colab. Resveratrolul inactivează semnalizarea PI3K/Akt prin reglarea pozitivă a BMP7 în celulele canceroase de colon umane. Oncol Rep. 2017; 38 :456–464. doi: 10.3892/or.2017.5662. [ PubMed ] [ CrossRef ] [ Google Scholar ]78.
Chen HJ, Hsu LS, Shia YT, Lin MW, Lin CM. Complexul beta-catenină/TCF ca o țintă nouă a resveratrolului în calea de semnalizare Wnt/beta-catenină. Biochem Pharmacol. 2012; 84 :1143–1153. doi: 10.1016/j.bcp.2012.08.011. [ PubMed ] [ CrossRef ] [ Google Scholar ]79.
Wolter F, Akoglu B, Clausnitzer A, Stein J. Reglarea în jos a complexului ciclin D1/Cdk4 are loc în timpul opririi ciclului celular indus de resveratrol în liniile celulare de cancer de colon. J Nutr. 2001; 131 :2197–2203. doi: 10.1093/jn/131.8.2197. [ PubMed ] [ CrossRef ] [ Google Scholar ]80.
Suzuki R, Kuroda H, Komatsu H, Hosokawa Y, Kagami Y, Ogura M, et al. Utilizarea selectivă a ciclinelor de tip D în afecțiunile maligne limfoide. leucemie. 1999; 13 :1335–1342. doi: 10.1038/sj.leu.2401485. [ PubMed ] [ CrossRef ] [ Google Scholar ]81.
Carlson B, Lahusen T, Singh S, Loaiza-Perez A, Worland PJ, Pestell R, et al. Reglarea în jos a ciclinei D1 prin reprimarea transcripțională în celulele de carcinom mamar uman MCF-7 induse de flavopiridol. Cancer Res. 1999; 59 :4634–4641. [ PubMed ] [ Google Scholar ]82.
Hsieh TC, Wu JM. Efecte diferențiate asupra creșterii, opririi ciclului celular și inducerea apoptozei de către resveratrol în liniile celulare de cancer de prostată uman. Exp Cell Res. 1999; 249 :109–115. doi: 10.1006/excr.1999.4471. [ PubMed ] [ CrossRef ] [ Google Scholar ]83.
Fontecave M, Lepoivre M, Elleingand E, Gerez C, Guittet O. Resveratrol, un inhibitor remarcabil al ribonucleotide reductazei. FEBS Lett. 1998; 421 :277–279. doi: 10.1016/S0014-5793(97)01572-X. [ PubMed ] [ CrossRef ] [ Google Scholar ]84.
Sengottuvelan M, Deeptha K, Nalini N. Influența resveratrolului dietetic asupra markerilor moleculari timpurii și tardivi ai carcinogenezei de colon induse de 1,2-dimetilhidrazină. Nutriție. 2009; 25 :1169–1176. doi: 10.1016/j.nut.2009.03.009. [ PubMed ] [ CrossRef ] [ Google Scholar ]85.
Gescher A, Steward WP, Brown K. Resveratrolul în managementul cancerului uman: cât de puternice sunt dovezile clinice? Ann NY Acad Sci. 2013; 1290 :12–20. doi: 10.1111/nyas.12205. [ PubMed ] [ CrossRef ] [ Google Scholar ]86.
Patel KR, Scott E, Brown VA, Gescher AJ, Steward WP, Brown K. Studii clinice ale resveratrolului. Ann NY Acad Sci. 2011; 1215 :161–169. doi: 10.1111/j.1749-6632.2010.05853.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]87.
Chow HH, Garland LL, Hsu CH, Vining DR, Chew WM, Miller JA și colab. Resveratrolul modulează enzimele care metabolizează medicamentele și cancerele într-un studiu pe voluntari sănătoși. Cancer Prev Res. 2010; 3 :1168–1175. doi: 10.1158/1940-6207.CAPR-09-0155. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]88.
Howells LM, Berry DP, Elliott PJ, Jacobson EW, Hoffmann E, Hegarty B și colab. Studiu pilot de fază I, randomizat, dublu-orb, al resveratrolului micronizat (SRT501) la pacienții cu metastaze hepatice – siguranță, farmacocinetică și farmacodinamică. Cancer Prev Res. 2011; 4 :1419–1425. doi: 10.1158/1940-6207.CAPR-11-0148. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]89.
Liang L, Liu X, Wang Q, Cheng S, Zhang S, Zhang M. Farmacocinetica, distribuția tisulară și studiul excreției resveratrolului și al promedicamentului său 3,5,4′-tri-O-acetilresveratrol la șobolani. Fitomedicina. 2013; 20 :558–563. doi: 10.1016/j.phymed.2012.12.012. [ PubMed ] [ CrossRef ] [ Google Scholar ]90.
Johnson JJ, Nihal M, Siddiqui IA, Scarlett CO, Bailey HH, Mukhtar H, et al. Creșterea biodisponibilității resveratrolului prin combinarea acestuia cu piperina. Mol Nutr Food Res. 2011; 55 :1169–1176. doi: 10.1002/mnfr.201100117. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]91.
la Porte C, Voduc N, Zhang G, Seguin I, Tardiff D, Singhal N, et al. Farmacocinetica la starea de echilibru și tolerabilitatea trans-resveratrolului 2000 mg de două ori pe zi cu alimente, quercetină și alcool (etanol) la subiecții umani sănătoși. Clin Pharmacokinet. 2010; 49 :449–454. doi: 10.2165/11531820-000000000-00000. [ PubMed ] [ CrossRef ] [ Google Scholar ]92.
Smoliga JM, Blanchard O. Îmbunătățirea eliberării de resveratrol la om: dacă biodisponibilitatea scăzută este problema, care este soluția? Molecule. 2014; 19 :17154–17172. doi: 10,3390/molecules191117154. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]93.
Wang S, Su R, Nie S, Sun M, Zhang J, Wu D și colab. Aplicarea nanotehnologiei în îmbunătățirea biodisponibilității și bioactivității substanțelor fitochimice derivate din dietă. J Nutr Bioch. 2014; 25 :363–376. doi: 10.1016/j.jnutbio.2013.10.002. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]94.
Paller CJ, Rudek MA, Zhou XC, Wagner WD, Hudson TS, Anders N și colab. Un studiu de fază I al extractului de piele de struguri de muscadină la bărbații cu cancer de prostată recurent biochimic: siguranță, tolerabilitate și determinarea dozei. Prostata. 2015; 75 :1518–1525. doi: 10.1002/pros.23024. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]95.
Kjaer TN, Ornstrup MJ, Poulsen MM, Jorgensen JO, Hougaard DM, Cohen AS, et al. Resveratrolul reduce nivelul precursorilor androgeni circulanți, dar nu are efect asupra testosteronului, dihidrotestosteronului, nivelurilor de PSA sau volumului prostatei. Un studiu randomizat de 4 luni la bărbați de vârstă mijlocie. Prostata. 2015; 75 :1255–1263. doi: 10.1002/pros.23006. [ PubMed ] [ CrossRef ] [ Google Scholar ]96.
Jazirehi AR, Bonavida B. Resveratrolul modifică expresia proteinelor reglatoare apoptotice și sensibilizează limfomul non-Hodgkin și liniile celulare de mielom multiplu la apoptoza indusă de paclitaxel. Mol Cancer Ther. 2004; 3 (1):71–84. doi: 10.4161/cbt.3.1.683. [ PubMed ] [ CrossRef ] [ Google Scholar ]97.
Popat R, Plesner T, Davies F, Cook G, Cook M, Elliott P, et al. Un studiu de fază 2 al SRT501 (resveratrol) cu bortezomib pentru pacienții cu mielom multiplu recidivat și/sau refractar. Br J Haematol. 2013; 160 :714–717. doi: 10.1111/bjh.12154. [ PubMed ] [ CrossRef ] [ Google Scholar ]98.
Sale S, Tunstall RG, Ruparelia KC, Potter GA, Steward WP, Gescher AJ. Comparația efectelor agentului chimiopreventiv resveratrol și analogului său sintetic trans 3,4,5,4′-tetrametoxistilben (DMU-212) asupra dezvoltării adenomului la șoarecele Apc(Min +) și ciclooxigenaza-2 în cancerul de colon derivat de la om celule. Int J Cancer. 2005; 115 :194–201. doi: 10.1002/ijc.20884. [ PubMed ] [ CrossRef ] [ Google Scholar ]99.
Schneider Y, Vincent F, Duranton B, Badolo L, Gosse F, Bergmann C, et al. Efectul anti-proliferativ al resveratrolului, o componentă naturală a strugurilor și a vinului, asupra celulelor canceroase de colon umane. Cancer Lett. 2000; 158 :85–91. doi: 10.1016/S0304-3835(00)00511-5. [ PubMed ] [ CrossRef ] [ Google Scholar ]100.
Nguyen AV, Martinez M, Stamos MJ, Moyer MP, Planutis K, Hope C și colab. Rezultatele unui studiu clinic pilot de fază I care examinează efectul resveratrolului derivat din plante și al pulberii de struguri asupra expresiei genei țintă a căii Wnt în mucoasa colonică și cancerul de colon. Cancer Manag Res. 2009; 1 :25–37. doi: 10.2147/CMAR.S4544. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]101.
Zhu W, Qin W, Zhang K, Rottinghaus GE, Chen YC, Kliethermes B, et al. Trans-resveratrolul modifică hipermetilarea promotorului mamar la femeile cu risc crescut de cancer de sân. Nutr Cancer. 2012; 64 :393–400. doi: 10.1080/01635581.2012.654926. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
Trans- resveratrolul, prezent în concentrație ridicată în pielea strugurilor roșii și a vinului roșu, are un efect antiproliferativ dependent de doză in vitro, previne formarea tumorilor mamare și a fost susținut ca agent chemopreventiv. Pe baza studiilor in vitro care demonstrează că trans- resveratrolul reglează în jos expresia 1) ADN metiltransferazelor și 2) a cancerului care promovează prostaglandina (PG) E 2 , am stabilit dacă trans- resveratrolul a avut un efect legat de doză asupra metilării ADN și a expresiei prostaglandinelor în oameni. Treizeci și nouă de femei adulte cu risc crescut de cancer mamar au fost randomizate în mod dublu-orb până la placebo, 5 sau 50 mg trans resveratrolul de două ori pe zi timp de 12 săptămâni. Evaluarea metilării a 4 gene legate de cancer ( p16, RASSF-1α, APC, CCND2 ) a fost efectuată pe probe de ductoscopie mamară. Specia predominantă de resveratrol în ser a fost metabolitul glucuronid. Total tranznivelurile serice ale metabolitului de resveratrol și glucuronid au crescut după consumul ambelor doze de trans- resveratrol ( P <0,001 pentru ambele). Metilarea RASSF-1 α a scăzut odată cu creșterea nivelului de trans- resveratrol seric ( P = .047). Modificarea metilării RASSF-1 α a fost direct legată de modificarea PGE 2 ( P= .045). Această lucrare oferă informații noi despre efectele trans- resveratrolului asupra sânului femeilor cu risc crescut de cancer mamar, inclusiv o scădere a metilării genei supresoare tumorale RASSF-1 α. Datorită dimensiunii limitate a eșantionului, constatările noastre ar trebui validate într-un studiu mai amplu.
INTRODUCERE
Trans- resveratrolul este un polifenol prezent în alimente care a fost recunoscut ca agent anticancer ( 1 ). De cis izomerul este în general prezent la niveluri mai scăzute în produsele alimentare și considerate a avea proprietăți anticancer mai puțin potent ( 2 ). Trans- resveratrolul se găsește în concentrație mare în pielea strugurilor roșii și în vinul roșu, cu cantități mai mici în dud și arahide. Niveluri ridicate se găsesc și în rizomul P. cuspidatum , iar majoritatea suplimentelor alimentare trans- resveratrol conțin P. cuspidatum . Trans- resveratrolul inhibă proliferarea epitelială a sânului uman într-un mod dependent de doză și de timp ( 3), sugerând că agentul are efecte chemopreventive. Agentul a inhibat formarea leziunilor mamare preneoplazice mamare de șoarece ( 4 ). De asemenea, a redus incidența tumorilor mamare cu 45% și multiplicitatea cu 55% atunci când a fost administrată ca parte a dietei la o doză de 10 părți pe milion la șobolanii femele Sprague Dawley tratați cu DMBA comparativ cu șobolanii tratați cu DMBA care nu au primit resveratrol. ( 5 ).
Un studiu caz-control a demonstrat un risc relativ de cancer de sân de 0,39 la femeile cu niveluri ridicate și 0,50 la femeile cu niveluri intermediare ale aportului total de resveratrol comparativ cu cele cu un nivel scăzut de ingestie ( 6 ). În ciuda dovezilor preclinice și epidemiologice convingătoare ale efectelor chemopreventive ale agentului, studiile care abordează mecanismul său de acțiune la om sunt limitate și se concentrează asupra efectului resveratrolului asupra proliferării.
Insulele CpG ale multor gene care sunt în mare parte nemetilate în țesutul normal sunt hipermetilate în grade diferite în cancerul de sân ( 7 ). Hipermetilarea se poate dezvolta devreme în procesul de carcinogeneză. Am demonstrat în epiteliul ductal al sânului la femei cu risc normal de cancer mamar că modificările de metilare apar la genele supresoare tumorale după ingestia polifenolilor genisteină și daidzeină prezente în soia ( 8 ). Se știe că metilarea dinucleotidelor CpG este mediată de cel puțin 3DNAmetil transferaze (DNMT): DNMT1, DNMT3a și DNMT3b ( 9 , 10 ). Scăderea expresiei DNMT1 și 3b într-o linie celulară de cancer de colon a dus la reexprimarea genelor supresoare tumorale ( 11 ). Am raportat anterior cătrans- resveratrolul a scăzut expresia DNTMT1 și 3b în celulele cancerului de sân într-o manieră dependentă de doză ( 12 ). Este plauzibil faptul că expresia redusă a ADN metiltransferazei 1 și 3b indusă de trans- resveratrol poate duce la demetilare și reexpresie a genelor supresoare tumorale.
Trans- resveratrolul este absorbit eficient după administrarea orală ( 13 ) și metabolizat rapid prin glucuronidare și sulfatare în ficat ( 14 ). Acest lucru este similar cu alte substanțe botanice, cum ar fi soia, unde polifenolii genistein și daidzein ( 15 ) sunt metabolizați rapid. În sânge, trans- resveratrolul se leagă atât de albumină, cât și de lipoproteine cu densitate mică (LDL) ( 16 ), ceea ce ajută la livrarea sa către suprafața celulei epiteliale pentru absorbția membranei celulare, permițând efectul biologic intracelular ( 16 ). Glucoronida poate fi scindată enzimatic ( 17 ) sau trans- resveratrol sulfat hidrolizat ( 18) la forma sa liberă, astfel încât, după administrarea resveratrolului, nivelurile intracelulare la rinichi, de exemplu, au fost predominant sub forma liberă, dar în urină predomină metaboliții ( 19 ). Se pare că cel puțin unii derivați de resveratrol au, de asemenea, un efect antiproliferativ ( 20 ).
Studiile preclinice sugerează că proprietățile chemopreventive ale trans- resveratrolului se datorează apoptoticului agentului, opririi ciclului celular, inhibării kinazei și proprietăților antiangiogene ( 21 ). Pe baza observației noastre că trans- resveratrolul reglează în jos DNMT-urile în cancerul de sân cu receptor estrogen pozitiv ( 12 ), am efectuat un studiu clinic dublu-orb, randomizat, controlat cu placebo la femei cu risc crescut de cancer de sân, pentru a determina efectele dependente de doză ale trans. -resveratrol pe 4 gene, inclusiv 3 ( CCND-2 , p16 și RASSF-1 α) pe care le-am demonstrat anterior ( 22) au fost semnificativ mai răspândite în tumoare decât în specimenele de țesut normal, iar gena APC , a cărei metilare aberantă este prezentă în special în forme mai agresive de cancer, cum ar fi cancerul de sân inflamator ( 23 ). De asemenea, am evaluat efectele resveratrolului asupra expresiei E 2 a prostaglandinei (PG) sistemice și specifice sânilor . Mai multe linii de dovezi susțin proprietățile de promovare a tumorii ale PGE 2 . Expresia PGE 2 este direct legată de proliferarea celulară ( 24 ), tumorile maligne ale sânului produc mai mult PGE 2 decât tumorile benigne ale sânului sau țesutul mamar normal ( 25 ) și femeile cu cancer mamar care au avut PGE 2nivelurile peste 15 ng / g au avut o rată de supraviețuire semnificativ mai slabă decât cele cu niveluri ≤15 ng / g ( 26 ). Am corelat modificările PGE 2 și ale metilării genelor cu doza de resveratrol și cu nivelurile circulante ale agentului și ale metaboliților săi.
Dozele de resveratrol au fost alese pe baza a ceea ce este disponibil în mod obișnuit în magazinele naturiste. Doza mai mică de trans- resveratrol este aproximativ de două ori cantitatea pe care o primește o persoană când consumă 2 pahare de vin roșu ( 14 ). Un studiu preclinic care a administrat o doză de resveratrol echivalentă cu aproximativ 14 mg / zi resveratrol la un om de 70 kg a suprimat în mod semnificativ formarea tumorilor mamare ( 5 ). Doza mai mare este mai mare decât se găsește în general în alimente, dar am dorit să evaluăm o doză mai mare de agent promovat pentru consum ca nutraceutic în magazinele de produse naturiste ( 27 ).Mergi la:
MATERIALE ȘI METODE
Subiecte
Pentru a fi eligibile, femeile trebuiau să aibă un risc crescut de cancer mamar, definit ca având o rudă de gradul I cu cancer mamar, un risc Gail de> 1,66% din dezvoltarea cancerului mamar în următorii 5 ani și / sau un istoric personal de o biopsie mamară care demonstrează hiperplazie atipică, in situ sau cancer de sân invaziv (tratat anterior și în prezent lipsit de boală). Fiecare participant a semnat consimțământul informat aprobat de un comitet de evaluare internă înainte de înscriere. Participantul a fost apoi randomizat în mod dublu-orb prin metoda plicurilor sigilate și a luat o capsulă de două ori pe zi timp de 12 săptămâni conținând 1 din 3 preparate: placebo, P. cuspidatum cu 5 mg trans- resveratrol sau P. cuspidatum cu 50 mg de trans-resveratrol. Regimul de două ori pe zi (mai degrabă decât dozarea zilnică) a fost ales din cauza metabolismului rapid al compusului părinte ( 27 ).
Au fost excluse femeile însărcinate, care alăptau sau care alăptaseră în decurs de 20 de luni de la înscrierea la studiu. Femeile nu ar fi putut lua estrogeni și / sau progestini exogeni în luna anterioară înscrierii sau în timpul studiului. Ei trebuie să fi fost supuși unui examen negativ de sân și, pentru femeile cu vârsta de 40 de ani sau mai mult, o mamografie benignă în termen de 6 luni de la înscriere. Subiecții au fost rugați să nu consume vin roșu sau să mănânce duduri și să limiteze consumul de arahide / produse de arahide la 1 oz. zilnic în timpul studiului. Consumul obișnuit de macronutrienți a fost evaluat și menținut la niveluri constante pe tot parcursul studiului. Consumul de medicamente pe bază de plante care conținea sau ar putea conține fitoestrogeni activi hormonal a fost interzis. Suplimentele alimentare funcționale concentrate (de exemplu, capsule sau pulberi) nu au fost permise.De asemenea, au fost excluse femeile care au menținut o dietă vegană sau macrobiotică. Subiecții care au luat tamoxifen, raloxifen sau un inhibitor de aromatază au fost excluși.
Subiecții au fost întrebați la momentul înscrierii dacă luau medicamente antiinflamatoare nesteroidiene (AINS) și aceste informații au fost înregistrate. Subiecților care iau AINS li s-a cerut să își mențină practica normală de utilizare. Subiecților li s-a dat un calendar pe care să înregistreze fiecare capsulă luată. Subiecții considerați că ingerează zilnic peste 500 µg trans- resveratrol din toate sursele au fost considerați neevaluabili. Preparatele trans- resveratrol au fost furnizate ca un singur lot de către InterHealth Nutraceuticals (Benicia, CA). A fost utilizat un preparat placebo constând din celuloză microcristalină. La întoarcere vizite 4 și 12 săptămâni după începerea trans-resveratrol, participanții au fost întrebați dacă au existat efecte secundare legate de medicamente. Conformitatea a fost evaluată printr-un calendar de capsule și prin colectarea de capsule neutilizate.
Colectarea specimenelor
Lichidul aspirat pentru mamelon (NAF) și probele de ser au fost colectate la momentul inițial, 4 și 12 săptămâni după pornirea agentului. Probele de ductoscopie mamară (MD) au fost colectate la momentul inițial și la 12 săptămâni, așa cum s-a descris anterior ( 28 , 29 ). NAF și serul au fost utilizate pentru a analiza PGE 2 . Serul a fost, de asemenea, utilizat pentru a măsura totalul trans – și totalul cis-metaboliti ai resveratrolului si resveratrolului (glucuronid si sulfat). Probele de MD au fost utilizate pentru a evalua modificările metilării. Pentru colectarea MD, a fost utilizată o rețea pentru mamelon pentru a indica locația canalului canulat la momentul inițial. S-au făcut încercări de a canula același canal înainte și după tratament. Toate analizele specimenelor au fost efectuate în mod orbit.
qMS-PCR
Un ADN purtător de spermă de somon a fost adăugat la ADN extras din fiecare probă MD, care a fost apoi tratată cu bisulfit de sodiu. O strategie PCR în 2 pași a fost utilizată pentru p16, RASSF-1α, APC și CCND2 . Prima rundă PCR de ADN MD a fost efectuată pentru cele 4 gene folosind un kit AmpliTaq Gold PCR (Applied Biosystems, Foster City, CA), în care atât copiile metilate cât și nemetilate ale unei gene specifice din probă au fost amplificate cu un primer universal a stabilit. Pentru a doua rundă de SYBR pe bază de verde qMSP, produsele PCR diluate au fost amplificate cu primerii specifici ai celor 4 gene atât pentru ADN metilat cât și pentru ADN nemetilat. Seturile de primer pentru qMSP au fost raportate anterior ( 30 , 31 ). S-a calculat procentul de ADN metilat din fiecare probă ( 32%)). ADN care a fost 100% metilat sau 100% nemetilat a fost utilizat pentru a genera o curbă standard pentru a cuantifica procentul de ADN metilat din fiecare probă.
Validare qMS-PCR
Pentru a verifica specificitatea produselor qMS-PCR din a doua rundă, ampliconii selectați pentru p16, RASSF-1α, APC și CCND2 au fost subclonați utilizând sistemul de clonare TOPO-TA (Invitrogen, Carlsbad, CA). ADN-ul plasmidic din 5-6 clone pozitive a fost izolat și secvențiat folosind un analizor ADN 3730 ADN (Applied Biosystems, Foster City, CA).
Cuantificarea resveratrolului
Capsule
Identitatea botanică a materialului furnizat de InterHealth Nutraceuticals a fost confirmată prin cromatografie lichidă de înaltă performanță (HPLC) la momentul inițial și asigurarea că degradarea nu a avut loc confirmată prin analiza capsulelor aleatorii, anual, ulterior. Două capsule de fiecare tip (placebo, resveratrol cu doze mici, resveratrol cu doze mari) au fost evaluate anual. Standardul pentru calcularea curbei standard a fost obținut de la Sigma. Deoarece la speciile de plante pot fi prezenți atât trans- resveratrol, cât și cis- resveratrol, ambii izomeri au fost măsurați. Forma cis a agliconei a fost obținută prin expunerea UV la trans– izomer. Probele au fost analizate pentru resveratrol total după hidroliza enzimatică a resveratrolului beta-D-glucopiranozide de glucozidază (Sigma) urmată de analiza HPLC cu UV (306 nm) și detectarea fluorescenței (ex 324 nm, em 370 nm).
Ser
Am încercat să măsurăm atât trans- și cis- resveratrolul liber, cât și cel conjugat , precum și metaboliții resuronatrolului glucuronid și sulfat așa cum este descris ( 33 , 34 ). Izomerii de resveratrol în 0,10 ml ser au fost incubați la 37 ° C peste noapte fără enzimă, β-D-glucuronidază / sulfatază și sulfatază (Sigma Aldrich, St. Louis, MO), acidulați la pH 3,0 cu acid acetic, vortexați și centrifugați la 13.000 rpm timp de 5 min. Probele au fost apoi adăugate la coloanele C18 Sep-pak-vac precondiționate (Waters). Coloanele au fost spălate cu apă și trans – și cis-resveratrol eluat cu metanol. Eluantul de metanol a fost dus la sec, reconstituit și analizat folosind un sistem de detectare HPLC ESA CoulArray Model 5600 cu 4 canale împreună cu o pompă HPLC isocratică ESA conectată la un autosamplator Spectrasystem AS3500 Thermo Separation Products utilizând un Luna (250 mm × 4,6 mm) coloană (Phenomenex) cu o fază mobilă de metanol / 50 mM acetat de sodiu (pH 4,8) (47:53, v / v). Setările CoulArray au fost de 200, 300, 450 și 600 mV. O curbă standard a fost construită folosind standardele cis și trans- resveratrol (Sigma). Deoarece metaboliții individuali de resveratrol sulfat și glucuronid nu erau disponibili cu ușurință, cis – și trans- resveratrol gratuit (fără enzimă), cis total – șis-au determinat concentrațiile trans- resveratrol (β-D-glucuronidază / sulfatază), și concentrațiile totale de sulfat de resveratrol (sulfatază). Nu s-a identificat cis – nici trans- resveratrol liber. Prin urmare, pentru a determina concentrația totală de glucuronid de resveratrol, am scăzut concentrația de sulfat de resveratrol total din concentrația combinată de trans – plus cis- resveratrol total . Eficiența extracției a fost> 90% -95%. Analiza duplicat a fost efectuată pentru multe dintre probe, cu rezultate consistente.
PGE 2
Fiecare specimen NAF și ser, standard și control au fost analizate în duplicat. PGE 2 a fost măsurat prin imunotest conform descrierii anterioare ( 35 ) conform instrucțiunilor producătorului (Assay Design, Inc., Ann Arbor, MI). Pe scurt, probele au fost diluate în tampon de testare de 100 µL furnizat de producător, pipetate în godeuri adecvate, incubate timp de 2 ore la temperatura camerei pe un agitator de plăci, spălate, soluție de substrat adăugată, urmată de incubare de 1 oră și absorbanță măsurată la 405 nm. O curbă standard a fost pregătită folosind diluții seriale ale PGE 2 . O curbă logistică cu 4 parametri a fost creată din standardele de concentrație cunoscută de PGE 2 și PGE 2concentrațiile de probe necunoscute se potrivesc ecuației standard de regresie a curbei, corectate pentru volumul alicotei și exprimate ca nanograme de PGE 2 / ml din proba originală. Calitatea potrivirii curbei standard, R 2 , a fost de 0,9993. A fost furnizat și utilizat un standard QA. CV-ul a fost <10% în intervalul standard.
Analize statistice
Au fost efectuate atât analize în cadrul grupului, cât și între grupuri. Nivelurile serice de resveratrol au fost transformate în logaritm înainte de analiză. A fost folosit un test t asociat pentru a detecta diferențele dintre punctele de timp pentru nivelurile de resveratrol și PGE 2 . Un test între grupul t a fost utilizat pentru a analiza diferențele dintre grupurile de tratament în variabile cantitative, cum ar fi fracțiile de p16 metilat , RASSF-1α, APC și CCND2 . Regresia liniară a fost efectuată pentru a investiga relațiile dintre variabilele cantitative, cum ar fi modificarea nivelurilor de resveratrol și modificările PGE 2 și a fracției de p16 metilat , RASSF-1α, APC și CCND2. Toate analizele au fost efectuate utilizând software-ul SAS JMP.Mergi la:
REZULTATE
Subiecte
Trei sute șase femei au fost examinate și 39 de subiecți înscriși. Motivele pentru care femeile nu au fost eligibile au inclus faptul că nu îndeplinesc criteriile de risc ridicat și / sau au primit un medicament sau un supliment alimentar nepermis în criteriile de excludere. Două femei și-au luat capsulele conform prescripției pentru cel puțin 4 săptămâni și 31 pentru întreaga perioadă de 12 săptămâni a studiului. Dintre cei 8 care nu au finalizat studiul, 1 s-a retras înainte de colectarea eșantionului inițial, 6 au abandonat după colectarea eșantionului din motive personale și o persoană s-a retras înainte de colectarea eșantionului de 12 săptămâni din cauza efectelor secundare. Unul dintre cei 31 de participanți care au terminat studiul a fost exclus din analiza statistică, deoarece timpul de la administrarea ultimei doze de medicamente până la recoltarea eșantionului a depășit 12 ore și trans -12 săptămâni-concentrația de resveratrol în serul său a fost <5% din concentrația de 4 săptămâni, în timp ce toate celelalte comparații de 4 și 12 săptămâni ale nivelurilor de trans- resveratrol au fost similare. O treime dintre participanții la grupurile de resveratrol cu doze mici și placebo și 22% dintre participanții la grupul cu doze mari de resveratrol, au indicat că iau medicamente antiinflamatoare nesteroidiene în momentul înscrierii în studiu. Utilizarea AINS nu a avut un efect semnificativ. la expresia PGE 2 și nici nu am găsit o interacțiune semnificativă între utilizarea AINS și nivelurile de PGE 2 sau de resveratrol.
Am putut analiza biomarkeri în toate momentele în NAF în 22/30 (73%) și în ser la 28/30 (93%) femei. Vârsta medie (61 de ani vs. 59,5 ani vs. 54 de ani) în grupurile placebo comparativ cu doze mici vs. O mediană de 98,2% din capsulele recomandate au fost consumate în timpul studiului. În general, efectele secundare au fost puține și ușoare. Singurul individ care s-a retras din cauza efectelor secundare a avut un istoric de sindrom de colon iritabil și a prezentat diaree CTC grad 1 9 săptămâni în tratament.
Nivelurile de resveratrol în capsule și în ser
La începutul studiului, se presupune că capsulele conțin placebo, 5 și 50 mg conțineau în medie 0 mg, 5,2 mg și respectiv 52,5 mg trans- resveratrol. CV-ul pentru măsurători a fost <10%. Conținutul trans- resveratrol al capsulelor a fost evaluat la intervale anuale ulterior, cu un CV în timp <10%. Timpurile medii și mediane de la ultima doză de resveratrol până la colectarea serului au fost de 4,2 și respectiv 3,4 ore, cu o medie și o medie de 3,8 și 3,5 ore la punctul de timp de 4 săptămâni și 4,7 și 3,3 ore la punctul de timp de 12 săptămâni. Timpurile medii și mediane au fost similare în rândul celor 3 grupuri, la fel ca și intervalul (până la 7,7, 8,6 și 10,9 ore) în grupurile cu resveratrol cu doze mici și, respectiv, cu placebo.
Nivelurile totale (liber plus metaboliți) trans- resveratrol au fost detectabile la 6/30 (20%) probe de ser la momentul inițial. Nivelurile au crescut semnificativ la 4 și 12 săptămâni (Fig. 1A) după ce participanții au luat fie resveratrol cu doze mici, fie mari ( P <.001 pentru toți), dar nu după tratamentul cu placebo. Tratamentul cu resveratrol în doze mari a dus la niveluri mai ridicate la 4 și 12 săptămâni decât după tratamentul cu doze mici ( P <0,001 pentru ambele).
Concentrațiile serice (ng / ml) de resveratrol după tratament. Modificarea (comparativ cu valoarea inițială-BL) a totalului trans (tRes; A) și cis (cRes; B) resveratrol, precum și resveratrol glucuronid (gRes; C) și sulfat (sRes; D) metaboliți 4 și 12 săptămâni după două ori -administrarea zilnică de placebo, doze mici sau mari de resveratrol. Sunt incluse bare de eroare standard. (*) deasupra barelor indică o modificare semnificativă față de valoarea inițială, P <0,05. Numărul de persoane din fiecare grup a fost după cum urmează: placebo, 9; doză mică de resveratrol, 12; și resveratrol în doze mari, 9.
Total cis- resveratrol (Fig. 1B) nu a fost detectabil în niciunul (0/32) din probele de ser inițiale și nici în probele la 4 și 12 săptămâni în grupurile placebo și cu doze mici de resveratrol. Nivelurile de cis- resveratrol au variat de la 0% –5,2% din resveratrolul total în grupul cu doze mari la 4 și 12 săptămâni. Nu am reușit să detectăm resveratrol gratuit în niciuna dintre probele de ser.
De asemenea, am măsurat metaboliții resveratrolului total și ai sulfatului total și, prin urmare, am calculat metaboliții glucuronidici totali în fiecare probă. Specia de glucuronide (Fig. 1C) cuprind 93% -100% din metaboliți, cu speciile de sulfat (Fig. 1D) cuprinzând restul. Creșterea metaboliților glucuronidici a fost similară cu cea a trans- resveratrolului după tratamentul cu resveratrol cu doze mici și mari.
PGE 2 nu a fost modificat semnificativ de doza sau de nivelul circulant de resveratrol
Am evaluat expresia PGE 2 în ambele NAF (Fig. 2A) și ser (Fig. 2B) înainte și după tratamentul cu resveratrol. Nivelurile PGE 2 nu s-au modificat semnificativ după 4 sau 12 săptămâni de tratament cu resveratrol fie în NAF, fie în ser.
Concentrațiile PGE 2 (ng / ml) după tratamentul cu resveratrol. Modificarea [comparativ cu valoarea inițială (BL)] a nivelului de lichid aspirat pentru mamelon (NAF; A) și a nivelului seric (B) de PGE 2 4 și 12 săptămâni după administrarea de două ori pe zi a placebo, resveratrol cu doze mici sau mari.
Modificarea metilării ADN este legată de concentrația de resveratrol
Am evaluat efectul dozei de resveratrol asupra modificării metilării ADN-ului (Fig. 3). Am putut măsura metilarea ADN-ului în cele 4 gene ( p16, CCND2, APC și RASSF-1a ) la momentul inițial și 12 săptămâni la 22/30 (73%) femei. Nici resveratrolul cu doze mici și nici mari nu a avut un efect semnificativ asupra oricăreia dintre cele 4 gene studiate. Cu toate acestea, fracția de ADN RASSF-1 α metilat a scăzut, în timp ce cea a APC a crescut pentru 3 din 4 femei după doze mari de resveratrol comparativ cu înainte de tratament. Datorită variabilității concentrației de resveratrol în rândul femeilor care primesc doze similare, am evaluat apoi schimbarea metilării pe baza concentrației de resveratrol în circulație.
Procent ADN metilat în genele supresoare tumorale după tratamentul cu resveratrol. Modificarea (comparativ cu valoarea inițială-BL) a procentului de metilare a p16 , APC, RASSF-1a și CCND2 12 săptămâni după administrarea de două ori pe zi a placebo, resveratrol cu doze mici sau mari.
Modificarea atât a resveratrolului seric, cât și a NAF PGE 2 Prezic schimbarea metilării RASSF-1 α
Am determinat modul în care nivelul de trans -resveratrol și resveratrol-glucuronid din ser s-a corelat cu modificarea fracției de ADN care a fost metilată. Nivelul nici unei specii nu s-a corelat cu modificarea metilării p16 , APC și CCND2 . Pe de altă parte, concentrația mai mare a ambelor specii s-a corelat cu o scădere mai mare a metilării α RASSF-1 ( P = 0,047) după 12 săptămâni de tratament (Fig. 4A).
Modificarea metilării RASSF-1a vs. schimbarea concentrației (A) trans- resveratrol și (B) NAF PGE 2 . A fost efectuată regresia liniară folosind cele mai mici pătrate pentru montarea modelului. Metilarea RASSF-1a a scăzut semnificativ odată cu creșterea concentrației trans- resveratrolului total ( P = 0,047, r 2 = -0,14) și cu scăderea PGE 2 ( P = 0,045, r 2 = 0,18).
De asemenea, am evaluat corelația dintre modificarea NAF și serul PGE 2 după tratament cu modificarea nivelului de ADN metilat pentru fiecare genă studiată. Modificarea PGE 2 serică nu a fost legată de modificarea metilării pentru niciuna dintre cele 4 gene. Modificarea NAF PGE 2 nu a fost legată de modificarea metilării p16, APC și nici CCND2 , în timp ce schimbarea fracției de ADN RASSF-1a metilat după tratament ( P = 0,045) s-a corelat pozitiv cu modificarea NAF PGE 2 (Fig. 4B).Mergi la:
DISCUŢIE
Am observat că specia predominantă de resveratrol în circulație a fost metabolitul glucuronid. Rapoartele sugerează că glucuronidul și majoritatea metaboliților sulfați au mai puțină activitate biologică decât compusul liber ( 36 ). Am proiectat studiul pe baza dovezilor că resveratrolul liber era prezent în țesuturi, cu metaboliții resveratrolului predominând în circulație. Probele MD și NAF nu au furnizat suficient material pentru analize de resveratrol. Deoarece investigația noastră a înscris persoane sănătoase, colectarea și analiza țesuturilor nu au fost fezabile. Un raport recent ( 37) care au tratat 20 de indivizi (10 per grup) cunoscuți a avea cancer colorectal cu 1 din 2 doze de resveratrol timp de 8 zile înainte de rezecția chirurgicală, a constatat că resveratrolul liber „a reprezentat o porțiune mult mai mare din totalul speciilor de resveratrol din țesutul colorectal decât în plasmă la un punct de timp echivalent postdozare. ” Acest raport a demonstrat, de asemenea, o scădere semnificativă a proliferării în țesutul colorectal după tratament. Raportul demonstrează că, deși conjugarea resveratrolului liber după ingestie duce la identificarea metaboliților ca specie predominantă în circulație, forma liberă a resveratrolului poate fi încă găsită la niveluri semnificative în țesuturile colorectale patologice și maligne ( 37 ). Deoarece bacteriile intestinale metabolizează resveratrolul ( 38), raportul fracționar al metaboliților poate varia între țesuturi intestinale și alte țesuturi.
Am raportat anterior un efect dependent de doză al trans-resveratrolului asupra expresiei genei ADN metitransferază ( 12 ) în celulele cancerului de sân in vitro. În prezentul raport, deși am observat o tendință nesemnificativă către metilarea RASSF-1a mai mică în funcție de doză, a existat o scădere semnificativă a metilării legate de nivelul circulant atât al trans- resveratrolului total, cât și al metabolitului său glucuronid (datele despre metaboliți nu sunt prezentate). Într-un raport anterior care a evaluat efectele chemopreventive ale polifenolilor din soia genisteină și daidzeină ( 8), am observat că nivelul circulant al genisteinei a fost un predictor mai bun al modificării metilării decât doza. Credem că acest lucru este valabil și pentru tratamentul cu polifenol resveratrol, probabil, deoarece rata la care compusul este metabolizat și eliminat variază de la individ la individ.
În studiile preclinice, s-a demonstrat că resveratrolul scade ciclooxigenaza (COX) -1 și -2 ( 39 ), ceea ce duce la o scădere a PGE 2 ( 40 ). Deși nu am observat o scădere a PGE 2 legată de doza sau nivelul circulant de trans- resveratrol sau metaboliții agentului, a existat o scădere semnificativă a expresiei PGE 2 în NAF în asociere cu o scădere a RASSF-1a (dar nu și a APC, p16, sau CCND2 ) metilare. Am raportat anterior că metilarea RASSF1a crește odată cu progresia bolii de la precancer la cancer în probele de țesut intraductal și de sân ( 22).). Scăderea metilării RASSF-1a și asocierea metilării scad cu scăderea PGE 2 sunt în concordanță cu efectele chemopreventive cunoscute ale resveratrolului.
Am observat o creștere dependentă de doză a trans- resveratrolului și a metabolitului glucuronid după ambele 4 și 12 săptămâni de tratament. Trans a fost izomerul resveratrol predominant în circulație, izomerul cis cuprinzând 5,2% sau mai puțin din totalul detectat. Specia serică predominantă, având în vedere resveratrolul liber și metaboliții, a fost metabolitul glucuronid, care cuprinde 93% -100% din totalul prezent în fiecare probă. Acest lucru este similar cu un raport anterior, după un tratament de scurtă durată, în care metabolitul glucuronid conținea 95% – 97% din speciile de resveratrol detectate în circulație. În acel studiu, resveratrolul liber nu a fost detectat în plasmă, dar a fost ușor detectat în țesutul colorectal benign și malign la aceiași subiecți ( 37). Atât resveratrolul liber, cât și metaboliții pot pătrunde în celulele din mai multe țesuturi ( 41 ). Bănuim că metaboliții glucuronid și sulfat sunt scindați intracelular de glucuronidaze și sulfataze la forma lor liberă în sân.
Un punct forte al studiului nostru este că, după cunoștințele noastre, acesta este primul raport care evaluează efectele dependente de doză ale resveratrolului asupra metilării ADN la un grup de indivizi cu risc crescut de cancer. Un al doilea punct forte a fost designul studiului, care a fost prospectiv, dublu-orb și controlat cu placebo. Limitările studiului au inclus dimensiunea modestă a eșantionului și faptul că nu am exclus de la înscriere persoanele care foloseau AINS. Deși utilizarea AINS nu pare să influențeze PGE 2 sau resveratrol, nu putem fi siguri că utilizarea AINS nu ne-a limitat capacitatea de a detecta un efect al resveratrolului asupra expresiei PGE 2 . De asemenea, nu putem exclude posibilitatea ca o doză mai mare de resveratrol să fi fost mai eficientă în scăderea PGE 2. Un studiu recent a indicat că dozele de 500 și 1000 mg pe zi au fost bine tolerate și au avut un efect antiproliferativ semnificativ asupra țesutului cancerului colorectal ( 34 ).
În concluzie, demonstrăm că izomerul trans și metabolitul glucuronid predomină în circulație după administrarea cronică de trans- resveratrol, cu o creștere dependentă de doză în ambele. Metilarea RASSF-1a a scăzut odată cu creșterea nivelului de trans -resveratrol și resveratrol-glucuronid în circulație și cu scăderea expresiei PGE 2 în sân. Observațiile noastre preliminare sugerează un mecanism nou pentru efectul chemopreventiv al trans- resveratrolului în sânul femeilor cu risc crescut.Mergi la:
CONFIRMARE
Acest studiu a fost finanțat parțial din grantul NIH CA124818.Mergi la:
Informații despre colaboratori
Weizhu Zhu, Departamentele de Chirurgie și Patologie, Universitatea din Dakota de Nord Școala de Medicină și Științe ale Sănătății, Grand Forks, Dakota de Nord, SUA.
Wenyi Qin, Departamentele de Chirurgie și Patologie, Universitatea din Dakota de Nord Școala de Medicină și Științe ale Sănătății, Grand Forks, Dakota de Nord, SUA.
Ke Zhang, Secția de biostatistică, Universitatea din Dakota de Nord Școala de Medicină și Științe ale Sănătății, Grand Forks, Dakota de Nord, SUA.
George E. Rottinghaus, Școala de Medicină Veterinară, Universitatea din Missouri, Columbia, Missouri, SUA.
Yin-Chieh Chen, Școala de Medicină Veterinară, Universitatea Missouri, Columbia, Missouri, SUA.
Beth Kliethermes, Departamentele de Chirurgie și Patologie, Universitatea din Dakota de Nord Școala de Medicină și Științe ale Sănătății, Grand Forks, Dakota de Nord, SUA.
Edward R. Sauter, Departamentele de Chirurgie și Patologie, Universitatea din Dakota de Nord Școala de Medicină și Științe ale Sănătății, Grand Forks, Dakota de Nord, SUA.Mergi la:
REFERINȚE
1. Subbaramaiah K, Chung WJ, Michaluart P, Telang N, Tanabe T și colab. Resveratrolul inhibă transcripția și activitatea ciclooxigenazei-2 în celulele epiteliale mamare umane tratate cu ester de forbol. J Biol Chem. 1998; 273 : 21875-21882. [ PubMed ] [ Google Scholar ]2. Roberti M, Pizzirani D, Simoni D, Rondanin R, Baruchello R și colab. Sinteza și evaluarea biologică a resveratrolului și a analogilor ca agenți inducători de apoptoză. J Med Chem. 2003; 46 : 3546–3554. [ PubMed ] [ Google Scholar ]3. Mgbonyebi OP, Russo J, Russo IH. Efect antiproliferativ al resveratrolului sintetic asupra celulelor epiteliale ale sânului uman. Int J Oncol. 1998; 12 : 865–869. [ PubMed ] [ Google Scholar ]4. Jang M, Cai L, Udeani GO, Slowing KV, Thomas CF și colab. Activitatea chimiopreventivă a cancerului a resveratrolului, un produs natural derivat din struguri. Ştiinţă. 1997; 275 : 218–220. [ PubMed ] [ Google Scholar ]5. Banerjee S, Bueso-Ramos C, Aggarwal BB. Suprimarea 7,12-dimetilbenz (a) carcinogenezei mamare induse de antracen la șobolani de către resveratrol: rolul factorului nuclear-kappaB, ciclooxigenazei 2 și metaloproteazei matricei 9. Cancer Res. 2002; 62 : 4945–4954. [ PubMed ] [ Google Scholar ]6. Levi F, Pasche C, Lucchini F, Ghidoni R, Ferraroni M, și colab. Risveratrol și risc de cancer mamar. Eur J Rac Prev. 2005; 14 : 139–142. [ PubMed ] [ Google Scholar ]7. Widschwendter M, Jones PA. Metilarea ADN și carcinogeneza sânilor. Oncogene. 2002; 21 : 5462-5482. [ PubMed ] [ Google Scholar ]8. Qin W, Zhu W, Shi H, Hewett JE, Ruhlen RL și colab. Izoflavonele din soia au un efect antiestrogen și modifică hipermetilarea promotorului mamar la femeile sănătoase în premenopauză. Cancerul Nutr. 2009; 61 : 238–244. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]9. Bestor TH. ADN metiltransferazele mamiferelor. Hum Mol Genet. 2000; 9 : 2395–2402. [ PubMed ] [ Google Scholar ]10. Leu YW, Rahmatpanah F, Shi H, Wei SH, Liu JC și colab. Interferența ARN dublu a DNMT3b și DNMT1 îmbunătățește demetilarea ADN și reactivarea genei. Cancer Res. 2003; 63 : 6110–6115. [ PubMed ] [ Google Scholar ]11. Rhee I, Bachman KE, Park BH, Jair KW, Yen RW și colab. DNMT1 și DNMT3b cooperează pentru a reduce la tăcere genele din celulele canceroase umane. Natură. 2002; 416 (6880): 552–556. [ PubMed ] [ Google Scholar ]12. Qin W, Zhu W, Sauter ER. Resveratrolul a indus metilarea ADN în cancerul de sân ER +. Proc Am Assn Cancer Res. 2005; 96 : 2750A. [ Google Scholar ]13. Walle T, Hsieh F, DeLegge MH, Oatis JE, Jr, Walle UK. Absorbție ridicată, dar biodisponibilitate foarte scăzută a resveratrolului oral la om. Eliminarea de droguri Metab. 2004; 32 : 1377–1382. [ PubMed ] [ Google Scholar ]14. de la Lastra CA, Villegas I. Resveratrolul ca agent antiinflamator și anti-îmbătrânire: mecanisme și implicații clinice. Mol Nutr Food Res. 2005; 49 : 405–430. [ PubMed ] [ Google Scholar ]15. Rowland I, Faughnan M, Hoey L, Wahala K, Williamson G și colab. Biodisponibilitatea fitoestrogenilor. Br J Nutr. 2003; 89 (Supliment 1): S45 – S58. [ PubMed ] [ Google Scholar ]16. Jannin B, Menzel M, Berlot JP, Delmas D, Lancon A și colab. Transportul resveratrolului, un agent chemopreventiv al cancerului, către ținte celulare: legarea proteinelor plasmatice și absorbția celulară. Biochem Pharmacol. 2004; 68 : 1113–1118. [ PubMed ] [ Google Scholar ]17. Wang LX, Heredia A, Song H, Zhang Z, Yu B și colab. Glucuronidele de resveratrol ca metaboliți ai resveratrolului la om: caracterizare, sinteză și activitate anti-HIV. J Pharm Sci. 2004; 93 : 2448–2457. [ PubMed ] [ Google Scholar ]18. Santner SJ, Feil PD, Santen RJ. Producția in situ de estrogen prin calea estronei sulfatazei în tumorile mamare: importanță relativă față de calea aromatazei. J Clin Endocrinol Metab. 1984; 59 : 29–33. [ PubMed ] [ Google Scholar ]19. Signorelli P, Ghidoni R. Resveratrolul ca nutrient anticancerigen: bază moleculară, întrebări deschise și promisiuni. J Nutr Biochem. 2005; 16 : 449–466. [ PubMed ] [ Google Scholar ]20. Colin D, Lancon A, Delmas D, Lizard G, Abrossinow J, și colab. Activitățile antiproliferative ale resveratrolului și ale compușilor înrudiți în celulele HepG2 derivate din hepatocite umane sunt asociate cu tulburări celulare biochimice relevate prin analize de fluorescență. Biochimie. 2008; 90 : 1674–1684. [ PubMed ] [ Google Scholar ]21. Delmas D, Lancon A, Colin D, Jannin B, Latruffe N. Resveratrolul ca agent chemopreventiv: o moleculă promițătoare pentru combaterea cancerului. Ținte de droguri Curr. 2006; 7 : 423-442. [ PubMed ] [ Google Scholar ]22. Zhu W, Qin W, Hewett JE, Sauter ER. Evaluarea cantitativă a hipermetilării ADN în țesutul și fluidele mamare maligne și benigne. Int J Rac. 2010; 126 : 474–482. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]23. Van der Auwera I, Van Laere SJ, Van den Bosch SM, Van den Eynden GG, Trinh BX și colab. Metilarea aberantă a promotorului genei adenomatoase de polipoză coli (APC) este asociată cu fenotipul inflamator al cancerului de sân. Br J Rac. 2008; 99 : 1735–1742. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]24. Sheng H, Shao J, Morrow JD, Beauchamp RD, DuBois RN. Modularea apoptozei și a expresiei Bcl-2 de către prostaglandina E2 în celulele cancerului de colon uman. Cancer Res. 1998; 58 : 362–366. [ PubMed ] [ Google Scholar ]25. Masferrer JL, Leahy KM, Koki AT, Zweifel BS, Settle SL, și colab. Activități antiangiogene și antitumorale ale inhibitorilor ciclooxigenazei-2. Cancer Res. 2000; 60 : 1306–1311. [ PubMed ] [ Google Scholar ]26. Fulton AM, Gimotty P, Alonsozana E, Dorsey R, Kundu N. Nivelurile ridicate de prostaglandină E2 (PGE2) în cancerul de sân la om sunt asociate cu o supraviețuire slabă pe termen lung. Proc Am Assn Cancer Res. 2000; 41 : 3660A. [ Google Scholar ]27. Chachay VS, Kirkpatrick CM, Hickman IJ, Ferguson M, Prins JB și colab. Resveratrol: pastile pentru a înlocui o dietă sănătoasă? Br J Clin Pharmacol. 2011; 72 : 27–38. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]28. Sauter ER, Daly M, Linahan K, Ehya H, Engstrom PF și colab. Nivelurile de antigen specific prostatei în lichidul aspirat pentru mamelon se corelează cu riscul de cancer mamar. Biomarkeri de epidemiol pentru cancer Prev. 1996; 5 : 967–970. [ PubMed ] [ Google Scholar ]29. Sauter ER, Ehya H, Klein-Szanto AJ, Wagner-Mann C, MacGibbon B. Rezultatele ductoscopiei fibro-optice la femeile cu și fără descărcare spontană a mamelonului. Cancer. 2005; 103 : 914-921. [ PubMed ] [ Google Scholar ]30. Gustafson KS, Furth EE, Heitjan DF, Fansler ZB, Clark DP. Profilarea metilării ADN a leziunilor intraepiteliale scuamoase cervicale folosind probe de citologie pe bază de lichide: o abordare care utilizează analiza caracteristică a receptorului. Cancer. 2004; 102 : 259–268. [ PubMed ] [ Google Scholar ]31. House MG, Guo M, Iacobuzio-Donahue C, Herman JG. Progresia moleculară a metilării promotorului în neoplasmele papilare mucinoase intraductale (IPMN) ale pancreasului. Carcinogeneză. 2003; 24 : 193–198. [ PubMed ] [ Google Scholar ]32. Fackler MJ, McVeigh M, Mehrotra J, Blum MA, Lange J și colab. Test de PCR cantitativ multiplex metilare specific pentru detectarea hipermetilării promotorului în gene multiple în cancerul de sân. Cancer Res. 2004; 64 : 4442–4452. [ PubMed ] [ Google Scholar ]33. Wang D, Hang T, Wu C, Liu W. Identificarea principalilor metaboliți ai resveratrolului în urina de șobolan prin HPLC-MS / MS. J Chromatogr B Analyt Technol Biomed Life Sci. 2005; 829 : 97–106. [ PubMed ] [ Google Scholar ]34. Yu C, Shin YG, Chow A, Li Y, Kosmeder JW și colab. Metabolizarea resveratrolului la om, șobolan și șoarece. Pharm Res. 2002; 19 : 1907–1914. [ PubMed ] [ Google Scholar ]35. Sauter ER, Qin W, Schlatter L, Hewett JE, Flynn JT. Celecoxibul scade concentrațiile de prostaglandină E2 în lichidul aspirat pentru mamelon de la femeile cu risc crescut postmenopauză și femeile cu cancer de sân. Cancer BMC. 2006 [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]36. Delmas D, Aires V, Limagne E, Dutartre P, Mazue F și colab. Transportul, stabilitatea și activitatea biologică a resveratrolului. Ann NY Acad Sci. 2011; 1215 : 48–59. [ PubMed ] [ Google Scholar ]37. Patel KR, Brown VA, Jones DJ, Britton RG, Hemingway D și colab. Farmacologia clinică a resveratrolului și a metaboliților săi la pacienții cu cancer colorectal. Cancer Res. 2010; 70 : 7392–7399. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]38. Rodrigo R, Miranda A, Vergara L. Modularea sistemului antioxidant endogen de polifenoli ai vinului în bolile umane. Clin Chim Acta. 2011; 412 : 410–424. [ PubMed ] [ Google Scholar ]39. Murias M, Handler N, Erker T, Pleban K, Ecker G și colab. Analogii resveratrolului ca inhibitori selectivi ai ciclooxigenazei-2: sinteză și relație structură-activitate. Bioorg Med Chem. 2004; 12 : 5571–5578. [ PubMed ] [ Google Scholar ]40. Dave M, Attur M, Palmer G, Al-Mussawir HE, Kennish L, și colab. Resveratrolul antioxidant protejează împotriva apoptozei condrocitelor prin efecte asupra polarizării mitocondriale și a producției de ATP. Artrita reumatică. 2008; 58 : 2786-2797. [ PubMed ] [ Google Scholar ]41. Juan ME, Maijo M, Planas JM. Cuantificarea trans-resveratrolului și a metaboliților săi în plasma și țesuturile de șobolan prin HPLC. J Pharm Biomed Anal. 2010; 51 : 391-398. [ PubMed ] [ Google Scholar ]
Studiu pilot dublu-orb randomizat de fază I a resveratrolului micronizat (SRT501) la pacienții cu metastaze hepatice – siguranță, farmacocinetică și farmacodinamică
Ultima versiune editată a editorului a acestui articol este disponibilă gratuit la Cancer Prev Res (Phila
Abstract
Resveratrolul fitochimic a fost supus unei ample investigații preclinice pentru proprietățile sale chimiopreventive. Disponibilitatea sistemică scăzută a compusului părinte datorită metabolismului rapid și extins poate confunda utilitatea acestuia ca agent potențial pentru a preveni malignitățile în organele aflate la distanță de locul absorbției. Micronizarea permite absorbția crescută a medicamentului, crescând astfel disponibilitatea. Aici descriem un studiu pilot al SRT501, resveratrol micronizat, administrat zilnic la 5,0 g timp de 14 zile, pacienților cu cancer colorectal și metastaze hepatice programate să facă hepatectomie. Scopul studiului a fost de a evalua siguranța, farmacocinetica și farmacodinamica formulării. SRT501 sa dovedit a fi bine tolerat. Nivelurile medii de resveratrol plasmatic după o singură doză de administrare SRT501 au fost 1942 ± 1422 ng / ml,depășind cele publicate pentru doze echivalente de resveratrol non-micronizat de 3,6 ori. Resveratrolul a fost detectabil în țesutul hepatic după administrarea SRT501 (până la 2287 ng / g). Caspaza-3 scindată, un marker al apoptozei, a fost semnificativ crescută cu 39% în țesutul hepatic malign după tratamentul cu SRT501, comparativ cu țesutul de la pacienții tratați cu placebo. SRT501 justifică explorarea clinică suplimentară pentru a evalua utilitatea sa clinică potențială.SRT501 garantează explorarea clinică suplimentară pentru a evalua utilitatea sa clinică potențială.SRT501 garantează explorarea clinică suplimentară pentru a evalua utilitatea sa clinică potențială.
Introducere
Resveratrolul (3,5,4′-trihidroxi- trans -stilben ) este o fitoalexină produsă de anumite plante atunci când este afectată de agenți patogeni precum bacteriile sau ciupercile. Apare în pielea strugurilor roșii și în arahide și a fost investigată pe larg în ceea ce privește proprietățile sale chimiopreventive. Se consideră că resveratrolul mediază, cel puțin parțial, acțiunile presupuse de prevenire a bolilor vinului roșu și este, de asemenea, considerat a preveni bolile cardiovasculare și morbiditatea asociate cu obezitatea și bătrânețea [ 1 – 3 ]. Numeroasele proprietăți farmacologice ale resveratrolului și sugestia că acesta poate fi un mimetic cu restricție de calorii care poate prelungi viața, a generat un interes considerabil de către presa laică. O abundență de date obținute în celulein vitro și în modelele preclinice a evidențiat o mulțime de mecanisme potențiale prin care resveratrolul poate preveni afecțiunile maligne, predominant cele germane până la supraviețuirea și proliferarea celulară [ 4 ], inclusiv apoptoza dependentă de p53 [ 5 ] și oprirea ciclului celular [ 6 ]. Cu toate acestea, datele clinice solide rămân rare. Recente studii pilot clinice de resveratrol, in care a fost administrat în doze unice sau repetate de până la 5,0 g pe zi, axat pe siguranța sa, farmacocinetica (PK) proprietăți și analiza biomarkeri potential de eficacitate [ 4 , 7 – 13 ]. Profilurile PK ale resveratrolului la voluntari sănătoși au dezvăluit un metabolism rapid și extins la resveratrol-4′- O-glucuronid, resveratrol-3′- O -glucuronid și resveratrol-3- O- sulfat după administrarea orală a unei singure [ 8 ] sau a unor doze zilnice repetate [ 9 ] de 0,5 până la 5,0 g. În ciuda detectării concentrațiilor relativ mari de resveratrol în țesutul colorectal al pacienților, disponibilitatea sa sistemică este sever redusă prin metabolismul său avid. În consecință, deși resveratrolul a demonstrat activitate împotriva unei largi varietăți de tumori maligne in vitro (inclusiv intestin, sân, vezică, prostată, ficat și tiroidă) și in vivo (revizuite în [ 3 , 14 – 16]), este probabil ca o biodisponibilitate slabă să limiteze eficacitatea la locurile îndepărtate de tractul GI. În ciuda acestui fapt, există dovezi care sugerează că resveratrolul produce efecte chimiopreventive și terapeutice în modelele preclinice de cancer hepatic (revizuite în [ 17 ]), făcând aluzie la potențialul de eficacitate împotriva hepatocarcinogenezei și a tumorilor maligne care metastazează preferențial ficatul.
Datorită solubilității apoase slabe prezentate de resveratrol, absorbția digestivă este foarte influențată de rata de dizolvare a medicamentului. Într-un efort de a crește absorbția în tractul gastro-intestinal și, astfel, compusul părinte disponibil sistemic, a existat un interes considerabil în manipularea farmaceutică a resveratrolului. Scăderea dimensiunii particulelor acestor substanțe chimice poate îmbunătăți rata de dizolvare a acestora și, astfel, absorbția lor [ 18]. Prin urmare, scopul acestui studiu clinic a fost să investigheze dacă consumul de SRT501, o formulare micronizată de resveratrol proiectată de Sirtris, o companie GSK (Cambridge, MA, SUA) este sigur și generează niveluri măsurabile și farmacologic active ale agentului părinte în circulație și în ficat. În prezent, nu există date clinice disponibile care să determine concentrațiile de resveratrol în țesuturile umane, altele decât colonul. În acest scop, pacienții cu cancer colorectal destinat să fie supuși rezecției metastazelor hepatice au fost recrutați într-un studiu pilot. Au ingerat fie placebo, fie SRT501 în 14 zile consecutive înainte de operație, iar nivelurile de resveratrol au fost cuantificate în sânge și în țesutul de rezecție hepatică.Efectele farmacodinamice potențiale ale SRT501 au fost investigate prin compararea expresiei / activării biomarkerilor proteici candidați asociați intrinsec cu supraviețuirea celulară și apoptoza în circulația și țesutul pacienților care au primit medicamentul și a celor care au fost tratați cu placebo.Mergi la:
Metode
Formulare de resveratrol
Resveratrolul microparticular cu dimensiunea particulelor <5 μm (SRT501) a fost fabricat de Sirtris Pharmaceuticals, o companie GSK. Se consideră că reducerea dimensiunii particulelor îmbunătățește biodisponibilitatea resveratrolului, în principal datorită creșterii suprafeței și a proprietăților îmbunătățite ale suspensiei. Un plic conținând SRT501 (5,0 g) a fost amestecat cu o alicotă (4 ml) de soluție de docusat de sodiu (docusat de sodiu, acid citric, colorant, glicerină, aromatizant, citrat de sodiu, zaharină sodică și sorbitol) formată până la 20 ml în apă distilată formând o suspensie uniformă și ingerată de pacienți.
Pacienți
Studiul pilot, proiectat și sponsorizat de Sirtris, a fost realizat la Spitalele Universitare din Leicester (UHL) NHS-Trust (UHL 10507, registru de probă: NCT00920803). Acesta a fost revizuit și aprobat de Leicestershire, Northamptonshire și Rutland Research Ethics Committee (UK) și desfășurat în conformitate cu liniile directoare aplicabile privind bunele practici clinice. Nouă subiecți (cu vârsta de 18 ani sau peste) care prezentau cancer colorectal confirmat în stadiul IV și metastaze hepatice, care nu primiseră intervenție terapeutică pentru cancerul lor în decurs de 6 săptămâni de la începerea studiului și aveau o speranță de viață> 3 luni, au fost recrutați în studiu. Toți pacienții urmau să fie supuși rezecției metastazelor hepatice. Participanții trebuiau să fie capabili fizic să respecte protocolul și aveau un ECG normal și nu aveau antecedente de HIV sau hepatită B / C.
Pacientul a fost rugat să se abțină de la cantități mari de alimente și băuturi care conțin resveratrol, cum ar fi arahide, struguri, dud și alcool, în 48 de ore de la zilele programate de colectare a PK și în ziua rezecției chirurgicale.
Design de studiu
Studiul a fost un studiu clinic de fază I, randomizat (2: 1), dublu-orb. Șase subiecți au fost randomizați pentru a primi SRT501 și trei placebo (dioxid de titan). Au fost furnizate intervenții în truse clinice sub formă de pulbere și reconstituite așa cum s-a descris mai sus. Pacienții au ingerat formularea preoperatorie zilnic timp de minimum 10 și maximum 21 de zile, în funcție de programarea chirurgicală. Probele de plasmă pentru evaluarea PK au fost obținute în zilele 1 și 2 și în ziua rezecției chirurgicale. Plasma pentru evaluarea PD a fost obținută pre-doză, imediat înainte și în timpul intervenției chirurgicale. Țesutul hepatic adiacent bolnav și normal a fost rezecat la 6-7 ore după ultima doză. Participanții au completat un jurnal de evenimente adverse zilnic (AE) care a fost evaluat folosind criteriile de toxicitate comună NCI v3. Dintre cei nouă pacienți recrutați, șapte au finalizat studiul,unul a murit din cauza complicațiilor postoperatorii și unul a încetat administrarea devreme din cauza EA (diaree). Probele PK au fost obținute de la toți cei nouă subiecți. Obiectivele principale ale studiului au fost determinarea siguranței și tolerabilității SRT501 și farmacocineticii resveratrolului. Obiectivele secundare au fost explorarea potențialelor modificări farmacodinamice provocate de SRT501 în sânge și țesuturi.
Colectie de mostre
Probele de sânge au fost colectate înainte de administrare în ziua 1, pentru PK la 0,25, 0,5, 1, 2, 4, 8, 24 de ore după prima doză și înainte și în timpul intervenției chirurgicale. Plasma (tuburile de heparină de sodiu) și serul au fost obținute și depozitate la -80 ° C până la analiză. Țesuturile au fost recoltate din materialul rezecat, o parte din care a fost fixată în formalină și încorporată în parafină și o parte a fost înghețată imediat și depozitată la -80 ° C până la analiză.
Pregătirea și analiza probelor
Cuantificarea resveratrolului în plasmă și țesuturi a fost efectuată de Laboratoarele Charles River, Montreal, Canada. Metodologia a fost conformă cu reglementările de bună practică de laborator ale Administrației SUA pentru Alimente și Medicamente (21CFR partea 58), utilizând o metodă internă validată de cromatografie lichidă validată în tandem-spectrometrie de masă (LC-MS / MS). Pe scurt, 50 μL probe de plasmă au fost extrase folosind metanol acidificat (acid ascorbic: metanol: acid formic, 50: 50: 0,025) (5 μL) conținând resveratrol – 13 C 6ca standard intern. Probele au fost apoi centrifugate (14000 rpm / 0 ° C / 10 min) și supernatantul a fost alicotat într-o placă de colectare cu 96 de godeuri pentru analiză. Separarea, cuantificarea și caracterizarea resveratrolului au fost realizate folosind un sistem Symbiosis Pharma (metoda XLC) împreună cu un spectrometru de masă MDS Sciex API 3000. O coloană Waters YMC ODS-AQ (3 μm, 4,6 × 50 mm) a fost legată de un filtru de precolumnă Peek 0,5 μm. Eluantul a constat dintr-o fază mobilă binară (A; apă: acid formic, 100: 0,025 (v / v), B; metanol: acetonitril: acid formic, 20: 80: 0,025 (v / v / v)). Gradientul de eluare a fost A85% B15% până la A5% B95% timp de 4 minute. Resveratrolul a fost caracterizat prin spectrometrie de masă cu electrospray (modul de ionizare negativă) cu tensiune de pulverizare ionică -4500 V, potențial de declustrare -25 V, potențial de focalizare -30 V, potențial de intrare -12 V,energie de coliziune −27 V, potențial de ieșire de coliziune 40 V și temperatură 550 ° C. Limita inferioară de cuantificare (LLOQ) a fost de 5 ng / ml și limita superioară de cuantificare (ULOQ) 1000 ng / ml.
Probele de țesut au fost tocate pe gheață și s-au folosit ~ 100 mg pentru analiză. Metanol acidificat (acid ascorbic: metanol: acid formic, 50: 50: 0,025) (5 μL) conținând resveratrol – 13 C 6ca un standard intern, s-au adăugat la fiecare probă matricele de 100 μL matrice verzi și 500 μL etanol, amestecul a fost rotit și lăsat pe gheață (10 min). Probele au fost omogenizate (Fastprep, 2,4-5,5 m / s, 20 sec), lăsate pe gheață (10 min) și omogenizarea repetată înainte de centrifugare (14000 rpm / 0 ° C / 10 min). O alicotă (100 μL) de supernatant a fost adăugată la 100 μL apă: acid formic (100: 0,25). Separarea, cuantificarea și caracterizarea resveratrolului au fost realizate folosind un cromatograf Agilent 1100 cu un spectrometru de masă MDX Sciex API 5000. O coloană Waters YMC ODS-AQ (3 μm, 4,6 × 50 mm) a fost legată de un filtru de precolumnă Peek de 0,5 μm. Eluantul a fost ca pentru analiza plasmatică; eluarea în gradient a fost de peste 6,5 minute de la A70% B30% la A0% B100%, apoi înapoi la A70% B30%. Condițiile MS au fost cele de mai sus, cu excepția: tensiunea de pulverizare ionică -4500 V,potențial de decliniere −120 V, potențial de intrare −10 V, energie de coliziune −27 V, potențial de ieșire de coliziune −10 V și temperatură 700 ° C. Pentru probele de țesut, LLOQ a fost stabilit la 2,5 ng / g și ULOQ la 500 ng / g. Colectarea datelor a fost efectuată utilizând versiunea Analyst 1.4.2. (MDX Sciex).
Analiza proliferării celulare și a apoptozei
Țesuturile fixate pe formalină au fost analizate pentru markeri de proliferare și apoptoză folosind sistemul NovoLink Polymer Detection (Novocastra Laboratories, Newcastle-upon-Tyne, Marea Britanie) împreună cu anti-Ki-67 (Dako, Cambridgeshire, Marea Britanie) și anti-clivat- anticorpii caspase-3 (Cell Signaling Technology, MA, SUA). Pe scurt, secțiunile încorporate în parafină au fost decerate la 65 ° C și rehidratate printr-o serie de spălări cu alcool. Recuperarea antigenului a fost realizată prin microunde fie în Tris-EDTA (pH 9,0) pentru Ki-67, fie tampon citrat (pH 6,0) pentru caspaza-3 scindată. Secțiunile au fost vizualizate la mărire x40 folosind un microscop cu lumină inversată Leica DC300 și un sistem de cameră. Imaginile au fost procesate utilizând Adobe Photoshop v7.0.1. Celulele de colorare pozitive au fost marcate pe 10 câmpuri vizuale aleatorii pe fiecare lamă și au fost exprimate ca procente de celule colorate pozitiv.Toate analizele au fost efectuate orbite.
Analiza PGE2, VEGF și IGF-1
Testele ELISA au fost întreprinse pentru a evalua nivelurile de VEGF (Invitrogen, Paisley, Marea Britanie) în ser, nivelurile PGE2 (Cayman Chemicals, Cambridge Marea Britanie) în plasmă și nivelurile IGF-1 (R&D Systems, Oxford, Marea Britanie) în extractele de țesut hepatic. Testele au fost efectuate conform instrucțiunilor producătorilor și plăcile ELISA analizate folosind un cititor de plăci Fluostar Optima (BMG Labtech, Aylesbury, Marea Britanie)
analize statistice
Analizele statistice pentru datele farmacocinetice au inclus regresia liniară cu 1 / ponderarea concentrației și statistici descriptive; mijloacele aritmetice și abaterile standard, acuratețea și precizia au fost calculate folosind Watson Laboratory Information Management System (LIMS) (versiunea 7.2.0.02) și Microsoft Excel (versiunea 2000/2003).
Analizele statistice pentru datele farmacodinamice au fost efectuate folosind SPSS 16.0 pentru Windows. Mijloacele au fost comparate printr- un singur sens, ANOVA unilateral, urmat de testul post hoc al lui Tukey. O valoare p mai mică de 0,05 a fost considerată semnificativă.Mergi la:
Rezultate
Siguranța SRT-501
Nouă pacienți au fost recrutați în studiu (vezi datele demografice tabelul 1). Șase persoane au ingerat 5,0 g de SRT501 și trei au primit placebo, zilnic, timp de aproximativ 14 zile înainte de operație. Un pacient cu SRT501 a încetat administrarea în ziua 13. AE posibil sau probabil atribuibile aportului de agenți sunt prezentate înmasa 2. Au fost în primul rând de natură gastro-intestinală, incluzând greață și diaree, și de grad ușor (gradul 1 NCI CTC v3.0). Alte EA au inclus frisoane, letargie, erupții cutanate, iritații ale pielii și înroșirea vasculară, care s-au rezolvat fără sechele, cu excepția unui caz de letargie în grupul placebo, care era în curs la urmărire. Un pacient a dezvoltat peritonită postoperatorie și insuficiență hepatică și a murit în timpul urmăririi. Investigatorul principal a considerat că acest eveniment advers grav (SAE) nu are legătură cu studiul medicamentului.
tabelul 1
Date demografice pentru pacienții randomizați la 5,0 g / zi SRT501 și placebo
SRT501 (n = 6)
Placebo (n = 3)
Vârsta (ani)
68,5 (± 10,8)
64,3 (± 6,35)
Gen
M: 5
M: 1
F: 1
F: 2
IMC
28,10 (± 4,35)
26,87 (± 1,61)
Cursa (100%)
Caucazian (100%)
caucazian
Zile de studiu
13,3 (± 1,03)
14,0 (± 0,00)
Doza cumulativă pe subiect (g)
66,7 (± 5,16)
70,0 (± 0,00)
IMC: indicele de masă corporală; F: femeie; M: bărbat.
masa 2
Listări de evenimente adverse considerate posibil sau probabil legate de intervenția SRT501 / placebo.
Eveniment advers
SRT501
Placebo
(n = 6)
(n = 3)
Nr de evenimente Nr de pacienți
Nr de evenimente Nr de pacienți
Tulburări gastrointestinale:
12
5
2
1
Prurit anal
1
1
0
0
Diaree
7
5
2
1
Greaţă
4
1
0
0
Tulburări generale:
1
1
0
0
Frisoane
1
1
0
0
Tulburări ale sistemului nervos:
1
1
1
1
Letargie
0
0
1
1
Neuropatie periferica
1
1
0
0
Tulburări cutanate / tisulare:
2
2
0
0
Eczemă
1
1
0
0
Iritatie de piele
1
1
0
0
Tulburări vasculare:
1
1
0
0
Flushing
1
1
0
0
Farmacocinetica resveratrolului și concentrația în țesutul hepatic
Plasma pacienților a fost analizată prin HPLC-MS / MS pentru resveratrol. Concentrațiile de resveratrol au fost sub limitele inferioare de cuantificare (LLOQ) pentru toate probele de la subiecții cărora li s-a administrat placebo și măsurabile la pacienții care au primit SRT501. Parametrii farmacocinetici pentru resveratrol sunt indicați întabelul 3, și concentrațiile plasmatice versus curbele de timp pentru subiecții individuali dinfigura 1. Nivelurile C max au fost atinse 2,8 ore după administrarea dozei, iar concentrația plasmatică maximă medie a fost de 1942 ng / ml (8,51 nmol / ml). Timpul mediu de înjumătățire plasmatică prin eliminare a plasmei a fost de puțin peste 1 oră.figura 1
Concentrația plasmatică a resveratrolului la pacienții individuali care au primit o doză (5,0 g) de SRT501. Analiza a fost efectuată prin HPLC / MS-MS, pentru detalii vezi Metode.
Tabelul 3
Statistici descriptive pentru farmacocinetica resveratrolului din plasma pacienților cărora li se administrează SRT501. Toate probele de la pacienții randomizați la grupul placebo au fost sub limita de cuantificare în toate momentele (datele nu sunt prezentate).
Cmax (ng / mL)
tmax (hr)
t 1/2 (oră)
ASC (0-24 ore) (ng / oră / ml)
n
6
6
3
3
Media geometrică
1942.00
2,80
1,06
6327,40
SD
1422,62
1.10
0,39
2247,20
Gamă
896 – 4890
2.0 – 4.0
0,81 – 1,54
5030 – 9140
Cmax: concentrația plasmatică maximă; tmax: timpul de concentrație plasmatică maximă; t 1/2 : timpul de înjumătățire plasmatică prin eliminare aparent; ASC: zona sub curba concentrației plasmatice față de timp de la 0 la 24 de ore.
Resveratrolul a fost cuantificat în țesuturile hepatice tumorale și adiacente normale. Nivelurile de resveratrol au fost sub LLOQ la toți subiecții tratați cu placebo și la unul dintre cei șase pacienți cu SRT501. Nivelurile medii de resveratrol la restul de cinci pacienți cărora li s-a administrat SRT501 au fost 1098 ± 1393 ng / g (4,81 nmol / g, interval 52-2834 ng / g) și 420 ± 341 ng / g (1,84 nmol / g, interval 46-914 ng / g) în tumoră și, respectiv, în țesutul normal.
Efectul SRT501 asupra farmacodinamicii
Efectele potențiale ale SRT501 asupra proceselor relevante pentru supraviețuirea celulară și apoptoză au fost măsurate în plasmă și țesut. Nu a existat nicio diferență între pacienții cărora li s-a administrat placebo sau SRT501 în ceea ce privește nivelurile plasmatice / serice de prostaglandină E2 (PGE-2) și factorul de creștere endotelial vascular (VEGF) (nereprezentat). Probele de țesut au fost analizate prin imunohistochimie, ELISA sau western blot pentru modificările induse de SRT501 la nivelurile de IGF-1, Ki67, phospho Akt (ser473), Akt1, phospho-GSK3, GSK3, phospho ERK, ERK, phospho-JNK, JNK, beta-catenin, survivin, BCL2, Bax sau PARP. În toate cazurile nu au existat diferențe semnificative între placebo și SRT501 (datele nu sunt prezentate). Apoptoza, reflectată de imunohistochimie pentru caspaza 3 clivată în țesutul tumoral, a crescut semnificativ cu 39% (până la 1,44% celule apoptotice totale, p = 0.038) la pacienții cu SRT501 comparativ cu cei care au luat placebo (figura 2).Figura 2
Apoptoza reflectată de caspaza-3 scindată (A, fotomicrografii reprezentative, B, cuantificarea fotomicrografiilor) în țesuturile hepatice normale și tumorale ale pacienților care au consumat SRT501 ( N = 6, bare negre) sau placebo ( N = 3, bare albe) zilnic pentru 14 zile. Valorile sunt media +/− SD. (a) reprezintă o diferență semnificativă între țesutul normal și țesutul tumoral pentru grupul SRT501. (b) reprezintă o diferență semnificativă între țesutul normal și cel tumoral pentru grupul placebo. (c) reprezintă o diferență semnificativă între SRT501 și țesutul tumoral placebo. ( P = <0,05, ANOVA, urmată de testul post hoc al lui Tukey).Mergi la:
Discuţie
Acest studiu reprezintă prima descriere a parametrilor farmacocinetici pentru resveratrol după ingestia SRT501, resveratrol micronizat, la subiecții cu cancer colorectal. Doza de resveratrol utilizată (5,0 g) este echivalentă cu unul dintre nivelurile de doză la care resveratrolul non-micronizat a fost administrat recent voluntarilor sănătoși [ 9]]. În studiul voluntar sănătos, zece subiecți pe 5,0 g de resveratrol non-micronizat au raportat douăzeci de AE posibil sau probabil legate de ingestia de capsule de resveratrol, care se compară cu șaptesprezece AE raportate pentru șase pacienți care au primit SRT501. Prin urmare, cele două formulări par să provoace o incidență similară a EA, deși în studiul anterior resveratrolul non-micronizat a fost administrat pe o durată mai lungă (29 de zile) decât SRT501. Toate AE raportate după administrarea SRT501 au fost ușoare, în timp ce doi dintre cei zece voluntari sănătoși pe resveratrol non-micronizat, au prezentat moderat și unul cu AE gastrointestinale severe [ 9]]. Această comparație oferă posibilitatea ca SRT501 formulat ca suspensie să fie mai bine tolerat decât resveratrolul nemicronizat administrat ca zece capsule de 0,5 g pe zi. Deoarece se aștepta ca resveratrolul micronizat să permită concentrații sistemice mai ridicate [ 19 ], această constatare sugerează, de asemenea, că ingestia unui număr mare de capsule sau prezența resveratrolului neabsorbit în tractul gastro-intestinal poate fi responsabilă pentru efectele adverse, mai degrabă decât pentru efectul real. concentrațiile plasmatice atinse.
Analiza farmacocinetică a relevat că media Cmax plasmatică medie pentru resveratrol măsurată după ingestia SRT501 (medie 1942 ng / ml, CV: 73,3%) a fost de 3,6 ori mai mare decât cea obținută în urma unei doze unice de resveratrol non-micronizat la voluntarii sănătoși ( medie 538.8 ng / ml (CV: 72,5%) [ 8 ] . , în concordanță cu biodisponibilitatea superioară a resveratrol atunci când este administrat ca SRT501 comparativ cu agentul non-micronizate timpul necesar pentru a ajunge la C max pentru resveratrolul a fost de 2,8 ore în cazul SRT501 în comparație cu 1,5 h pentru resveratrol non-micronizat [ 8 ].
Concentrațiile de resveratrol obținute în metastazele hepatice după administrarea SRT501 (0,22-12,4 μM) au fost de ordinul mărimii observate pentru a provoca efect farmacologic în celulele cancerului colorectal uman in vitro și în modelele preclinice in vivo . Această interpretare este coroborată de constatarea că SRT501 a cauzat o creștere mică, dar semnificativă, a imunoractivității caspazei-3 clivate în țesutul tumoral, în comparație cu țesutul echivalent de la subiecții tratați cu placebo. Deși nu am putut confirma aceste efecte prin intermediul altor markeri farmacodinamici, acest lucru se poate datora sensibilității scăzute a tehnicilor utilizate. Mecanisme prin care au fost observate concentrații scăzute de resveratrol pentru a induce efectul apoptotic și anti-proliferativ in vitroinclud inducerea redistribuirii Fas și asocierea acestuia cu complexul de semnalizare inducătoare de moarte (DISC) [ 20 ] și prin inhibarea semnalizării wnt și scăderea ulterioară a localizării nucleare a β-cateninei [ 21 ]. Într-un model preclinic de hepatocarcinogeneză inițiată de dietilnitrosamină, apoptoza semnificativă mediată de resveratrol a rezultat probabil din creșterea expresiei Bax și, prin urmare, din creșterea raportului Bax: Bcl2 [ 22 ]. În timp ce nivelurile de resveratrol tisular nu au fost determinate în acest studiu, extrapolarea datelor din studiile PK pe rozătoare [ 23 , 24] ar sugera concentrațiile hepatice de resveratrol necesare pentru inducerea apoptozei, să fie doar moderat mai mari decât maximul realizat în studiul SRT501 descris aici.
Aceasta este prima demonstrație că resveratrolul poate atinge concentrații potențial active în țesuturile umane care sunt îndepărtate de tractul GI.
În rezumat, rezultatele acestui studiu pilot al SRT501 permit trei concluzii: i. consumul său zilnic timp de 14 zile pare a fi bine tolerat la pacienții cu cancer colorectal, ii. C max pentru SRT501 a fost mai mare decât cel raportat pentru doza echivalentă de resveratrol non-micronizat, iii. ingestia sa a furnizat niveluri de resveratrol măsurabile într-un țesut îndepărtat de tractul gastro-intestinal (în special ficatul), iar aceste concentrații au fost însoțite de un efect farmacologic semnificativ.
Resveratrolul micronizat, după cum este exemplificat de SRT501, justifică explorarea clinică suplimentară în cohorte mai mari de subiecți, pentru a testa ipoteza că resveratrolul poate fi util ca agent chimiopreventiv sau chimioterapeutic al cancerului.Mergi la:
Mulțumiri
Sprijinul acordat : Această lucrare a fost susținută de Cancer Research UK în cadrul unui program de finanțare [C325 / A6691], de Cancer Research UK în colaborare cu Departamentul de Sănătate din Marea Britanie pentru o subvenție a Centrului de Medicină a Cancerului Experimental [C325 / A7241] și de Sirtris, A Compania GSK.
Sprijin financiar : Această lucrare a fost susținută de Cancer Research UK pe o subvenție de program [C325 / A6691], de Cancer Research UK în colaborare cu Departamentul de Sănătate din Marea Britanie cu privire la o subvenție experimentală a Centrului de Medicină a Cancerului [C325 / A7241] și de Sirtris, A Compania GSK.Mergi la:
Note de subsol
Conflict de interese: Acest studiu a fost sponsorizat și parțial finanțat de Sirtris, o companie GSK.Mergi la:
Referințe
[1] Barger JL, Kayo T, Vann JM, Arias EB, Wang J, Hacker TA și colab. O doză mică de resveratrol alimentar imită parțial restricția calorică și întârzie parametrii de îmbătrânire la șoareci. Plus unu. 2008; 3 : e2264. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ][2] Baur JA, Sinclair DA. Potențialul terapeutic al resveratrolului: dovezi in vivo. Nat Rev Drug Discov. 2006; 5 : 493-506. [ PubMed ] [ Google Scholar ][3] Un Bishayee. Prevenirea cancerului și tratamentul cu resveratrol: de la studii de rozătoare la studii clinice. Cancer Prev Res (Phila) 2009; 2 : 409-18. [ PubMed ] [ Google Scholar ][4] Shankar S, Singh G, Srivastava RK. Chimioprevenție prin resveratrol: mecanisme moleculare și potențial terapeutic. Biosci din față. 2007; 12 : 4839–54. [ PubMed ] [ Google Scholar ][5] Ea QB, Bode AM, Ma WY, Chen NY, Dong Z. Activarea p53 și apoptoza indusă de resveratrol este mediată de protein kinaze și p38 kinază reglate de semnal extracelular. Cancer Res. 2001; 61 : 1604–10. [ PubMed ] [ Google Scholar ][6] Joe AK, Liu H, Suzui M, Vural ME, Xiao D, Weinstein IB. Resveratrolul induce inhibarea creșterii, stoparea fazei S, apoptoza și modificări ale expresiei biomarkerului în mai multe linii celulare de cancer uman. Clin Cancer Res. 2002; 8 : 893–903. [ PubMed ] [ Google Scholar ][7] Almeida L, Vaz-da-Silva M, Falcao A, Soares E, Costa R, Loureiro AI, și colab. Profilul farmacocinetic și de siguranță al trans-resveratrolului într-un studiu în creștere cu doze multiple la voluntari sănătoși. Mol Nutr Food Res. 2009; 53 (Supliment 1): S7-15. [ PubMed ] [ Google Scholar ][8] Boocock DJ, Faust GE, Patel KR, Schinas AM, Brown VA, Ducharme MP și colab. Studiu farmacocinetic de creștere a dozei de fază I la voluntari sănătoși ai resveratrolului, un potențial agent chimiopreventiv al cancerului. Biomarkeri de epidemiol pentru cancer Prev. 2007; 16 : 1246–52. [ PubMed ] [ Google Scholar ][9] Brown VA, Patel KR, Viskaduraki M, Crowell JA, Perloff M, Booth TD și colab. Studiul repetat al dozei de agent chimiopreventiv al cancerului Resveratrol la voluntari sănătoși: siguranță, farmacocinetică și efect asupra axei factorului de creștere asemănător insulinei. Cancer Res. 2010; 70 : 9003-11. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ][10] la Porte C, Voduc N, Zhang G, Seguin I, Tardiff D, Singhal N și colab. Farmacocinetica stării de echilibru și tolerabilitatea trans-resveratrolului 2000 mg de două ori pe zi cu alimente, quercetină și alcool (etanol) la subiecți umani sănătoși. Clin Pharmacokinet. 2009; 49 : 449–54. [ PubMed ] [ Google Scholar ][11] Nunes T, Almeida L, Rocha JF, Falcao A, Fernandes-Lopes C, Loureiro AI și colab. Farmacocinetica trans-resveratrolului după administrarea repetată la subiecți vârstnici și tineri sănătoși. J Clin Pharmacol. 2009; 49 : 1477–82. [ PubMed ] [ Google Scholar ][12] Patel KR, Brown VA, Jones DJ, Britton RG, Hemingway D, Miller AS și colab. Farmacologia clinică a resveratrolului și a metaboliților săi la pacienții cu cancer colorectal. Cancer Res. 2010; 70 : 7392–9. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ][13] Walle T, Hsieh F, DeLegge MH, Oatis JE, Jr., Walle UK. Absorbție ridicată, dar biodisponibilitate foarte scăzută a resveratrolului oral la om. Eliminarea de droguri Metab. 2004; 32 : 1377–82. [ PubMed ] [ Google Scholar ][14] Aggarwal BB, Bhardwaj A, Aggarwal RS, Seeram NP, Shishodia S, Takada Y. Rolul resveratrolului în prevenirea și terapia cancerului: studii preclinice și clinice. Anticancer Res. 2004; 24 : 2783–840. [ PubMed ] [ Google Scholar ][15] Athar M, Back JH, Tang X, Kim KH, Kopelovich L, Bickers DR și colab. Resveratrol: o revizuire a studiilor preclinice pentru prevenirea cancerului uman. Toxicol Appl Pharmacol. 2007; 224 : 274–83. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ][16] Aziz MH, Kumar R, Ahmad N. Chimioprevenția cancerului prin resveratrol: studii in vitro și in vivo și mecanismele de bază (revizuire) Int J Oncol. 2003; 23 : 17–28. [ PubMed ] [ Google Scholar ][17] Bishayee A, Polita T, Darvesh AS. Resveratrol în chimioprevenția și tratamentul carcinomului hepatocelular. Cancer Treat Rev. 36 : 43-53. [ PubMed ] [ Google Scholar ][18] Hintz R, Johnson K. Efectul distribuției mărimii particulelor asupra vitezei de dizolvare și absorbției orale. Jurnalul Internațional de Farmacie. 1989; 51 : 8. [ Google Scholar ][19] Elliott PJ, Jirousek M. Sirtuins: ținte noi pentru bolile metabolice. Curr Opin Investig Droguri. 2008; 9 : 371–8. [ PubMed ] [ Google Scholar ][20] Delmas D, Rebe C, Lacour S, Filomenko R, Athias A, Gambert P, și colab. Apoptoza indusă de resveratrol este asociată cu redistribuirea Fas în plute și formarea unui complex de semnalizare care induce moartea în celulele cancerului de colon. J Biol Chem. 2003; 278 : 41482–90. [ PubMed ] [ Google Scholar ][21] Hope C, Planutis K, Planutiene M, Moyer MP, Johal KS, Woo J, și colab. Concentrațiile scăzute de resveratrol inhibă fluxul semnalului Wnt în celulele derivate din colon: implicații pentru prevenirea cancerului de colon. Mol Nutr Food Res. 2008; 52 (Supliment 1): S52–61. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ][22] Bishayee A, Dhir N. Chimioprevenția mediată de resveratrol a hepatocarcinogenezei inițiate de dietilnitrosamină: inhibarea proliferării celulare și inducerea apoptozei. Chem Biol Interact. 2009; 179 : 131–44. [ PubMed ] [ Google Scholar ][23] Sale S, Verschoyle RD, Boocock D, Jones DJ, Wilsher N, Ruparelia KC și colab. Farmacocinetica la șoareci și proprietățile inhibitoare ale creșterii agentului chemopreventiv al cancerului putativ resveratrol și analogul sintetic trans 3,4,5,4′-tetrametoxistilbene. Br J Rac. 2004; 90 : 736-44. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ][24] Vitrac X, Desmouliere A, Brouillaud B, Krisa S, Deffieux G, Barthe N și colab. Distribuția [14C] -trans-resveratrolului, un polifenol chemopreventiv al cancerului, în țesuturile șoarecilor după administrare orală. Life Science. 2003; 72 : 2219–33. [ PubMed ] [ Google Scholar ]
Mecanismul prin care cancerul mediază atrofia musculară a fost delimitat în ultimele 3 decenii și include un rol proeminent al citokinelor derivate din tumori, cum ar fi IL-6, TNFa și IL-1.Aceste citokine interacționează cu receptorii lor concomitenți pe mușchi pentru a activa factorul de transcripție în aval NF-kB și pentru a induce proteoliza sarcomerului.Experimental, inhibarea semnalizării NF-κB previne în mare parte pierderea musculară indusă de cancer, indicând rolul său proeminent în atrofia musculară.Resveratrolul, un fitoalexin natural găsit în pielea strugurilor, sa dovedit recent că inhibă NF-κB în celulele canceroase, ceea ce ne-a determinat să presupunem că ar putea avea un rol protector în casexia cancerului.Prin urmare, am investigat dacă resveratrolul adminsitrat pe cale orala zilnic ar putea proteja împotriva pierderii mușchilor scheletici și a atrofiei cardiace într-un model de șoarece stabilit.Se demonstrează că resveratrolul inhibă atrofia mușchilor scheletici și cardiacă indusă de tumorile adenocarcinom C26 prin inhibarea activității NF-kB (p65) în mușchiul scheletic și inima .Aceste studii demonstrează pentru prima dată utilitatea terapiei cu resveratrol oral pentru a oferi beneficii clinice în atrofia indusă de cancer prin inhibarea NF-kB în mușchi.Aceste descoperiri pot fi aplicate în tratamentul bolilor cu patofiziologie paralelă, cum ar fi distrofia musculară și insuficiența cardiacă.
Introducere
Casexia de cancer este un sindrom de risipire care afectează până la 80% dintre pacienții cu cancer cu boală avansată 1 – 3 .În timp ce cașexia cancerului este estimată a fi cauza morții la mai mult de 20% din toți pacienții cu cancer, morbiditatea și prăbușirea calității vieții afectează aproape toți pacienții2 , 4 , 5 .Din punct de vedere clinic, manifestările de cașexie a cancerului sunt bine stabilite și includ pierderea masei musculare scheletice și a țesutului adipos, anorexie, astenie și anemie, cu modificări considerabile ale metabolismului lipidic, carbohidrat și proteic2 , 5 , 6 .Pacienții rămân într-o stare hipermetabolică pe măsură ce sarcina tumorală crește, datorită producției tumorale a citokinelor pro-inflamatorii catabolice5
Casexia de cancer este o tulburare metabolică complexă care diminuează masa corporală slabă ca răspuns la creșterea citokinelor inflamatorii eliberate de tumori, incluzând TNFa, IL1 și IL-6 7 , 8 .Aceste citokine derivate din tumori interacționează cu receptorii lor pe mușchi striat pentru a activa factorul de transcripție NF-κB pentru a spori atrofia musculară prin creșterea degradării sarcomerelor 9-12 .Degradarea și proteoliza accelerată a proteinei din aparatul contractil este mediată de sistemul proteazomic ubiquitin, incluzând ligazele ubiquitin specifice în mușchi, cum ar fi inelul muscular deget-1 (MuRF1) 7 ,13 .Inflamația citokinică activată de NF-kB este centrală în atrofia musculară găsită într-o serie de boli, incluzând distrofia musculară, insuficiența cardiacă, sepsisul și cancerul 14-16 .Inhibarea farmacologică și genetică a NF-kB previne pierderea musculară dependentă de ubiquitină10 , 17 și poate conduce la reținerea masei musculare, a forței și poate favoriza regenerarea musculară 18-20 .În consecință, inhibarea activității NF-kB are un potențial enorm pentru prevenirea și tratamentul atrofiei musculare.
Resveratrolul, un fitoalexin natural găsit în pielea strugurilor, s-a dovedit recent că are efecte benefice asupra unei varietăți de procese ale bolii, incluzând carcinomul hepatocelular 21 și boala neurodegenerativă22.Activitatea resveratrolului în aceste boli a fost datorată, în parte, capacității sale de a inhiba translocarea și acumularea nucleară a NF-kB.Aceasta face acest lucru prin inhibarea primară a kinazei IkB (IKK), a cărei activare este necesară pentru activitatea NF-kB.În studiul de față, testăm ipoteza că tratamentul cu resveratrol inhibă cașexia indusă de cancer prin inhibarea NF-kB pentru a proteja împotriva atrofiei cardiace induse de cancer in vivo .
Materiale si metode
Linii de celule
Celulele adenocarcinom C-26 transplantabile s-au menținut așa cum s-a descris anterior 23 , 24
Animale și implantarea tumorilor
96 de șoareci CD2F1 de sex feminin, în vârstă de 45 până la 70 de zile (greutate de 19 până la 22 de grame), au fost obținute de la Charles River Laboratories (Wilmington, MA) și divizate aleatoriu în patru grupe: 1) șoareci martor, 2) șoareci martor tratați cu resveratrol 100 mg / kg, 200 mg / kg sau 500 mg / kg), 3) șoareci purtători de tumori și 4) șoareci purtători de tumori tratați cu resveratrol (100 mg / kg, 200 mg / kg sau 500 mg / kg ).În ziua 0, șoarecii selectați să meargă în grupurile care poartă tumori au fost injectați subcutanat în flancul drept cu 150 pL (aproximativ 750.000 de celule) de celule adenocarcinom C-26.
Tratamentul cu resveratrol
Trans-resveratrolul (Cayman Chemical, Ann Arbor, MI, catalogul # 70675) a fost solubilizat în 1,5% metilceluloză / 0,2% Tween-20 cu agitare viguroasă.Începând din ziua 6, șoarecii au primit tratament oral pe cale orală unică, administrat intragastric, utilizând un ac de 20 g gavage.Șoarecii tratați cu resveratrol au primit tratamentul zilnic cu o singură gavaj de 100 mg / kg, 200 mg / kg sau 500 mg / kg resveratrol, în timp ce martorii au primit un tratament comparabil de 250 μl de vehicul în monoterapie.
Analiza compoziției corpului
Înainte de injectarea tumorală, toate animalele au fost supuse analizei compoziției corporale utilizând ecografice RMN echo-resonanță cantitativă (QMR) (EchoMRI-100, Echo Medical Systems, Houston TX).
Ecocardiografia
Ecocardiografia a fost efectuată pe șoareci conștienți utilizând un sistem de biosimicroscopie cu ultrasunete Visual Sonics Vevo 770, așa cum a fost descris anterior de laboratorul nostru 25 .
Determinarea totală a izolației ARN / determinarea PCR în timp real a expresiei mRNA
ARN-ul total a fost izolat din țesutul înghețat rapid folosind Trizol (Sigma Chemical, St. Louis, MO), cADN realizat utilizând kitul de arhivare cu capacitate mare (Applied Biosystems) de mare capacitate.RT-PCR semi-cantitativ a fost realizat folosind reactivi MJ Research Inc. Thermo Cycler (Waltham, MA) și New England Biolab (Ipswich, MA) folosind primerii detaliați în date suplimentare .
Analiza activității NF-kB
Extractele nucleare au fost izolate din inimă și mușchiul gastrocnemius imediat după recoltare și activitatea NF-kB determinată conform protocolului producătorului (Cayman Chemical, Ann Arbor MI).
Materialele și metodele suplimentare pot fi găsite în secțiunea de date suplimentare .
Rezultate
Resveratrolul nu a evidențiat nici o toxicitate indusă la nivelurile utilizate în acest studiu
Studiile farmacologice care detaliază metabolizarea resveratrolului atât la animale cât și la om au fost efectuate și toate indică absorbția rapidă, distribuția, metabolismul și excreția resveratrolului 26 , 27 .După administrarea orală, distribuția largă a țesuturilor la șobolani a fost observată după 24 de ore și scade semnificativ după 72 de ore.Studiile toxicologice au fost de asemenea completate cu toleranță la doze de până la 700 mg / kg la șobolani 28 fără efecte toxice și la mai multe studii clinice de fază I sunt în curs de administrare la doze orale de până la 100 mg / kg și zi la om26 , 28 .Prin urmare, am administrat resveratrol pe cale orală la 100-500 mg / kg / zi în studiul de față.Am monitorizat toxicitatea prin histologie și letalitate pe tot parcursul studiului.Nu s-au observat evenimente care să cuprindă administrarea de droguri, incluzând orice toxicitate sau letalitate evidentă cauzată de gavajul oral zilnic de resveratrol 100-500 mg / kg și zi.În plus, nu am observat nicio hepatotoxicitate sau nefrotoxicitate indusă de medicamente în toate dozele de administrare a resveratrolului, așa cum a fost confirmată de un patolog independent, orbit, pe examinarea histologică ( Figura 1 ).
Resveratrolul protejează împotriva pierderii de greutate indusă de adenocarcinom C26 la șoareci
Am testat capacitatea resveratrolului de a preveni atrofia indusă de cancerul scheletului și a mușchilor cardiace în modelul soarecilor cu adenocarcinom C26 29 descris anterior de laboratorul nostru 24 .Celulele adenocarcinom C26 cultivate au fost injectate în flancul din spate unde se dezvoltă într-o tumoare și induc pierderea musculară la șoareci CD2F1 în decurs de 2 săptămâni.În studiul prezent, tumorile au fost palpabile în ziua 6 după inoculare la șoarecii de subset care purtau tumori.În ziua 17, la finalizarea studiului, nu au existat diferențe semnificative în greutatea tumorii în rândul tuturor șoarecilor purtători de tumori ( Figura 1A ).purtători de Tumori țesut au avut o pierdere medie în greutate de 8,8 ± 1,7% față de greutățile lor inițiale, în timp ce șoarecii de control au câștigat 2,1 ± 1,8% în perioada experimentală ( Figura 1B ), în ciuda tuturor grupurilor care au un consum echivalent de hrană ( Figura 1C ).
În contrast, șoarecii provocați cu tumori și tratați cu 200 mg / kg de resveratrol au avut greutate similară cu grupul martor.Șoarecii tratați cu 500 mg / kg și provocați cu tumora și-au menținut greutatea în comparație cu șoarecii purtători de tumori ( Figura 1B ).
Șoarecii cu tumori tratați cu 100 mg / kg de resveratrol nu au fost protejați împotriva cașexiei induse de tumori, pierzând 4,6 ± 2,9% din greutatea corporală față de valoarea inițială.
Aceste constatări demonstrează că resveratrolul în doze> / = 200 mg / kg protejează împotriva pierderii de greutate induse de tumori la acest model de șoarece.
Greutatea tumorală finală și consumul mediu de alimente
(A)Nu au existat diferențe semnificative în greutatea tumorilor în grupurile care poartă tumori la toate greutățile tumorale și în subgrupul de tumori mai mari de 0,56 g la momentul recoltării utilizând testele t neparate.( B) S-au observat schimbări semnificative în scăderea în greutate a carcasei la șoarecii purtători de tumori netratate comparativ cu șoarecii martor, șoarecii martor care au primit resveratrol la toate dozele și șoarecii purtători de tumori care au primit resveratrol la dozele de 200 mg / kg și 500 mg / kg.Greutatea carcasei a fost determinată prin scăderea greutății tumorii față de greutatea corporală.Greutatea totală a carcaselor a fost analizată și reprezentată de variația procentuală față de valoarea inițială în rândul tuturor grupurilor.
De asemenea, nu s-au observat diferențe semnificative în consumul mediu de alimente (C) per șoarece pe zi.n / s = nu a fost detectată nici o semnificație ## = p <0,0001, # = p <0,001, ** = p <0,01, * = p <0,05.Pentru (A) N = 24 (Tumor), N = 8 (Tumor + R100), N = 7 (Tumor + R200), N = 8 (Tumor + R500).Pentru (B) și (C) N = 24 (Tumor), N = 8 (Tumor + R100), N = 7 (Tumor + R200), N = 8 (Tumor + R500).
Pentru a delimita dacă resveratrolul a avut vreun efect independent asupra masei musculare, șoarecii de control fără tumori au fost tratați cu 100-500 mg / kg în paralel cu șoarecii tratați cu tumora.Am constatat că șoarecii de control non-tumorali tratați numai cu resveratrol au prezentat greutăți corporale egale cu șoarecii de control la doze mai mici (100 mg / kg, 200 mg / kg).La cea mai mare doză testată (500 mg / kg), șoarecii au pierdut greutatea în comparație cu controalele corespunzătoare vârstei ( Figura 1B ).Aceste rezultate arată că 500 mg / kg de resveratrol poate determina o scădere în greutate în absența cașexiei, în timp ce nivelurile mai scăzute (100-200 mg / kg) nu afectează greutatea corporală.
Niciuna dintre dozele de resveratrol testate nu determină o creștere a masei musculare, sugerând că efectele protectoare ale resveratrolului nu s-au datorat creșterilor independente ale masei musculare.
Resveratrolul protejează împotriva pierderii de carne slabă indusă de cașexie și pierderea masei grase
Casexia de cancer se caracterizează prin pierderea atât a masei musculare scheletice, cât și a țesutului adipos rezultat din modificări ale metabolismului proteinelor și lipidelor.2 , 5 , 6 Următoarele au determinat compoziția corporală a șoarecilor provocați cu tumori în prezența sau absența resveratrolului prin analiza compoziției corpului RMN pentru a determina masa corporală slabă (LBM) și masa de grăsime (FM).Am identificat faptul că resveratrolul de 200 și 500 mg / kg protejează în mod semnificativ împotriva pierderii induse de tumoare a compoziției LBM și FM la șoarecii purtători de tumori ( figura 2A și 2B ).Comparativ cu martorii, șoarecii purtători de tumori netratați au pierdut 5,3% LBM (16,1 ± 0,3 față de 17,1 ± 0,1 g), comparativ cu șoarecii purtători de tumori tratați cu resveratrol, care au fost complet protejați (LBM (200 mg / kg) = 17,4 ± 0,19 g ; LBM (500 mg / kg) = 17,3 ± 0,3 g).Șoarecii purtători de tumori netratați au pierdut 33,3% din FM limitat, comparativ cu șoarecii de control (1,80 ± 0,15 față de 3,1 ± 0,10 g).Tumorile din șobolan tratate cu 200 și 500 mg / kg au fost protejate în mod semnificativ (2,7 ± 0,36 g și 2,4 ± 0,17 g, respectiv).Tratamentul cu resveratrol administrat șoarecilor de control fără tumori nu a afectat LBM sau FM.Deoarece resveratrolul nu a cauzat creșteri ale masei corporale totale (inclusiv LBM și FM) în absența tumorii, aceste studii indică faptul că protecția resveratrolului împotriva pierderii de greutate indusă de tumori nu se datorează capacității sale de a mări greutatea musculară, LBM sau FM .
Compoziția finală a corpului în rândul șoarecilor care nu poartă tumori și care poartă tumori
Întreaga compoziție a corpului de șoarece a fost determinată și prezentată pentru a arăta masa finală în grame în rândul tuturor grupurilor.
(A)S-a observat o pierdere semnificativă a masei musculare slabe la șoarecii purtători de tumoare netratată comparativ cu șoarecii martor, șoarecii martor care au primit resveratrol la toate dozele și șoarecii purtători de tumori care au primit resveratrol la nivelurile de 200 mg / kg și 500 mg / kg.
(B)Compozițiile în masă de grăsimi au fost comparate între aceleași subseturi cu o pierdere semnificativă de masă de grăsime prezentată în grupul care poartă un tumoră netratată.Grupurile care poartă tumori care primesc resveratrol au prezentat răspunsuri semnificative prin menținerea unei cantități mai mari de grăsimi în grupurile de 200 mg / kg și 500 mg / kg.Grupurile de control netratate și tratate au fost, de asemenea, semnificativ mai mari în compoziția masei de grăsime comparativ cu purtătorul de tumoare netratat.p <0,0001, # = p <0,001, ** = p <0,01, p = 0,05.Pentru (A) și (B) N = 24 (Tumor), N = 8 (Tumor + R100), N = 8 (Tumor + R200), N = 8 (Tumor + R500).Pentru (C) N = 17 (Tumor), N = 6 (Tumor + R100), N = 6 (Tumor + R200), N = 7 (Tumor + R500).
Resveratrol inhibă gastrocnemius indus de tumori și atrofie cardiacă
Cvadricepsul izolat și inima au fost colectate și cântărite imediat după recoltare.Tumorile cvadricepsice ale șoarecelui au o greutate semnificativ mai mică (~ 23%) decât mușchiul cvadriceps corespunzător de la șoarecii de control ( Figura 3A ).muchiquadriceps izolat de la șoarecii de control tratați cu resveratrol 100 mg / kg, 200 mg / kg și 500 mg / kg nu a fost semnificativ diferit față de șoarecii martor ( Figura 3A ).Dimpotrivă, mușchii cvadriceps de la șoarecii cu tumoare provocată tratați cu resveratrol 200 mg / kg și 500 mg / kg au fost semnificativ mai mari (16,6 ± 6,5%, respectiv 10,3 ± 4,9%) decât șoarecii cu tumori netratate ( Figura 3A )
doze > = 200 mg / kg de resveratrola protejat împotriva atrofiei mușchilor scheletici indusă de tumori.soarecii purtători de tumori au demonstrat atrofie semnificativă la nivelul membrelor posterioare, așa cum reiese din pierderea de masă de 12,2% comparativ cu martorii care nu poartă tumori ( Figura 3B ).Pentru a determina un efect mai general asupra mușchilor scheletici, am cântărit, de asemenea, masa membrelor din spate.Masa musculară a membrelor din spate a fost păstrată la animalele tratate cu resveratrol (200 și 500 mg / kg), evidențiate prin diferențe semnificative la șoarecii tratați cu tumori având o masă mai mare de 12,4 ± 5,7% și, respectiv, 9,2 ± 4,3% față de cei netratați tumoare provocate de soareci.Tratamentul cu resveratrol la șoarecii de control nu diferă semnificativ de șoarecii de control la toate cele trei doze testate.
Greutăți finale în rândul șoarecilor care nu poartă tumori și care poartă tumori
(A)Quadricepsul a fost analizat individual și s-a arătat o pierdere semnificativă în greutate în grupul purtător al tumorii netratate în comparație cu controlul, controalele tratate la toate dozele, precum și grupurile cu tumori tratate la dozele de 200 și 500 mg / kg.Grupul purtător al tumorilor tratate cu 100 mg / kg nu a fost semnificativ diferit în greutatea cvadricepsului din grupul care purta tumora netratată.
(B)Grupurile purtătoare de tumori care au primit resveratrol au prezentat răspunsuri semnificative prin menținerea greutăților din spate comparabile cu cele de control în grupurile de 200 mg / kg și 500 mg / kg.Grupurile de control netratate și tratate au fost, de asemenea, semnificativ mai mari la greutățile din spate, comparativ cu șoarecii purtători de tumori netratați.S-au comparat greutățile din spate între aceleași subgrupuri cu pierderi semnificative de greutate prezentate în grupul purtător al tumorii netratate.
(C)S-au observat scăderi semnificative ale greutății inimii la șoarecii purtători de tumori netratate comparativ cu șoarecii martor, șoarecii martor care au primit resveratrol la toate dozele și șoarecii purtători de tumori care au primit resveratrol la dozele de 200 mg / kg și 500 mg / kg.Schimbarea greutății inimii printre șoarecii care nu poartă tumori și care poartă tumori.Inimi individuale au fost cântărite și sunt reprezentate grafic aici pentru a arăta greutatea inimii ca procent din greutatea medie a inimii de control în rândul tuturor grupurilor.p <0,0001, # = p <0,001, ** = p <0,01, p = 0,05.Pentru controlul N = 8 (Control + R100), N = 8 (Control + R200), N = 8 (Control + R500) = 6 (Tumor + R100), N = 6 (Tumor + R200), N = 7 (Tumor + R500).
soarecii purtători de tumori au prezentat o pierdere semnificativă a greutății totale a inimii în comparație cu șoarecii de control.Tratamentul cu 200 și 500 mg / kg a fost protector împotriva acestei pierderi de greutate totală a inimii de 8,6% identificată după 17 zile de sarcină tumorală ( Figura 3C ).
Tratamentul șoarecilor de control cu 100 mg / kg, 200 mg / kg, 500 mg / kg de resveratrol nu a afectat greutatea totală a inimii ( Figura 3C ).
Pentru a caracteriza în continuare efectele resveratrolului asupra inimii, am efectuat ecocardiografia trans-toracică in vivo pe șoareci conștienți din control, provocată de tumori cu și fără resveratrol 500 mg / kg ( Figura 4A, 4B , Tabelul 1 ).Am identificat faptul că 17 zile de sarcină tumorală a determinat o scădere a peretelui anterior și posterior al ventriculului stâng, reflectând atrofia cardiacă identificată prin scăderea raportului greutate corporală / greutate corporală și grosimea relativă a peretelui ( Figura 4 ).În primul nostru set de studii, șoarecii care au provocat tumora și care au fost analizați prin ecocardiografie au pierdut aproximativ 8,8% din greutatea corporală după 17 zile de tumoare ( Figura 1B ).Cu toate acestea, în studiile noastre de ecocardiografie, au pierdut doar 6,8% (= 21,8-20,4 / 21,8) (tumora vs. control, Tabelul 1 ).Aceasta se poate datora dimensiunii eșantionului și variației dimensiunii tumorii pe care am văzut-o la șoarecii individuali.Tratamentul șoarecilor provocați de tumori cu resveratrol 500 mg / kg a împiedicat complet scăderea grosimii peretelui anterior și posterior ( Figura 4A, 4B , Tabelul 1 ).Tratamentul șoarecilor de control cu resveratrol nu a afectat grosimea peretelui VS de bază sau greutatea inimii ( Figura 4A , Tabelul 1 ), ceea ce sugerează că protecția resveratrolului adusa inimii nu a fost o creștere a masei musculare indusă de medicament, dar mai probabil datorită unei inhibiții a semnalizării căile care mediate atrofia cardiacă.
Resveratrolul protejează împotriva atrofiei cardiace induse
(A) Ecocardiografia conștientă a ventriculului stâng a identificat scăderi semnificative ale grosimii peretelui anterior și posterior în diastol, greutatea inimii față de greutatea corporală (HW / BW) și grosimea peretelui relativ, care nu au fost identificate când 500 mg / kg / zi s-a administrat resveratrol.(B) Înregistrări reprezentative M-mode din imaginile utilizate în analiza (A).A fost efectuată o analiză ANOVA cu o singură cale pentru a determina semnificația, urmată de o comparație între perechi de Holm-Sidak și semnificație între grupuri.* P <0,05.BW, greutatea corporală, greutatea corporală, greutatea inimii;RWT = LVPW; d / s + LVAntW;d / s / LVEDD sau LVEDS;Valoarea LV (index) = [1,055 * ((ExLVD; d) 3 – (LVEDD; d) 3 ).N = 4 pe grup.
tabelul 1
Ecocardiografia transtoracică la tratamentul neanestetizat pentru tumoare, tumoră sau tumoră plus resveratrol la șoareci maturi la vârsta de 16 zile după implantarea tumorii (± SE)
A fost efectuat un ANOVA cu o singură cale pentru a determina semnificația.
Control N = 8
tumoare N = 4
Tumor + 500R N = 4
Control + 500R N = 4
Greutatea corporală (g)
21,8 ± 0,4
20,4 ± 1,1
21,6 ± 0,8
20,6 ± 0,4
HR (bpm)
651 ± 9
617 ± 27
640 ± 13
659 ± 13
LVEDD (mm)
2,63 ± 0,09
2,95 ± 0,08 *
2,82 ± 0,13
2,50 ± 0,07
LVESD (mm)
1,11 ± 0,05
1,35 ± 0,07
1,19 ± 0,10
1,08 ± 0,06
LV% EF
88,97 ± 0,46
85,78 ± 2,78
88,64 ± 1,86
88,20 ± 0,94
LV Vol;d (ml)
25,9 ± 2,2
33,8 ± 2,2
30,6 ± 3,5
22,8 ± 1,5
LV Vol;s (ml)
2,9 ± 0,3
4,7 ± 0,7
3,5 ± 0,8
1,7 ± 1,5
BW, greutate corporală;LV, ventriculul stâng;HR, ritm cardiac;LVEDD, dimensiunea diastolică la capătul LV;LVESD, dimensiunea sistolică a capătului LV;LV EF% = [(LV Vol; d-LV Vol; s / LV Vol; d) * 100];d, volumul LV în diastol;Volumul LV Vol; s, Volumul LV în sistol;LV Post W.
* p <0,05 față de Control, Tumor + 500R și Control + 500R.
Resveratrolul inhibă activitatea factorului de transcripție NF-κB în mușchii scheletici indusă de tumori și atrofia inimii
Tumora de adenocarcinom C-26 induce cașexia prin eliberarea de TNFa, IL-1, IL-6, care interacționează cu receptorii lor concomitenți pe mușchi pentru a activa factorul de transcripție în aval NF-κB30-34, pentru a induce cașexia și atrofia musculară.Studii recente au demonstrat experimental rolul dominant al NF-kB în atrofia musculară 18-20 , 35 , 36 și capacitatea resveratrolului de a inhiba NF-κB în celulele canceroase 37.Prin urmare, am investigat dacă inhibarea resveratrolului de atrofie indusă de tumori și atrofia inimii s-a datorat în parte inhibării sale de NF-kB ( Figura 5 ).Am identificat activitatea NF-κB (subunitatea p65) legată de activitatea nucleară semnificativ crescută în gastrocnemiusul și inima indusă de tumoare și complet inversată la șoarecii tratați cu cele mai ridicate niveluri de resveratrol (200 mg / kg și 500 mg / kg) (Figura 5A , 5B ).Gastrocnemiusul muscular de la șoareci purtători de tumori netratate a avut o activitate medie NF-kB mai mare cu 30,5% comparativ cu șoarecii martor ( Figura 5A ).S-a observat o scădere semnificativă a activității NF-kB (p65) la șoarecii purtători de tumori, cărora li s-au administrat resveratrol 200 și 500 mg / kg (scăzut cu 39% și, respectiv, 51%) ( Figura 5A ).Șoarecii de control tratați au avut, de asemenea, o scădere semnificativă a activității NF-kB (p65) după tratamentul cu doze de 200 și 500 mg / kg (scăzând 34% și, respectiv, 41%) comparativ cu șoarecii fără tumori provocați de tumoare, indicând că resveratrolul a scăzut NF endogen -kB indusă de tumoare în mușchii scheletici.
(A) Extracțiile nucleare pe mușchiul gastrocnemius au fost analizate în vederea activității ADN-ului legat de p65.S-a observat o creștere semnificativă a activității la șoarecii purtători de tumori netratați în comparație cu martorul.Tumorile purtătoare de tumori tratate cu resveratrol au prezentat o scădere semnificativă a activității NF-kB la doze de 200 mg / kg și 500 mg / kg.De asemenea, în grupul tratat fără tumori tratat, activitatea NF-kB a fost semnificativ scăzută și comparabilă cu controlul cu administrarea de resveratrol la dozele de 200 mg / kg și 500 mg / kg.
(B)Grupurile purtătoare de tumori care au primit 200 mg / kg și 500 mg / kg resveratrol au prezentat o inhibare semnificativă a activității NF-kB indusă de tumoare.
(C) Șoarecii purtători de tumori care au primit 200 mg / kg și 500 mg / kg resveratrol au avut o inhibare a expresiei MuRF1 indusă de tumoare.expresia mRNA a fost măsurată și descrisă aici ca normalizată la controalele GAPDH.# = p <0,001, ** = p <0,01, * = p <0,05.Pentru (A) și (B): N = 12 (Control);N = 4 (Tumor + R200), N = 4 (Control + R100), N = 4 (Control + R200) ).Pentru (C): N = 4 (Control), N = 3 (Control + R100), N = 3 (Control + R200) Tumor + R100), N = 3 (Control + R200), N = 3 (Control + R500).
Analiza inimii a arătat că provocarea tumorală a indus o creștere de 19% a activității NF-kB comparativ cu inimile de control ( Figura 5B ).Tratamentul șoarecilor purtători de tumori cu resveratrol 200 și 500 mg / kg a redus semnificativ activitatea NF-kB (p65) comparativ cu șoarecii cu tumoare provocată (21,6% și, respectiv, 31,2%) ( Figura 5B ).Tratamentul cu resveratrol 200 și 500 mg / kg a determinat o reducere semnificativă a activității NF-kB (p65) la șoarecii martor (18,7% și, respectiv, 33,8%) comparativ cu șoarecii fără tumori provocați de tumori.Studii recente au demonstrat că inducerea NF-kB a ligazelor ubiquitin E3 reglează atrofia musculară scheletică 10 , 11 .Mai mult, laboratorul nostru a determinat rolul critic al atrofiei cardiace ubiquitin ligase finger-1 (MuRF1) în atrofia cardiacă 25 .Mai exact, am identificat faptul că șoarecii care nu au prezentat MuRF1 nu au putut, în esență, să se atrofie ca răspuns la dexametazonă 25 .Aceasta contrastează cu mușchiul scheletic, unde atât MuRF1, cât și Atrogin-1 joacă un rol parțial în inhibarea atrofiei musculare scheletice 13 .Am urmărit în continuare investigația noastră asupra modului în care inhibarea resveratrolului de NF-kB poate afecta expresia MuRF1 cardiacă in vivo.
Resveratrol inhibă expresia mRNA MuRF1 cardiacă
Afectarea tumorală a indus expresia mRNA MuRF1 în inimă în mod semnificativ (~ 20%) după 17 zile de tumoare ( Figura 5C ).În timp ce tratamentul cu resveratrol nu a afectat expresia MuRF1 la animalele de control, rezveratrolul de 200 și 500 mg / kg a inhibat semnificativ MuRF1 indus de tumoare comparativ cu provocarea tumorii în monoterapie (~ 25% și, respectiv, 75%), atingând niveluri de mRNA ale MuRF1 semnificativ mai mici care au fost găsite la animalele de control ( figura 5C ).Deoarece MuRF1 joacă un rol esențial în atrofia cardiacă in vivo 25 și MuRF1 este reglat în parte de către NF-κB 10,11 , aceste constatări sugerează un mecanism prin care resveratrolul inhibă atrofia cardiacă indusă de tumori prin inhibarea indusă de NF-κB Expresia MuRF1 in vivo.
Discuţie
Modelul C26-adenocarcinom al cașexiei de cancer a fost utilizat în ultimele 3 decenii pentru cercetarea istoricului natural al carcinoamelor și a terapiei antitumorale, așa cum a fost recent revizuită de Aulino și colab., 2010 29 .Aproximativ 190 de lucrări pot fi găsite prin căutarea în PubMed pentru cancerul C26, multe dintre ele concentrându-se pe Cachexia 29 indusă de C26.Modelul C26-adenocarcinom a fost stabilit în 1975 într-un efort de a crea modele care ar putea fi utilizate pentru a testa biologic și chemo-terapii38.Acesta poate fi stabilit la șoareci nud și SCID, precum și șoareci de tip sălbatic, sugerând că limfocitele joacă un rol mic, dacă este cazul, în fiziopatologia progresiei tumorale 31 .Mecanismul predominant prin care linia celulară de adenocarcinom C26 induce cașexia este prin eliberarea citokinelor inflamatorii 31 , paralel cu mecanismul prin care cancerul provoacă cașexie la om8 .
Studiile inițiale ale modelului de șoarece cu C26-adenocarcinom, așa cum a fost utilizat în studiul prezent, au identificat un rol proeminent al IL-6 în inducerea atrofiei musculare.În aceste studii, inhibarea IL-6 utilizând reactivi specifici citokinelor a prevenit atrofia musculară indusă la șoarecii purtători de adenocarcinom C26.Studiile suplimentare au arătat că IL-6 nu a fost singurul mecanism pentru inducerea cașexiei musculare 34 , 39 .De asemenea, s-a descoperit că celulele de adenocarcinom C26 secretă TNFa, IL-1 și IL-6 in vivo, împreună cu factorii de cașexie identificați care induc atrofia mușchilor scheletici in vivo 31 , 33 .Aceste citokine proinflamatorii se leagă de receptorii lor congenitali găsiți pe mușchiul și inima scheletului, pentru a activa factorul de transcripție NF-κB40, care mediază efectele lor din aval.Activarea NF-kB este esențială pentru procesul de atrofie a mușchiului striat.Acest lucru a fost dovedit convingător prin utilizarea complecșilor NF-kB inhibitori negativi dominanți (IKK), care inhibă în mod eficient NF-kB.Supraexpresia unei proteine de fuziune IKKα / β-EGFP negativă dominantă, care inhibă atrofia indusă de NF-κB în mod semnificativ inhibată până la 70% in vivo 17 .Alte studii au identificat, de asemenea, că inhibarea farmacologică sau genetică a IKK / NF-κB inhibă în mod asemănător pierderea musculară dependentă de ubiquitină (proteoliza) 10 .În aceste studii, inhibarea NF-kB a inhibat expresia ligandului ubiquitin MuRF1, una dintre cele două ligase ubiquitin care sunt necesare pentru apariția atrofiei mușchilor scheletici10.
Mediatorii mediator de inflamare, cum ar fi TNFa, sunt inductori puternici ai liganței ubiquitin MuRF1 13 .Studiile recente au arătat că expresia MuRF1 este reglată de factorul de transcripție NF-kB10 , 41 , care este activat de un număr de citokine proinflamatorii, incluzând TNFa.În mod similar, tumora de adenocarcinom C26 la șoareci induce catabolismul proteinei prin inducerea ligandului ubiquitin MuRF1 42.Încărcarea C26 induce o pierdere specifică a componentelor sarcomerice de filament groase, dar nu subțiri 24 și modifică expresia izoformei 43 a miozinei, în concordanță cu acțiunile lui MuRF1 44 .Complexul de distrofină este scăzut la șoarecii cu azotocarcinom C26, un fenomen esențial pentru degradarea musculară 42 .Acest model a permis demonstrarea rolului celulelor stem în mușchii scheletici în timpul cașexiei 45 , 46.
În timp ce MuRF1 joacă doar un rol parțial în medierea atrofiei mușchilor scheletici in vivo 13 , laboratorul nostru a identificat recent că activitatea MuRF1 este necesară pentru a induce orice atrofie cardiacă 25 . În mod specific, am identificat faptul că atrofia semnificativă indusă de dexametazonă indusă în inimile de tip sălbatic este în esență absentă la șoareci MuRF1 – / 25 . Prin urmare, am concentrat studiile noastre actuale asupra expresiei MuRF1 a ligazei ubiquitin în inimă. Am presupus că inhibarea resveratrolului de NF-κB cardiac ar reduce expresia MuRF1 cardiacă, reprezentând un mecanism prin care atrofia cardiacă este inhibată în modelul de șoarece de cașexie indus de C26-adenocarcinom. De fapt, am identificat faptul că resveratrolul a inhibat expresia MuRF1 crescută în tumoră in vivo (Figura 5C ). Prin urmare, concluziile noastre constau în prevenirea consecventă a resveratrolului de atrofie cardiacă datorată, în parte, inhibării MuRF1, posibil prin inhibarea lui NF-kB.toate acestea, MuRF1 este reglementatde alți factoritranscripție cum ar fi transcrierea FOXO factori de 47 – 49 , deci este posibil ca resveratrolul inhiba MuRF1 prin alte cai decât NF-kB mecanisme în inimă. Constatarile din prezentul studiu demonstreaza ca resveratrolul protejeaza impotriva atrofiei cardiace induse de inima prin inhibarea NF-kB si MuRF1, sugerand unul sau mai multe mecanisme prin care este cardioprotectiva.
Casexia la pacienții cu cancer uman prezintă pierderi de mușchi, adesea cu pierderea de grăsime asociată cu anorexia, inflamația și rezistența la insulină 50 . Studiile anterioare au demonstrat că cașexia indusă de C26-adenocarcinom induce o pierdere atât a mușchiului muscular, cât și a grăsimii, așa cum am identificat în studiul de față 51 , 52 . Detaliile privind modul în care cașexia determină pierderea de grăsime nu a fost complet delimitată, dar creșterea expresiei lipazei sensibile la hormoni este un efect descendent al factorilor secretați de tumori 50 . Deși este posibil ca citokinele inflamatorii și NF-κB să poată mediaze acest lucru în mușchi, nu este suficient să se ajungă la aceste căi complexe de semnalizare în adipocite în prezent pentru a susține această ipoteză 53 .
Unul dintre punctele forte ale studiului actual este că resveratrolul nu a afectat dimensiunea tumorilor adenocarcinom C26, permițându-ne să determinăm eficacitatea resveratrolului în special la nivelul mușchilor și inimii scheletice. Cu toate acestea, este important de remarcat faptul că resveratrolul a fost raportat pentru a inhiba creșterea tumorii, incluzând mai multe celule de mielom (linie de celule IM-9), celule HeLa (carcinom de col uterin) și celule K-562 (leucemie mieloidă cronică) 37 , 54 , 55 . De fapt, eficacitatea inhibării NF-kB de către resveratrol pentru a proteja împotriva pierderii mușchilor scheletici indusă de tumoarea MAC16 (utilizând 1 mg / kg / zi) într-un studiu anterior efectuat de Wyke și colab. 56ar putea fi atribuită inhibării sale de creștere tumorală în sine, făcând studiul prezent unic din punct de vedere mecanicist. Abilitatea resveratrolului de a inhiba creșterea tumorală în sine poate fi foarte benefică pentru pacienții cu cașexie. Împreună cu studiul nostru prezent, aceste constatări indică faptul că resveratrolul protejează împotriva cașexiei cauzate de cancer prin două mecanisme distincte: 1) prin inhibarea directă a creșterii tumorale, determinând moartea celulelor rezultând într-o reducere a eliberării totale a citokinei; și 2) prin inhibarea semnalizării NF-kB determinată de citokine la nivelul mușchiului și inimii scheletice. Deoarece resveratrolul nu afectează tumora de adenocarcinom C26 în studiul prezent, resveratrolul poate fi chiar mai puternic decât rezultatele studiului prezent demonstrează.
Contribuția complicațiilor cardiace la cachexia cancerului la om nu a fost studiată pe scară largă. Cu toate acestea, mulți dintre mediatorii inflamatorii au descoperit cafexia se găsesc în insuficiența cardiacă și cauzează direct disfuncție cardiacă, incluzând IL-6, IL-1 și TNFa. Cel mai bine studiat este TNFα, care poate provoca insuficiență cardiacă și a fost implicată în patofiziologia unui număr de boli cardiace 7 . Acest lucru sugerează că implicarea cardiacă poate fi frecventă în cazexia cardiacă, în special în cazul unei boli severe. În mod experimental, a fost demonstrat ca cașexia induce insuficiența cardiacă, perturba structura miocardică și modifică compoziția proteinelor contractile la modelele de șoarece 43 , 57. Nu am identificat disfuncția sistolică brută prin ecocardiografie în studiul de față. Cu toate acestea, tumorile mai mari care se formează pe perioade mai lungi pot să fi condus la disfuncție cardiacă în modelul nostru.
În studiul de față, demonstrăm pentru prima dată că resveratrolul inhibă direct atrofia mușchilor scheletici și cardiacă la doze orale de 200-500 mg / kg / zi într-un model de șoarece de cașexie indusă de tumori și de atrofie cardiacă fără nici o toxicitate aparentă lasoarece Un studiu recent realizat de Busquets, et al. a constatat că resveratrolul nu a împiedicat pierderea de mușchi scheletici indusă de cancer, utilizând un model de șobolan cu hepatom de ascites Yoshida AH-130 (tratat cu resveratrol 1 mg / kg / zi) sau cu șoareci purtători ai carcinomului pulmonar Lewis (la 5 și 25 mg / kg / zi resveratrol intraperitoneal) 58 . De asemenea, sobolanilor li sa administrat resveratrol oral (3 mg / kg / zi) în combinație cu ulei de pește intragastric 58. Rețineți că în cadrul modelului intragastric s-au utilizat doze semnificativ mai mici comparativ cu studiul nostru prezent (100-500 mg / kg și zi). Incapacitatea resveratrolului de a proteja împotriva pierderii masei musculare scheletice sau a pierderii de greutate corporală în rozătoarele purtătoare de tumori poate fi, prin urmare, datorată dozelor reduse de resveratrol utilizate în acest studiu anterior.În concordanță cu această ipoteză, utilizarea noastră de 100 mg / kg și zi prin gavaj oral nu a avut niciun efect asupra pierderii musculare, în timp ce o doză de cel puțin 200 mg / kg / zi a fost necesară pentru a proteja din nou cazhexia indusă de tumori. Doza efectivă în studiul actual de șoarece este de aproximativ 3 ori mai mare decât doza testată la om fără toxicitate. În studiul clinic de fază I, o singură doză de 5 g / zi a fost administrată persoanelor fără toxicitate evidentă. 26La o persoană de 70 kg, această doză ar fi de aproximativ 71 mg / kg / zi (5000 mg / 70 kg) sau aproximativ 2,8 ori mai mică decât doza efectivă minimă găsită în studiile actuale (200 mg / kg / zi vs. 71 mg / kg / zi).
Efectele expunerii pe termen lung a resveratrolului în contextul tratării cașexiei cancerului în modelul prezent nu sunt cunoscute. Totuși, trebuie subliniat faptul că șoarecii care au primit 500 mg / kg / zi ( Figura 1B ) au pierdut greutatea în prezența sau absența tumorii, în timp ce șoarecii care au primit doze mai mici nu au făcut-o. Deoarece aceste mici, dar semnificative modificări au fost observate când s-au administrat 500 mg / kg / zi într-o perioadă scurtă de timp (11 zile), studiile viitoare ar trebui să fie atenți la reacțiile adverse potențiale cu terapie cu resveratrol pe cale orală (adică pierderea în greutate) la doze de 500 mg / kg / zi. Acest lucru ar fi deosebit de important în studiile pe termen lung care durează mai mult de 11 zile, ceea ce este probabil modul în care acest medicament va fi utilizat pentru a trata cachexia la pacienți.
Terapiile actuale pentru cașexie includ terapia modală unică sau combinațiile de agenți progestative, glucocorticoizi, inhibitori selectivi ai COX-2 sau medicamente antiinflamatoare nesteroidiene (AINS) cum ar fi aspirina sau indometacinul. Au fost dezvoltate noi tratamente pentru a inhiba într-o anumită măsură citokinele sau NF-κB. Acestea includ progestine cu doză mare 59 , acid eicosapentaenoic (EPA) 60 , beta-hidroxi-β-metilbutirat (HMB) 61 , talidomidă 62 și AINS 63 . Cu toate acestea, acești agenți au efecte secundare adverse și rezultatele au fost variabile și marginale în inversarea cașexiei clinic. Resveratrolul este utilizat în prezent într-o varietate de boli umane, incluzând carcinomul hepatocelular 21și boala neurodegenerativă 22 fără toxicitate marcată 7 . Prin urmare, poate avea o aplicare în cazexia (inclusiv mușchiul scheletic și atrofia cardiacă) și, în general, la insuficiența cardiacă într-o varietate de contexte ale bolii umane, deși acest lucru nu a fost testat direct în prezent.
Examinarea histologică a țesuturilor hepatice și a rinichilor a fost efectuată la mărirea de 20X pentru a căuta semne de toxicitate la medicament. (A) La momentul sacrificării, au fost recoltate ficat și probele histologice preparate în mod standard așa cum este descris în secțiunea metode. Parenchimul hepatic normal a fost observat cu citoplasma uniformă între toate grupurile așa cum se vede în proba de control. (B) Nu s-a detectat nici o toxicitate medicamentoasă în ficat din orice grupuri tratate cu resveratrol, așa cum s-a observat din histologia mouse-ului tratat cu resveratrol cu tumora de 500 mg / kg. (C, D) Nu s-a detectat nici o dovadă de pierdere de hepatocite, inflamație sau steatoză în nici un grup la orice doză. Analiza histologică a rinichilor a evidențiat glomeruli, tubuli și interstițiu normali, fără nici o dovadă de necroză tubulară acută, glomeruloscleroză,sau inflamația interstițială, așa cum s-a demonstrat la un șoarece de control (C), precum și la un șoarece purtătoare de tumori (D) tratate cu resveratrol de 500 mg / kg. N = 4 șoareci pe grup au fost analizate histologic (Control, Control + 100R, Control + 200R, Control + 500R, Tumor, Tumor + 100R, Tumor + 200R, Tumor + 500R). Un șoarece reprezentativ din control și Tumor + 500R este prezentat în figură.
Toate lucrările au fost efectuate la Universitatea Carolina de Nord din Chapel Hill, SUA
Acordarea sprijinului acordat
SS a fost susținută de Universitatea din Carolina de Nord, Departamentul de ORL / Chirurgia capului și a gâtului. Aceasta lucrare a fost sustinuta de catre Universitatea din Carolina de Nord Programul de Grant Stiinta Transferului (la MC), American Heart Association Development Scientist Grant (la MW), precum si Institutul National Heart, Lung si Blood acorda R01HL104129 (la MW).
1. Tisdale MJ. Cachexia la pacienții cu cancer. Nat Rev Cancer.2002;2 : 862-871.[ PubMed ]
2. Fearon KC, Moses AG. Cachexia de cancer. Int J Cardiol.2002;85 : 73-81.[ PubMed ]
3. Inagaki J, Rodriguez V, Bodey GP. Procedura: cauze de deces la pacienții cu cancer.Cancer.1974;33 : 568-573.[ PubMed ]
4. Warren S. Cauzele imediate ale decesului în cancer. Am J Med Sci. 1935; 184 : 610-616.
5. Argiles JM, Busquets S, Lopez-Soriano FJ. Rolul central al citokinelor în pierderea musculară în timpul cancerului. Int J Biochem Cell Biol.2005;37 : 2036-2046.[ PubMed ]
6. Costelli P, Baccino FM. Cachexia de cancer: de la modele experimentale la managementul pacientului. Curr Opin Clin Nutr Metab Îngrijire.2000;3 : 177-181.[ PubMed ]
7. Pajak B, Orzechowska S, Pijet B, Pijet M, Pogorzelska A, Gajkowska B, și colab. Crossroads de semnalizare a citokinelor – urmărirea pentru stoparea cașexiei musculare.J. Physiol Pharmacol.2008;59(Suppl 9): 251-264.[ PubMed ]
8. Carson JA, Baltgalvis KA. Interleukina 6 ca regulator cheie al masei musculare în timpul cașexiei. Exerc Sport Sci Rev. 2010; 38 : 168-176.[ Articol gratuit PMC ][ PubMed ]
9. Tisdale MJ. Mecanismele cașexiei de cancer. Physiol Rev. 2009; 89 : 381-410.[ PubMed ]
10. Cai D, Frantz JD, Tawa NE, Jr, Melendez PA, Oh BC, Lidov HG și colab. Activarea IKKbeta / NF-kappaB cauzează pierdere musculară severă la șoareci.Cell.2004;119 : 285-298.[ PubMed ]
11. Wyke SM, Tisdale MJ. NF-kappaB mediază degradarea proteinelor indusă de proteoliză și expresia sistemului ubiquitin-proteazom în mușchiul scheletic. Br J Cancer.2005;92 : 711-721.[ Articol gratuit PMC ][ PubMed ]
12. Drott C, Ekman L, Holm S., Waldenstrom A, Lundholm K. Efectele încărcării tumorale și malnutriției asupra funcției miocardice în inima izolată de șobolan de lucru. J Mol Cell Cardiol.1986;18 : 1165-1176.[ PubMed ]
13. Bodine SC, Latres E, Baumhueter S, Lai VK, Nunez L, Clarke BA, și colab. Identificarea ligazelor ubiquitin necesare pentru atrofia mușchilor scheletici.Ştiinţă.2001; 294 : 1704-1708.[ PubMed ]
14. Li H, Malhotra S, Kumar A. Semnalarea factorului nuclear-kappa B în atrofia mușchilor scheletici. J Mol Med.2008;86 : 1113-1126.[ Articol gratuit PMC ][ PubMed ]
15. Tisdale MJ. Există un mecanism comun care leagă pierderea musculară în diferite tipuri de boli? Curr Opin Suport Palliat Îngrijire.2007;1 : 287-292.[ PubMed ]
16. Tisdale MJ. Sunt factorii tumorali responsabili pentru risipirea țesutului gazdă în cazexia de cancer? Viitorul Oncol.2010;6 : 503-513.[ PubMed ]
17. Van Gammeren D, Damrauer JS, Jackman RW, Kandarian SC. IkappaB kinazele IKKalpha și IKKbeta sunt necesare și suficiente pentru atrofia mușchilor scheletici. FASEB J. 2009; 23 : 362-370.[ Articol gratuit PMC ][ PubMed ]
18. Eley HL, Russell ST, Tisdale MJ. Atenuarea atrofiei musculare într-un model murin al cașexiei prin inhibarea protein kinazei dependente de dsARN. Br J Cancer.2007;96 : 1216-1222.[ Articol gratuit PMC ][ PubMed ]
19. Acharyya S, Villalta SA, Bakkar N, Bupha-Intr T, Janssen PM, Carathers M, și colab. Interplay-ul semnalizării IKK / NF-kappaB în macrofage și myofibre promovează degenerarea musculară în distrofia musculară Duchenne.J Clin Invest.2007;117 : 889-901.[ Articol gratuit PMC ][ PubMed ]
20. Mourkioti F, Kratsios P, Luedde T, Song YH, Delafontaine P, Adami R și colab. Ablația vizată a IKK2 îmbunătățește forța musculară scheletică, menține masa și promovează regenerarea.J Clin Invest.2006;116 : 2945-2954.[ Articol gratuit PMC ][ PubMed ]
21. Bishayee A, Politis T, Darvesh AS. Resveratrol în chimioprevenție și tratamentul carcinomului hepatocelular. Cancer Treat Rev. 2010; 36 : 43-53.[ PubMed ]
22. Pallas M, Casadesus G, Smith MA, Coto-Montes A, Pelegri C, Vilaplana J, și colab. Resveratrol și bolile neurodegenerative: activarea SIRT1 ca cale potențială către neuroprotecție. Curr Neurovasc Res.2009;6 : 70-81.[ PubMed ]
23. O’Connell TM, Ardeshirpur F, Asher SA, Winnike JH, Yin X, George J, și colab. Analiza metabolică a cașexiei de cancer relevă modificări distincte ale lipidelor și glucozei. Metabolomica.2008;4 : 216-225.
24. Acharyya S, Ladner KJ, Nelsen LL, Damrauer J, Reiser PJ, Swoap S, și colab. Cachexia de cancer este reglementată prin direcționarea selectivă a produselor genetice ale mușchilor scheletici.J Clin Invest.2004;114 : 370-378.[ Articol gratuit PMC ][ PubMed ]
25. Willis MS, Rojas M, Li L, Selzman CH, Tang RH, Stansfield WE, și colab. Degetul inelar muscular 1 mediază atrofia cardiacă in vivo. Am J Physiol Heart Circ Physiol.2009;296 : H997-H1006.[ Articol gratuit PMC ][ PubMed ]
26. Boocock DJ, Faust GE, Patel KR, Schinas AM, Brown VA, Ducharme MP și colab. Studiu farmacocinetic de escaladare a dozei de fază I la voluntari sănătoși de resveratrol, un potențial agent chimiopreventiv al cancerului.Cancer Epidemiol Biomarkers Prev.2007;16 : 1246-1252.[ PubMed ]
27. Walle T, Hsieh F, DeLegge MH, Oatis JE, Jr, Walle UK. Absorbție ridicată, dar cu o biodisponibilitate foarte redusă a resveratrolului oral la om.Drug Metab Dispos.2004;32 : 1377-1382.[ PubMed ]
28. Williams LD, Burdock GA, Edwards JA, Beck M, Bausch J. Studii de siguranță efectuate pe trans-resveratrol de înaltă puritate pe animale experimentale. Food Chem Toxicol.2009;47 : 2170-2182.[ PubMed ]
29. Aulino P, Berardi E, Cardillo VM, Rizzuto E, Perniconi B, Ramina C, și colab. Caracterizarea moleculară, celulară și fiziologică a carcinomului de colon C26 care induce cașexia în cancer la șoarece. BMC Cancer.2010;10 : 363.[ Articol gratuit PMC ][ PubMed ]
30. Sturlan S, Beinhauer BG, Oberhuber G, Huang L, Aasen AO, Rogy MA. Transferul genelor in vivo al interleukinei-4 murine inhibă cașexia cancerului mediată de colonul 26 la șoareci. Anticancer Res.2002;22 : 2547-2554.[ PubMed ]
31. Yasumoto K, Mukaida N, Harada A, Kuno K, Akiyama M, Nakashima E, și colab. Analiza moleculară a rețelei de citokine implicată în cașexia la șobolanii cu șoareci adenocarcinom de colon 26.Cancer Res.1995;55 : 921-927.[ PubMed ]
32. Zhou W, Jiang ZW, Jiang J, Li N, Li JS. [Rolul NF-kappa B în cazexia de cancer] Zhonghua Wai Ke Za Zhi.2004;42 : 683-686.[ PubMed ]
33. Zhou W, Jiang ZW, Tian J, Jiang J, Li N, Li JS. Rolul de NF-kappaB și citokină în cazexia de cancer experimental. World J Gastroenterol.2003;9 : 1567-1570.[ Articol gratuit PMC ][ PubMed ]
34. Soda K, Kawakami M, Kashii A, Miyata M. Manifestările de cașexie de cancer induse de adenocarcinomul de colon 26 nu sunt pe deplin atribuite interleukinei-6.Int J Cancer.1995;62 : 332-336.[ PubMed ]
35. Smith HJ, Wyke SM, Tisdale MJ. Mecanismul atenuării factorului de inducere a proteolizei a determinat degradarea proteinelor în mușchi de beta-hidroxi-beta-metilbutirat.Cancer Res.2004;64 : 8731-8735.[ PubMed ]
37. Holmes-McNary M, Baldwin AS, Jr Proprietățile chemopreventive ale trans-resveratrolului sunt asociate cu inhibarea activării kinazei IkappaB.Cancer Res.2000;60 : 3477-3483.[ PubMed ]
38. Corbett TH, Griswold DP, Jr, Roberts BJ, Peckham JC, Schabel FM., Jr Relații de inducție tumorală în dezvoltarea cancerelor transplantabile ale colonului la șoareci pentru teste de chimioterapie, cu o notă privind structura cancerigenă.Cancer Res.1975;35 : 2434-2439.[ PubMed ]
39. Soda K, Kawakami M, Kashii A, Miyata M. Caracterizarea șoarecilor care poartă subclonele adenocarcinomului de colon 26 descalifică interleukina-6 ca unic inductor al cașexiei. Jpn J Cancer Res.1994;85 : 1124-1130.[ Articol gratuit PMC ][ PubMed ]
40. Vallabhapurapu S, Karin M. Reglarea și funcția factorilor de transcripție NF-kappaB în sistemul imunitar. Annu Rev Immunol.2009;27 : 693-733.[ PubMed ]
41. DJ de sticlă. Hipertrofia mușchilor scheletici și căile de semnalizare a atrofiei. Int J Biochem Cell Biol.2005;37 : 1974-1984.[ PubMed ]
42. Acharyya S, Butchbach ME, Sahenk Z, Wang H, Saji M, Carathers M, și colab. Dysfuncția complexă a glicoproteinei dystrophin: o legătură normativă între distrofia musculară și carachexia cancerului.Cancer Cell.2005;8 : 421-432.[ PubMed ]
43. Diffee GM, Kalfas K, Al-Majid S, McCarthy DO. Expresia alterată a izoformelor miozinei musculare scheletice în cazexia de cancer. Am J Physiol Cell Physiol.2002;283 : C1376-C1382.[ PubMed ]
44. Cohen S, Brault JJ, Gygi SP, Glass DJ, Valenzuela DM, Gartner C, și colab. În timpul atrofiei musculare, componentele filamentului groase, dar nu subțiri sunt degradate prin ubiquitație dependentă de MuRF1. J Cell Biol.2009;185 : 1083-1095.[ Articol gratuit PMC ][ PubMed ]
45. Schwarzkopf M, Coletti D, Sassoon D, Marazzi G. Cachexia musculară este reglementată de o cale dependentă de p53-PW1 / Peg3.Genele Dev.2006;20 : 3440-3452.[ Articol gratuit PMC ][ PubMed ]
46. Berardi E, Aulino P, Murfuni I, Toschi A, Padula F, Scicchitano BM, și colab. Muschiul scheletic este îmbogățit în celule stem hematopoietice și nu celule inflamatorii la șoareci cachectic.Neurol Res.2008;30: 160-169.[ PubMed ]
47. Waddell DS, Baehr LM, van den Brandt J, Johnsen SA, Reichardt HM, Furlow JD, și colab. Receptorul glucocorticoid și FOXO1 activează sinergic gena muRF1 asociată atrofiei mușchilor scheletici. Am J Physiol Endocrinol Metab.2008;295 : E785-E797.[ Articol gratuit PMC ][ PubMed ]
48. Stitt TN, Drujan D, Clarke BA, Panaro F, Timofeyva Y, Kline WO, și colab. Calea IGF-1 / PI3K / Akt previne exprimarea liganzilor ubiquitin indusă de atrofie musculară prin inhibarea factorilor de transcripție FOXO. Mol Cell.2004;14 : 395-403.[ PubMed ]
49. Sacheck JM, Ohtsuka A, McLary SC, Goldberg AL. IGF-I stimulează creșterea musculară prin suprimarea distrugerii proteinelor și a expresiei liganzilor de ubiquitină legați de atrofie, atrogin-1 și MuRF1. Am J Physiol Endocrinol Metab.2004;287 : E591-E601.[ PubMed ]
50. Tisdale MJ. Cachexia de cancer. Curr Opin Gastroenterol.2010;26 : 146-151.[ PubMed ]
51. Weyermann P, Dallmann R, Magyar J, Anklin C, Hufschmid M, Dubach-Powell J, și colab. Disponibil pe cale orală antagoniști selectivi ai receptorilor de melanocortină-4 stimulează administrarea de alimente și reduce cașexia indusă de cancer la șoareci.Plus unu.2009;4 : e4774.[ Articol gratuit PMC ][ PubMed ]
52. Zhou X, Wang JL, Lu J, Song Y, Kwak KS, Jiao Q, și colab. Inversarea canhexiei și a pierderii musculare de către antagoniștii ActRIIB duce la supraviețuirea prelungită.Cell.2010;142 : 531-543.[ PubMed ]
53. PH negru. Consecințele inflamatorii ale stresului psihologic: relația cu rezistența la insulină, obezitatea, ateroscleroza și diabetul zaharat, tip II. Med Ipoteze.2006;67 : 879-891.[ PubMed ]
54. Bhardwaj A, Sethi G, Vadhan-Raj S, Bueso-Ramos C, Takada Y, Gaur U, și colab. Resveratrolul inhibă proliferarea, induce apoptoza și depășește rezistența chimică prin reglarea în jos a proteinelor STAT3 și factorului nuclear-cappaB-reglat antiapoptotic și celulelor de supraviețuire celulară în celulele mielomului multiplu uman.Sânge.2007;109 : 2293-2302.[ PubMed ]
55. Baatout S, Derradji H, Jacquet P, Ooms D, Michaux A, Mergeay M. Apoptoza indusă de radiații a liniilor celulare canceroase după tratamentul cu resveratrol. Int J Mol Med.2004;13 : 895-902.[ PubMed ]
56. Wyke SM, Russell ST, Tisdale MJ. Inducerea expresiei proteazomului în mușchiul scheletic este atenuată de inhibitorii activării NF-kappaB. Br J Cancer.2004;91 : 1742-1750.[ Articol gratuit PMC ][ PubMed ]
57. Tian M, Nishijima Y, Asp ML, Stout MB, Reiser PJ, Belury MA. Modificări cardiace în cazexia indusă de cancer la șoareci. Int J Oncol.2010;37 : 347-353.[ PubMed ]
58. Busquets S, Fuster G, Ametller E, Olivan M, Figueras M, Costelli P, și colab. Resveratrolul nu ameliorează pierderea musculară în diferite tipuri de modele de cașexie pentru cancer.Clin Nutr.2007;26 : 239-244.[ PubMed ]
59. Loprinzi CL, Schaid DJ, Doza AM, Burnham NL, Jensen MD. Modificări ale compoziției corporale la pacienții care au primit greutate în timpul administrării acetat de megestrol.J Clin Oncol.1993;11 : 152-154.[ PubMed ]
60. Fearon KC, Von Meyenfeldt MF, Moses AG, Van Geenen R, Roy A, Gouma DJ, et al. Efectul unei suplimente orale îmbogățite pe bază de proteine și energie densă N-3 asupra pierderii de greutate și a țesutului slab în cazexia de cancer: un studiu randomizat dublu-orb. Intestin.2003;52 : 1479-1486.[ Articol gratuit PMC ][ PubMed ]
61 mai PE, Barber A, D’Olimpio JT, Hourihane A, Abumrad NN. Inversarea pierderilor legate de cancer utilizând suplimentarea orală cu o combinație de beta-hidroxi-beta-metilbutirat, arginină și glutamină. Am J Surg.2002;183 : 471-479.[ PubMed ]
62. Gordon JN, Trebble TM, Ellis RD, Duncan HD, Johns T, Goggin PM. Thalidomidă în tratamentul cazexiei de cancer: un studiu randomizat, controlat cu placebo. Intestin.2005;54 : 540-545.[ Articol gratuit PMC ][ PubMed ]
63. Lundholm K, Gelin J, Hyltander A, Lonnroth C, Sandstrom R, Svaninger G, și colab. Tratamentul antiinflamator poate prelungi supraviețuirea la pacienții subnutriți cu tumori solide metastatice.Cancer Res.1994;54 : 5602-5606.[ PubMed ]
Carcinomul hepatocelular (HCC) este clasat la nivel global drept al șaselea cancer cel mai diagnosticat și al doilea cel mai mortal cancer. Pentru a înrăutăți lucrurile, există doar opțiuni terapeutice limitate disponibile în prezent; prin urmare, este necesar să se găsească un rezervor din care să se poată obține noi tratamente HCC. Domeniul fitomedicinei poate fi soluția la această problemă, deoarece oferă o abundență de molecule derivate din plante, care arată capabilități de a fi eficiente împotriva proliferării, invaziei, migrației și metastazelor HCC. În revizuirea noastră, colectăm și analizăm dovezile actuale cu privire la aceste efecte fitochimice promițătoare asupra HCC și analizăm potențialul lor ca chimioterapii viitoare. În plus, sunt furnizate informații despre semnalizarea din spatele acestor numeroase substanțe fitochimice, în încercarea de a înțelege mecanismele lor. Această revizuire face accesibilă corpul actual de cunoștințe referitoare la fitochimice ca tratamente HCC, pentru a servi drept referință și inspirație pentru cercetări ulterioare în acest subiect.
Abstract
Carcinomul hepatocelular (HCC) este cea mai abundentă formă de cancer la ficat. Reprezintă 75-85% din cazurile de cancer la ficat și, deși se situează la nivel global ca al șaselea cel mai frecvent cancer, ocupă locul al doilea în mortalitatea cauzată de cancer. Decesele cauzate de HCC se datorează, de obicei, răspândirii metastatice a cancerului. Din păcate, există multe provocări și limitări cu cele mai recente terapii și medicamente pentru HCC, ceea ce face dificil ca pacienții să primească îngrijiri care prelungesc viața. Deoarece există în mod clar o cerere mare pentru opțiuni de terapie alternativă pentru HCC, este prudent să apelați la plante pentru soluție, deoarece fitochimicele lor au fost de mult timp folosite și venerate pentru numeroasele lor scopuri medicinale. Această revizuire explorează compușii fitochimici promițători identificați din studiile pre-clinice și clinice care sunt utilizați fie independent, fie împreună cu tratamentele de terapie a cancerului deja existente. Substanțele fitochimice discutate în această revizuire au fost clasificate în mai multe categorii: lipide, polifenoli, alcaloizi, polizaharide, extracte întregi și combinații fitochimice. Aproape 80% dintre compuși nu au reușit să progreseze în studiile clinice din cauza lipsei de informații privind toxicitatea pentru celulele normale și biodisponibilitatea. Deși rămân obstacole mari, fitochimicele pot fi utilizate fie ca terapie alternativă, fie ca terapie integrativă, împreună cu chimioterapiile HCC existente. În concluzie, fitochimicele au un mare potențial ca opțiuni de tratament pentru carcinomul hepatocelular.
Cancerul este a doua cauză de deces la nivel mondial. Conform estimărilor privind incidența și mortalitatea cancerului produse de Agenția Internațională pentru Cercetare a Cancerului (GLOBOCAN 2020), cancerul hepatic este clasat pe locul șase cel mai frecvent diagnosticat de cancer, totuși pe locul al doilea în ceea ce privește rata mortalității [1 ] . Există mai multe tipuri de cancer hepatic, care includ carcinomul hepatocelular, colangiocarcinomul intrahepatic și hepatoblastomul. Dintre aceste subtipuri, carcinomul hepatocelular (HCC) este cel mai răspândit [ 2 ], reprezentând 75–85% din toate cazurile de cancer hepatic diagnosticate [ 3 ]. Datorită absenței unor opțiuni de tratament eficiente, ratele mortalității pentru HCC sunt ridicate, cu o rată actuală de supraviețuire la 5 ani de 8,37% [ 4 ]. Metastazele celulelor canceroase la locurile distale ale corpului este principala cauză de deces la pacienții cu HCC [ 5 ]. Infecțiile cu virusul hepatitei C (VHC) și virusul hepatitei B (VHB) cresc riscul de a dezvolta HCC [ 6 , 7 ]. HCC se poate dezvolta și din cauza bolilor hepatice cronice, cum ar fi ciroza [ 8 ], fibroza [ 9 ] sau boala ficatului gras non-alcoolic (NAFLD) [ 10 ]. În ceea ce privește, aceste boli devin din ce în ce mai răspândite la nivel global, ceea ce duce la așteptarea că ratele HCC vor continua să crească în viitor. Ratele ridicate de mortalitate asociate cu ratele de incidență ridicate reflectă realitatea că HCC devine rapid o problemă majoră de sănătate la nivel mondial.
În prezent, există foarte puține tratamente chirurgicale disponibile pentru HCC. De obicei, HCC în stadiu incipient poate fi tratat prin rezecție chirurgicală, ablație hepatică sau transplant hepatic. Cu toate acestea, fiecare dintre aceste opțiuni de tratament are propriile obstacole și dezavantaje [ 11 ]. Mai mult, odată ce s-a produs răspândirea extrahepatică, aceste terapii scad în eficacitate. Din acest motiv, puțini pacienți cu HCC se califică pentru oricare dintre tratamentele menționate mai sus și, pentru cei care se califică, reapariția cancerului este o preocupare și o posibilitate legitimă [ 12 , 13 , 14 ].
Din punct de vedere farmaceutic, există, de asemenea, puține opțiuni sistemice de tratament medicamentos. Inhibitorii multi-kinazei, cum ar fi sorafenib și lenvatinib, sunt, în general, tratamentele sistematice de primă linie administrate pacienților cu cazuri de HCC nerezecabil [ 15 , 16 ]. Cu toate acestea, aceste medicamente nu sunt caracterizate ca curative și sunt administrate fie ca o încercare de a controla progresia cancerului, astfel încât intervenția chirurgicală să fie posibilă, fie ca o încercare de creștere a speranței de viață; în multe cazuri, cu doar câteva luni [ 17 ]. În plus, este bine cunoscut faptul că pacienții dezvoltă rezistență la aceste medicamente în timp [ 18 , 19 , 20 ], rezultând o eficacitate de scurtă durată. În plus, datorită acestor tratamente, pacienții suferă de leziuni tisulare nețintite și efecte secundare, cum ar fi căderea părului, număr scăzut de celule roșii din sânge, infecție a tractului gastrointestinal și imunosupresie. În prezent, sunt disponibile doar opțiuni limitate de tratament pentru a trata HCC și chiar și acele tratamente nu sunt foarte eficiente. Prin urmare, pacienții care nu răspund la niciunul dintre tratamente nu au altă opțiune decât îngrijirea paliativă [ 21 , 22 , 23 ].
Fitochimice în tratamentul și prevenirea carcinomului hepatocelular
Din punct de vedere istoric, produsele naturale au jucat un rol dominant în evoluția medicinei tradiționale sofisticate, înregistrările disponibile care arată că produsele naturale au fost folosite în medicină încă din 2900 î.Hr. [24 ] . Unul dintre cele mai bine documentate exemple este documentul egiptean antic Ebers Papyrus , care enumeră peste 700 de compuși pe bază de plante care au fost utilizați în scopuri medicinale [ 25 ]. După cum au recunoscut strămoșii noștri cu mult timp în urmă, natura este o sursă bogată de medicamente, care ar putea trata multe afecțiuni.
Deși medicamentele pe bază de plante sunt încă utilizate pe scară largă în întreaga lume, persistă o întârziere vastă în acceptarea validității lor de către comunitatea științifică și medicală din cauza lipsei de dovezi științifice. Studii suplimentare asupra acestor compuși sunt foarte necesare pentru a ajuta la eliminarea decalajului actual dintre cercetarea preclinica și aplicarea clinică, deoarece plantele sunt bogate în metaboliți secundari care s-au dovedit anterior a fi agenți chimioterapeutici și/sau chimiopreventivi puternici utilizați pentru numeroase boli cardiovasculare cronice, boli metabolice, neurodegenerative și neoplazice [ 26 , 27 , 28 ]. Deși există dovezi promițătoare, multor substanțe fitochimice le lipsește soliditatea științifică concretă pentru a putea fi continuate în siguranță la niveluri ulterioare de cercetare, pur și simplu pentru că în prezent nu au fost suficient studiate. În ciuda acestor obstacole implicate în traducerea clinică de succes, multe substanțe fitochimice au alimentat multe progrese în domeniul tratamentului cancerului, inclusiv HCC. Fitochimice dietetice, cum ar fi curcumina [ 29 , 30 ], resveratrol [ 31 , 32 ], quercetină [ 33 ], silibin [ 34 ], N-trans-feruloil octopamină [ 35 ], licopen [ 36 , 37 ], emodin [ 36 , 37 ], 3 39 , 40 ], cofeina [ 41 ] și floretina [ 42 , 43 ] s-au dovedit că posedă proprietăți anti-cancer împotriva HCC. Substanțele fitochimice sunt eficiente în atenuarea principalelor semne distinctive ale cancerului, cum ar fi proliferarea, migrarea și invazia și sunt capabile să inducă apoptoza pentru a controla metastaza HCC prin țintirea markerilor moleculari cheie implicați în aceste procese.figura 1). În plus, conform rapoartelor anterioare, 60% din medicamentele anticancer utilizate în prezent au fost obținute din surse naturale [ 35 ]. De fapt, sursele naturale prezintă aproape 50% din toate entitățile chimice noi aprobate în perioada 2000–2006 [ 44 ].
Substanțele fitochimice și mecanismele lor de a viza metastaza HCC sunt prezentate aici. Substanțele fitochimice afectează procesele majore ale metastazei HCC, cum ar fi ciclul celular, proliferarea, migrarea, invazia și apoptoza prin țintirea markerilor moleculari cheie implicați în aceste procese.
În timp ce unele cercetări au fost efectuate pentru a furniza dovezi pentru eficacitatea tratamentelor derivate din surse vegetale, există mai multe progrese de făcut, deoarece este evidentă necesitatea unor opțiuni alternative sau integratoare în tratamentul cancerului. Produsele naturale derivate din plante ar oferi o disponibilitate largă de opțiuni pentru testarea tratamentelor eficiente pentru HCC. Astfel de produse sunt în mod natural abundente în natură, iar cercetările până în prezent au descoperit că duc la mai puține și mai puține efecte secundare toxice [ 45 ]. Dacă medicina (în domeniul cancerului) va continua să evolueze în viitor, este necesară o explorare și o înțelegere ulterioară a proprietăților și potențialelor fitochimice.
3. Tipuri de fitochimice cu activitate anticanceroasă împotriva HCC
Câțiva compuși naturali derivați din surse vegetale au fost evaluați pentru eficacitatea lor ca potențiale opțiuni de tratament împotriva carcinomului hepatocelular (tabelul 1). În plus, există numeroase substanțe fitochimice testate în studiile clinice pentru tratamentul HCC, ceea ce arată speranța în viitorul fitochimicelor ca agent terapeutic pentru HCC (masa 2). Studiile preclinice asupra claselor majore de molecule bioactive sunt în curs de desfășurare și produc rezultate promițătoare. Și mai promițător este faptul că, mai multe substanțe fitochimice arată că sunt eficiente împotriva diferitelor stadii de metastază, care este principala cauză de deces pentru cei cu HCC (Figura 2). Aici, discutăm efectele anticancer ale mai multor categorii majore de substanțe fitochimice asupra HCC. Studiile discutate în această revizuire au fost obținute căutări amănunțite în baza de date PubMed folosind combinații ale termenilor „carcinom hepatocelular”, „HCC”, „cancer la ficat”, „fitochimic”, „medicament pe bază de plante”, „compuși naturali”, „biomolecule” și „compuși pe bază de plante”.
Fitochimicele vizează markerii de semnalizare moleculari cruciali implicați în diferite procese cheie ale metastazei HCC. Câteva substanțe fitochimice inhibă etapele specifice ale metastazei HCC, cum ar fi tranziția epitelial-mezenchimală (EMT), motilitatea, invazia, intravazația, extravazarea și colonizarea în organe îndepărtate.
tabelul 1
Lista compușilor naturali și mecanismul lor de acțiune asupra carcinomului hepatocelular din studii preclinice.
Inhibarea proliferării prin oprirea ciclului la faza sub-G0/G1, reglarea în sus a markerilor pro-apoptotici Bax și a caspazei clivate 9, cu reglarea în jos a markerului antiapoptotic Bcl-2.
Inducerea apoptozei prin creșterea permeabilității mitocondriilor, ceea ce duce la activarea markerilor pro-apoptotici. Inhibarea exprimării ARNm XIAP, de obicei crescută în celulele canceroase.
Inhibarea EMT prin creșterea expresiei E-caderinei și scăderea expresiei vimentinei. Inhibarea EMT și prin prevenirea activării STAT3 și prin inhibarea expresiei Twist1.
Inhibarea supraviețuirii, invaziei și metastazelor prin suprimarea căilor de semnalizare MEK/ERK și Akt. În plus, împiedică metastaza prin reglarea în sus a E-caderinei.
Inhibarea tranziției epiteliale la mezenchimale prin inhibarea expresiei melcului, vimentinei, MMP2/MMP9 și creșterea expresiei markerului epitelial E-caderinei. Inhibă celulele stem prin inhibarea căii HGF/Met.
Ziyuglicozidă II (3β-3-α-1-arabinopiranosiloxi-19-hidroxiurs-12-en-28-oicacid)
In vitro
HepG2, SMMC-7721
Impiedica proliferarea ciclului celular si provoaca apoptoza prin oprirea ciclului celular la G0/G1. Suprimarea migrației și invaziei prin reglarea în jos a MMP2 și MMP9, inhibând, de asemenea, calea EGFR/NF-kB.
Expresia indusă a supresoarelor tumorale miRNA-29b și p53. Apoptoza indusă prin reglarea în jos a Bcl-2 anti-apoptotic și reglarea în sus a Bax pro-apoptotic, Bac, citocrom-c citosol, caspaza-3 scindată, caspaza-9 scindată și PARP.
Inhibați proliferarea prin blocarea căilor de semnalizare PI3K/AKT și mTOR. Induce apoptoza prin reglarea în jos a proteinelor anti-apoptotice XIAP și supraviețuire și prin reglarea în sus a proteinelor pro-apoptotice scindate caspaza-3 și PARP. Inhibă capacitatea invazivă și migratoare. Inhiba metastazele prin cresterea adeziunii celula-celula.
Scade proliferarea prin oprirea ciclului celular la faza G2 prin suprareglarea p53 și p21. Induce apoptoza prin exprimarea crescută a genelor pro-apoptotice Bcl-2, caspaza 8, caspaza 9, caspaza 3, PARP, receptorul TNF, NF-kB și IkB. Reglează în jos genele anti-apoptotice Birc-8 și BCL212.
Reduce capacitatea metabolică a celulelor canceroase în condiții hipoxice prin reducerea generării de lactat și glucoză; inhibă expresia regulatorului metabolic HIF-1a. Determină diferențierea celulelor canceroase prin activarea HNF-4a, scăzând astfel potențialul metastatic.
Diverse fructe și legume, cum ar fi Kale, fructe de pădure, mere
Quercetină
In vitro In vivo
HepG2
Induce apoptoza prin suprareglarea p53 și Bax. Inhibă glicoliza prin reducerea enzimei cheie de glicoliză HK-2, precum și prin reducerea expresiei mTOR și Akt fosforilate.
Diverse fructe și legume precum căpșuni, mere, castraveți, struguri, ceapă
Fisetin
In vitro
HepG2,
Inhibă proliferarea prin oprirea ciclului celular. Induce apoptoza și necroptoza prin exprimarea crescută a TNFalfa, Bax, caspaza-3 și PARP și prin exprimarea crescută a RIPK1, pRIPK1, RIPK3, pRIPK3 și MLKL; scăderea expresiei NF-kB, pNF-kB și pIKkB.
Scade stemness prin inhibarea căii de semnalizare TGF-beta/SMAD2. Scade capabilitățile invazive prin reglarea în jos a MMP-9 și MMP-1. Suprimă formarea tumorii în modelul de xenogrefă.
Determină citotoxicitate prin creșterea producției de ROS și eliberarea de citocrom-c. Impiedica cresterea prin exprimarea crescuta a miR-6809-5p, care blocheaza activarea regulatorului de semnalizare a celulelor de crestere FLOT1.
Inhibă proliferarea prin inhibarea ERalpha36 și prin inhibarea căilor PI3K/Akt și MAPK/ERK. Induce apoptoza prin activarea caspazei 3 și inducerea buclei de feedback ERα36-EGFR-Her-2.
Inhibă migrarea, invazia și proliferarea prin reglarea în jos a MMP-9 și prin suprimarea semnalizării PI3K-AKT-NF-κB indusă de TPA. Inhiba metastazele in vivo.
Sensibilizează celulele la anoikis prin stimularea mecanismelor pro-apoptotice, cum ar fi inducerea caspazei-9, capzazei-8 și caspazei-3 și prin reglarea în jos a proteinelor anti-apoptotice. Inhibă migrarea prin inhibarea MMP-2 și MMP-9 și prin suprimarea căilor de semnalizare AKT și ERK.
Inhibă proliferarea și induce apoptoza inducând pierderea potențialului membranei mitocondriale și niveluri ridicate de ROS intracelular. Inhibă capacitatea de migrare prin reglarea în jos a markerilor mezenchimale și reglarea în sus a markerului epitelial E-caderinei; reduce MMP-2 și MMP-9.
Inhibă migrarea și invazia prin inducerea opririi ciclului celular G2/M. Induce apoptoza prin reglarea în jos a genelor CCNB1, ceea ce duce la activarea căii p53 prin reglarea în sus a Bax, caspazei-3, caspazei-9, p21 și p53. Suprimă creșterea tumorii și angiogeneza în modelul de șoarece xenogrefă.
Reglarea în jos a expresiei VEGF indusă de hipoxie, inhibă proliferarea și invazia, promovând apoptoza
TACE (7 zile înainte de ginsenozidă): Oxaliplatin (75 mg/m 2 ), 5-fluorouracil (500 mg/m 2 ), epirubicin (30–50 mg/m 2 ), administrat până când a rămas în vase mai mult de 10 bătăi ale inimii, ginsenozidă: 20 mg de două ori pe zi
Oral
Combinația TACE și ginsenozidă Rg3 poate prelungi supraviețuirea globală în comparație cu TACE singur.
Activitate de modulare a imunității prin căile IL-6/Jak2/Stat3, celulele ucigașe naturale, celulele T, IFN-γ și celulele supresoare derivate din mieloid
600 mg sau 800 mg de două ori pe zi
Oral
Activitățile de modulare imunitară ale Icaritinei au fost corelate cu profilurile de siguranță și cu beneficiile preliminare de supraviețuire.
Pacienților cu un singur program de chimioterapie anterior li sa permis Niciunul
Faza II Faza I
Interferența sintezei ADN-ului
125 mg/m 2 săptămânal Începând cu 7,5 mg/m 2 /zi cu o creștere de 2,5 mg/m 2 /zi
Nespecificat Rezervor de cateterism percutanat într-un buzunar subcutanat
Irinotecanul a prezentat activitate modestă asupra HCC cu o toxicitate substanțială la pacienți. Perfuzia intra-arterială de Irinotecan nu a avut evenimente adverse majore de la compus, dar a avut preocupări legate de managementul dispozitivului intra-arterial.
Ginseng, atractylodes, Tuckahoe, rădăcină de lemn dulce, radix bupleuri, yam, cortex moutan, salvia miltiorrhiza , turmeric, rhizome zehoariae
Terapia Jianpi Huayu
Hepatectomie
Studiu clinic randomizat
Îmbunătățește funcția imunitară
Ginseng 20 g, atractylodes 15 g, Tuckahoe 15 g, rădăcină de lemn dulce 6 g, radix bupleuri 15 g, yam 12 g, cortex moutan 10 g, salvia miltiorrhiza 15 g, turmeric 10 g, rhizoma zedoariae 10 g, administrat o dată pe zi, 10 g, zile după hepatectomie
Nu este specificat
Terapia Jianpi Huayu și hepatectomia împreună au scăzut reapariția postoperatorie și metastazele, în timp ce supraviețuirea fără boală și supraviețuirea generală s-au îmbunătățit.
Rădăcină de pilose Asiabell, rizom Largehead Atractylodes, tubercul de Pinella, fructe de păducel, rizom de igname comun, Fu-ling, coajă de arecă, rădăcină de lemn dulce, iarbă de pelin Virgate. Rădăcină de pilose Asiabell, rizom Atractylodes cu capul mare, tubercul de Pinella, fructe de păducel, rizom de igname comun, Fu-ling, coajă de arecă, rădăcină de lemn dulce, iarbă de pelin Virgate
TACE: 5-fluorouracil (1000 mg/m 2 ), cisplatină (80 mg/m 2 ) și decoct Jianpi Ligan (administrat în aceeași zi cu TACE): Radix Codonopsis (20 g), Rhizoma Atractylodis macrocephala (10 g), Poria cocos (15 g), Radix Glycyrrhizae (5 g), Rhizoma Diosscoreae (15 g), Rhizoma Pinelliae (10 g), Fructus Crataegi (15 g), Semen Nelumbinis (20 g), Herba Artemisia scoparia (50 g), Pericarpium Arecae (25 g) decoct Jianpi Ligan 100 ml pe cale orală o dată pe zi, la 30 de minute după mese, compus din: rădăcină de pilose Asiabell (20 g), rizom de Atractylodes de capul mare (10 g), Fu-ling (15 g), rădăcină de lemn dulce ( 5 g), Rizom de igname comun (15 g), Tubercul Pinellia (10 g), Fructe de păducel (15 g), Semen Nelumbinis (20 g), Iarbă de pelin Virgate (50 g), Coji de Areca (25 g)
Nu este specificat Oral
TACE și decoctul Jianpi Ligan combinate au arătat o reducere a efectelor secundare și a îmbunătățit supraviețuirea pe termen lung la pacienții cu HCC nerezecabil tratați cu TACE. RFA și decoctul Jianpi Ligan combinate au arătat o rată semnificativ mai mare de succes a tratamentului și o supraviețuire generală de 3 ani semnificativ mai mare în comparație cu martorii.
Rădăcină de Salvia chinensis, rădăcină de Actinidia valvata, Cremastra appendiculata Rădăcină de Salvia chinensis, rădăcină de Actinidia valvata, Cremastra apendiculare, rădăcină de Salvia chinensis, ruber de Pseudobulbus cremastrae
Granule Jiedu Granule Jiedu + rădăcină de Salvia chinensis Benth + tubercul de Pseudobulbus cremastrae seu Pleiones + rădăcină de Gallus domesticus Brison + extract din Buro gargarizans Cantor
Studiu clinic retrospectiv Studiu controlat randomizat
Mecanismul exact nu este clar Inhibarea creșterii celulelor tumorale, inducerea apoptozei, suprimarea angiogenezei, creșterea funcției imune
TACE: lipiodol (10–20 ml) și pirarubicină (30 mg) evaluate prin radiografie, izodoză de radiochirurgie Gamma Knife (GKR) de 55%, 5,9 g granule Jiedu de două ori pe zi după 1 săptămână de TACE Medicina tradițională pe bază de plante (THM) folosind Cinobufacini (50 ml) administrat o dată pe zi, granulă Jiedu (4,5 g) administrat de două ori pe zi și TACE: pirarubicină (10 mg), mitomicină (10 mg), iodipină (2 până la 5 ml)
Granula Jiedu combinată cu TACE și Gamma Knife Radiosurgery este sigură pentru pacienții cu HCC cu tromboză tumorală a venei portă. Granula Jiedu a promovat și un prognostic mai bun. Medicina tradițională pe bază de plante a fost superioară TACE pentru prevenirea recidivei bolii și prelungirea supraviețuirii globale în HCC mic.
Lipiodol marcat cu iod-131 intra- arterial Lipiodol marcat cu 131I Chemoembolizare cu Lobaplatin amestecat cu Lipiodol (ulei iodat)
Rezecție Niciuna Transplant hepatic ortotopic
Studii prospective randomizate Studiu clinic retrospectiv controlat Studiu clinic retrospectiv controlat
Eradică locurile tumorale microscopice Eradică focarele microscopice tumorale Induce ischemia tumorală și permite concentrarea locală mare de chimioterapie
2 ml de iod-131 lipiodol la 1850 MBq per tratament 5 ml volum de iod-131 lipiodol la o doză de 2,2 GBq Lobaplatină (50 mg/m 2 ), ulei iodat (cuprinzând între 2 ml și 5 ml)
Canularea arterei Intra-arterial Pompă subcutanată și cateter
Lipiodolul marcat cu iod-131 intra-arterial după rezecție a scăzut rata de recurență și a crescut supraviețuirea fără boală și supraviețuirea globală la pacienții cu HCC. Injecția intra-arterială de iod-131 sa dovedit a fi sigură și a avut beneficii pentru supraviețuirea globală la pacienții cu HCC avansat. Chemoembolizarea pe bază de lobaplatină poate îmbunătăți supraviețuirea globală și poate provoca un răspuns tumoral cu efect pentru pacienții cu HCC nerezecabil.
Administrat sub formă de perfuzii de 1 oră în zile selectate și doze crescânde de: 70 mg/m 2 , 80 mg/m 2 , 90 mg/m 2 și 100 mg/m 2
Pe cale intravenoasă
Un pacient a obținut remisie parțială și alți 9 pacienți au avut o boală stabilă prelungită. A existat, de asemenea, o îmbunătățire a simptomelor legate de tumoră.
Rădăcină Pseudostarwort, rădăcină milkvetch, poria, tubercul alb de atratylodes, rădăcină de rehmannia, dendrobium, rădăcină de bujor alb, rădăcină de angelica chinezească, plantă de aldenlandia, lobelia chinezească, ghimbir sălbatic, rădăcină de lemn dulce
Amestecul Shenqi
Coagularea la microunde
Studiu clinic retrospectiv
Stimularea limfocitelor, stimularea macrofagelor, creșterea imunității umorale, îmbunătățirea imunității celulare, reducerea transaminazelor glutamice piruvice, îmbunătățirea sistemului reticuloendotelial, hepatoprotector, antiinflamator
Coagulare la microunde: putere de ieșire 60 W sau 800 de secunde, amestec Shenqi (dată a doua zi după coagularea cu microunde): 20 ml (20 g de compuși bruti), de 3 ori pe zi
Oral
Amestecul Shenqi și coagularea la microunde au ucis celulele tumorale și au prevenit reapariția, precum și a îmbunătățit imunitatea celulară fără reacții adverse.
Preveniți proliferarea celulelor canceroase, îmbunătățiți proliferarea limfocitelor T și B
40 ml de injecție Xiaoaiping
Picurare intravenoasă
Injecția de Xiaoaiping a îmbunătățit calitatea vieții la pacienții cu HCC avansat, a îmbunătățit imunitatea și a prelungit supraviețuirea fără progresie.
Peptidă polizaharidă izolată din Yunzhi/kawaratake
Nici unul
Studiu controlat randomizat
Inhibă eliberarea IL-17F, menține nivelurile de prolactină, crește TRAIL R1
Coriolus versicolor (2,4 g/zi)
Sugerat ca intravenos
Nu există nicio diferență de timp până la progresie la utilizarea Coriolus versicolor (Yunzhi), dar pacienții cu HCC au avut o calitate mai bună a vieții în comparație cu placebo. Coriolus versicolor (Yunzhi) ar trebui explorat în continuare pentru îngrijirea paliativă.
Lipidele sunt o clasă de compuși biochimici, formați în principal din acizi grași. S-a demonstrat că diferite lipide și derivații lor din plante au potențial anti-cancer împotriva HCC. Amaranthus spinosus , altfel cunoscut sub numele de amarant spinos, este o plantă originară din climatele tropicale calde, inclusiv din America tropicală. Este considerată o buruiană invazivă în anumite medii, dar o legumă cu frunze în altele [ 132 ] și a fost folosită medicinal în regiunea Sub-Himalaya din India [ 133 ]. Studiile au arătat utilizarea potențială a medicamentului pentru icter [ 133 ], diabet [ 134 ], diureză [ 135 ], constipație gastrointestinală [ 136 ] și infecții bacteriene [ 137 ]. Până de curând, au fost publicate puține studii cu privire la proprietățile sale anticancerigene împotriva carcinomului hepatocelular. Într-un studiu, (14E, 18E, 22E, 26E)-metil nonacosa-14, 18, 22, 26 tetraenoat, un acid gras izolat din A. spinosus , a demonstrat mai întâi capacitatea sa puternică de a inhiba proliferarea celulelor HepG2 HCC și apoptoza prin suprareglare. de Bax și reglarea în jos a Bcl-2 și ciclinei B1, atunci când sunt administrate la o doză de 25,52 µmol/L [ 46 ]. De asemenea , IC50 al doxorubicinei pentru aceeași linie celulară a fost, de asemenea, găsit în acest studiu a fi 24,68 pmol/L. Acest lucru sugerează că doza pentru acidul gras din Amaranthus spinosus este fezabilă de atins în tratamentele din lumea reală, deoarece este comparabilă cu doza de doxorubicină utilizată în prezent pentru chimioterapie.
Lipiodolul (un ulei derivat din semințele de mac) conține 38% iod în greutate [ 138 ] și este curățat din celulele HCC mai puțin rapid decât este curățat din hepatocitele normale. Acest compus poate fi, de asemenea, făcut să fie radioactiv, rezultând lipiodol marcat cu iod-131. Aceste caracteristici ale lipiodolului permit ca tratamentul radioactiv al lipiodolului marcat cu iod-l31 să fie utilizat ca instrument de tratament al HCC. Atunci când este administrat pacienților, permite țesuturilor hepatice canceroase să primească o doză de radiații de aproximativ opt ori mai puternică decât țesutul hepatic normal, ceea ce reprezintă o modalitate ideală de a viza țesuturile canceroase specifice din cadrul pacienților [125 , 138 ] . Deși a fost utilizat inițial ca mediu de contrast pentru utilizarea radiologiei, mai multe studii clinice au găsit succes în utilizarea lipiodolului marcat cu iod-131 ca tratament capabil de a crește supraviețuirea globală a pacienților cu HCC [125 , 126 , 127 ] . Acest tratament s-a dovedit a fi bine tolerat și chiar eficient în tumorile mici [ 125 ]. Lipiodolul este un bun exemplu de fitochimic care ajută la furnizarea unei căi pentru tratamentul HCC, în ciuda faptului că este utilizat doar ca purtător de radiații. Este important de remarcat faptul că fitochimicele nu trebuie neapărat să omoare definitiv celulele canceroase pentru a putea fi utilizate în continuare pentru a combate cancerul prin diferite mecanisme.
Terpenoide
Terpenoidele sunt lipide nesaponificabile sau lipide simple care se găsesc în multe plante și ciuperci din întreaga lume. Ele sunt formate din numere diferite de unități izoprene legate și pot fi clasificate ca monoterpene, diterpene, triterpene, sesquiterpene etc., în funcție de numărul de unități izoprene prezente în moleculă [139 , 140 , 141 , 142 ] . Terpenoizii sunt una dintre cele mai mari clase de fitochimice [ 143 ] care s-au dovedit a avea efecte anticancerigene împotriva diferitelor tipuri de cancer, inclusiv carcinomul hepatocelular [ 144 , 145 ]. Ciupercile sunt deosebit de bogate în terpenoide [ 145 ], iar extractele lor sunt în prezent explorate ca potențiale produse farmaceutice [ 146 ], unele ciuperci fiind deja utilizate în cadrul clinic [ 146 , 147 , 148 ]. Astrakurkurone (o triterpenă izolată din ciuperca comestibilă Astraeus hygrometricus ) și-a demonstrat potențialul de utilizare ca agent citotoxic împotriva liniilor celulare de HCC (Hep3B și HepG2) la doze deosebit de mici (DL50 de 58,8 µM și respectiv 122 µM) [47 ] . De asemenea, s-a constatat că toxicitatea este selectivă, deoarece nu a fost găsită nicio citotoxicitate atunci când a fost administrată la o linie celulară hepatică normală, chiar și la cea mai mare doză testată (250 pM). Astrakurkurona a împiedicat proliferarea celulară prin oprirea ciclului celular în faza sub-G0/G1 și a arătat dovezi de depolarizare a membranei mitocondriale prin producția excesivă de ROS și întreruperea potențialului membranei mitocondriale [47 ] . Compusul a reglat, de asemenea, markerii pro-apoptotici, au scindat caspaza-9 și Bax, în timp ce a reglat în jos markerul anti-apoptotic, Bcl-2. Acest studiu a demonstrat că triterpenul astrakurkurone este un potențial fitochimic anti-cancer pentru tratamentul HCC.
Fructele conțin și terpenoide care demonstrează efecte anticancerigene. Ziziphus jujuba , un fruct care seamănă cu curmalul, aparține familiei Rhamnaceae ; este cultivat în Asia, Australia, Europa și regiunile mediteraneene. Acest fruct conține diferiți compuși bioactivi, inclusiv mulți acizi triterpenici diferiți [ 149 ]. Un studiu anterior al lui Kim și colab., a indicat că acidul ursolic, unul dintre acizii triterpenici ai Ziziphus jujuba , a scăzut viabilitatea celulară și a prezentat activitate pro-apoptotică împotriva celulelor HepG2 (când este administrat la o doză de 30 µM) [ 48 ]. Deși mecanismul exact nu a fost încă elucidat, se sugerează că acidul ursolic (UA) este capabil să inducă oprirea ciclului celular prin întreruperea formării furculiței ADN în timpul etapei de inițiere a ciclului celular [48 ] . Mai mult, UA determină o expresie crescută a p21/WAF1, care poate provoca, de asemenea, oprirea ciclului celular și poate induce apoptoza prin eliberarea citocromului c și a activării caspazei-3 [48 ] . Un alt studiu a demonstrat, de asemenea, efectele antitumorale ale UA folosind celulele HCC Huh-7 [ 50 ]. S-a demonstrat că acidul oleanolic (OA), un alt acid triterpenic, împreună cu UA, induc apoptoza (la o doză de 20 µM) acționând asupra permeabilității mitocondriilor, ducând la eliberarea citocromului C și la activarea markerilor pro-apoptotici caspazei. -9, caspaza-3 și PARP. Studiul a arătat, de asemenea, că OA și UA ar putea inhiba expresia inhibitorilor X-linkați ai mARN-ului proteinei apoptotice (XIAP), care este de obicei crescut în celulele canceroase [ 50 ]. În plus, atunci când OA a fost injectată subcutanat (1,0 g/kg) la șoareci sau șobolani, nu a provocat nicio toxicitate sau mortalitate [ 150 ]. În plus, după administrarea orală de OA (la 180 mg/kg) la șoareci timp de 10 zile, nu s-au observat anomalii în creier, inimă, ficat, rinichi, tiroidă, testicule, stomac, intestin sau splină [150 ] . De asemenea, s-a demonstrat că OA și UA induc apoptoza într-o manieră dependentă de doză (2–8 µmol/L) în diferite linii celulare de cancer hepatic uman prin creșterea fragmentării ADN-ului, scăderea activității Na+-K+-ATPazei, scăderea membranei mitocondriale. potențialul, creșterea activității caspazei-3 și caspazei-8 și suprimarea producției de proteină de semnalizare angiogenă VEGF și molecule de adeziune celulară ICAM-1 [ 151 ]. Eficacitatea la astfel de doze mici este o caracteristică promițătoare, care ar putea permite o tranziție mai ușoară și mai sigură la practica clinică. Cu cât este mai mică doza necesară, cu atât mai fezabilă poate fi realizarea unui astfel de nivel de dozare la om.
Ginsenozide de tip diol sunt un alt triterpenoid major izolat din Panax ginseng ; o plantă medicinală tradițională chinezească care a fost folosită de mult timp ca remediu antiinflamator, anticancer și anti-patogen [ 152 ]. Deși P. ginseng este compus din multe componente active, una dintre aceste componente (o ginsenozidă), în special, protopanaxadiolul (PPD), s-a dovedit a fi capabilă să exercite efecte antioxidante și antiinflamatorii asupra carcinomului hepatocelular HepG2 și PLC/PRF/5. celule la tratamente de 20 și 40 pM [ 51 ]. În plus, PPD are potențialul de a inhiba migrarea, invazia și proliferarea acestor celule canceroase prin creșterea nivelului de expresie al markerului epitelial E-cadherină și scăderea nivelului de expresie al markerului mezenchimal vimentin. PPD își poate prezenta, de asemenea, efectele sale anticancerigene prin țintirea căii STAT3/Twist1 [ 51 ]. STAT3 este un jucător cheie în proliferare, migrare și angiogeneză. De obicei, STAT3 este prezent în citoplasmă într-o stare inactivă până când este stimulat de citokine, prin care se translocă în nucleu și devine activ. PPD inhibă translocarea STAT3 în nucleu. Twist1, o genă țintă a STAT3, este, de asemenea, afectată de PPD. S-a descoperit că supraexpresia Twist1 induce EMT a celulelor canceroase. În plus, inactivarea STAT3 de către PPD împiedică capacitatea lui Twist1 de a induce EMT pentru a controla metastazele [ 51 ]. PPD are, de asemenea, un mare potențial de a fi utilizat sinergic cu chimioterapiile existente. Atunci când a fost administrat intragastric la șoareci nuzi BALB/C femele cu xenogrefe de celule canceroase, tratamentul combinat de oxaliplatină (OXA) cu platină cu PPD (30 mg/kg PPD + 10 mg/kg OXA) s-a dovedit a fi semnificativ eficient în suprimarea tumorii. creștere, în comparație cu tratamentul cu PPD (30 mg/kg) sau OXA (10 mg/kg) în monoterapie [ 51 ]. Capacitatea de a spori efectele chimioterapeutice ale medicamentelor anticancer existente este puternică, mai ales dacă fitochimicul are mai puține efecte secundare decât medicamentul anticancer. Acest lucru ar putea permite administrarea de doze mai mici de medicamente anticancer și, prin urmare, ar putea duce la mai puține efecte secundare dăunătoare la pacienți. Cu toate acestea, cercetarea asupra efectelor secundare care pot fi cauzate de administrarea PPD este încă destul de departe. În ciuda acestui fapt, promisiunea de bază pe care PPD o aduce la masă este grozavă.
Ardipusilloside I este o triterpenă izolată din planta medicinală chineză Ardisia pusilla A. DC. Acest compus exercită efecte anti-cancer împotriva liniilor de celule HCC HepG2 și SMMC-7721 prin inhibarea severă a supraviețuirii, invaziei și metastazei acestora (când este administrat la o concentrație de 50 pM). Se crede că aceste efecte sunt realizate prin suprimarea căilor de semnalizare MEK/ERK și Akt, ducând la reglarea în jos a metaloproteinazei matriceale-2 (MMP-2) și a metaloproteinazei matriceale-9 (MMP-9). Mai mult, s-a demonstrat că ardipusilozida I reduce metastaza celulelor HCC prin activarea Rac1, ceea ce duce la reglarea în sus a E-caderinei, ducând ulterior la capacități metastatice reduse. Experimentele in vivo au prezentat, de asemenea, efectele anticancer ale Ardipusilloside I. Două grupuri de șoareci masculi atimici BALB/c nuzi au fost implantate ortotopic cu celule HCC și tratate cu ardipusilozidă I în două doze diferite (50 mg/kg/zi și 100 mg/zi). kg/zi). Ambele tratamente au condus la animalele care au dezvoltat mai puține tumori metastatice în plămâni, comparativ cu martorii netratate. Aceste dovezi sugerează capacitatea ardipusilozidei I de a avea un impact negativ asupra capacității metastatice a HCC [ 52 ]. Mai mult decât atât, dozele testate în aceste modele animale pot fi fezabile pentru a fi traduse la oameni. Când se trece de la studiile preclinice la studiile clinice, necesarul de doză va scădea de obicei în funcție de greutatea corporală și metabolismul animalului [ 153 ]. Deoarece ardipusilozida I s-a dovedit că are efecte netoxice asupra astrocitelor normale [ 154 ], acest compus prezintă un mare potențial de a deveni un agent terapeutic împotriva HCC.
Lanatozidul C sau isolanidul este o cardioglicozidă triterpenică extrasă din planta Digitalis ferruginea (și anume „global ruginit”). S-a demonstrat că creșterea celulelor Mahlavu HCC cu deficit de PTEN a fost redusă semnificativ la tratamentul cu lanatozidă C (la 2 uM) prin inducerea opririi ciclului celular G2/M, activarea JNK, inducerea apoptozei și producția de ROS in vitro. În plus, prin RMN-ul șoarecilor nuzi (8-16 săptămâni), s-a demonstrat că lanatozida C reduce dimensiunea tumorilor xenogrefe HCC in vivo cu aproape 50%, după administrarea prin hrănire prin gavaj (6 mg/kg) timp de 21 de zile. 53 ]. Este încurajator să se observe astfel de efecte in vivo după administrarea prin gavaj, deoarece aceasta indică faptul că compusul poate fi eficient chiar și atunci când este administrat oral. Administrarea orală este mai sigură în comparație cu alte forme, deoarece ajută la scăderea riscului de citotoxicitate [ 155 , 156 ]. Deși trebuie făcute mult mai multe cercetări, lanatozida C pare să aibă efecte benefice prin ceea ce este în general considerat o cale mai sigură de administrare a medicamentelor. În plus, atunci când a fost administrat la linii celulare embrionare și hepatice normale (0,1 până la 500 µM timp de 24 de ore), nu a fost observată nicio toxicitate [ 157 ].
Diospyros kaki (cunoscut și sub numele de curmal kaki) are o abundență de fitochimice de oferit, deoarece este bogat în terpenoide și flavonoide. S-a demonstrat că extractele de etanol din frunzele de D. kaki reglează EMT și caracteristicile stemness ale HCC prin inhibarea căii factorului de creștere a hepatocitelor (HGF)/Met. Liniile de celule HepG2, Hep3B și SNU475 tratate cu extract etanolic de D. kaki (50 µg/mL) au indicat o expresie mai scăzută a CD44 și keratinei19, precum și o expresie mai scăzută a melcului, vimentinei, MMP-1 și MMP-2/MMP -9. Simultan, a fost observat un nivel crescut de expresie al E-caderinei după tratamentul cu extract de frunze de HGF+ D. kaki , sugerând că D. kaki are capacitatea de a inhiba creșterea celulelor canceroase în ciuda adăugării de HGF la celule. În plus, acest studiu a testat eficacitatea tratamentului de chimioterapie standard de aur Sorafenib în combinație cu 50 µg/mL de D. kaki și a constatat că IC 50 a lui Sorafenib a scăzut de la 3,7 µg/mL, 5,9 µg/mL și 9,5 µg/mL în liniile de celule canceroase Hep3B, HepG2 și, respectiv, SNU475, la 1,8 pg/mL, 2,7 pg/mL și 5,9 pg/mL. Nu s-a descoperit că extractul de frunze de kaki provoacă toxicitate în celulele canceroase la concentrații de până la 50 µg/mL, care a fost doza maximă testată. Mai mult, a fost efectuat un test de citotoxicitate pe șoareci femele ICR în vârstă de 5 săptămâni, cărora li s-au administrat doze orale de extract de Kaki la 2 g/kg o dată pe zi timp de 14 zile. LD50 nu a putut fi determinată din acest experiment, deoarece toți șoarecii au supraviețuit toate cele 14 zile fără nicio modificare a greutății corporale în comparație cu martorii . În plus, atunci când au fost testate pe linii celulare umane normale (MCF10A și HPL1A), nu a fost observată nicio citotoxicitate [ 158 ]. Deși este benefic și promițător faptul că extractul nu s-a dovedit a fi toxic pentru celule in vitro și nici pentru întregul organism in vivo, poate fi șocant să auzi că un tratament anti-cancer prospectiv nu are efecte citotoxice asupra liniilor celulare canceroase. Cu toate acestea, extractul de D. kaki pare să aibă un efect net pozitiv, deoarece scade potențialul metastatic al celulelor canceroase [ 54 ]. Metastaza este o complicație potențială substanțială în orice cancer și este motivul pentru care mulți pacienți nu supraviețuiesc mult timp după diagnostic, mai ales în cazul HCC. O substanță fitochimică care arată impact asupra metastazelor, deși nu arată în mod direct citotoxicitate față de celulele canceroase, poate încă îmbunătăți viața și rezultatele multor pacienți cu cancer.
Sanguisorba officinalis este o plantă din familia Rosaceae , care crește în regiunile mai reci din nordul Europei. O saponină triterpenoidă numită Ziyuglicozidă II (3β-3-α-1-arabinopiranosiloxi-19-hidroxiurs-12-en-28-oicacid) se găsește în rădăcinile acestei plante. Ziyuglicozida II este bine cunoscută pentru proprietățile sale anticancerigene, inclusiv având un efect de ameliorare asupra carcinomului gastric [ 159 ], cancerului de sân [ 160 ] și cancerului de colon [ 161 ]. Un alt studiu asupra Ziyuglycoside II a indicat că este, de asemenea, eficient în împiedicarea creșterii HCC în liniile de celule canceroase HepG2 și SMMC-7721 prin creșterea activității ROS și inducerea opririi ciclului celular la G0/G1, ceea ce duce la apoptoză. Mai mult, s-a observat că acest compus scade capacitățile migratoare și invazive ale acestor celule HCC într-o manieră dependentă de doză (6,6 µg/mL–10 µg/mL), prin suprimarea expresiei MMP-2 și MMP-9, precum și ca prin inhibarea căii EGFR/NF-kB [ 55 ]. În plus, Ziyuglycoside II a prezentat un nivel scăzut de toxicitate pentru celulele normale (BRL3A și HEK293T), indicând că citotoxicitatea sa este selectivă pentru HCC [ 55 ].
Brucea javanica este un arbust înflorit al cărui fruct a fost mult timp folosit în medicina tradițională chineză pentru tratamentul dizenteriei, malariei și chiar cancerului [ 162 ]. Fructul este foarte bogat în quasinoide, care sunt lactone triterpenice cunoscute pentru că posedă o activitate antitumorală semnificativă. Uleiul de Brucea javanica (BJO) este obținut din extracte din semințele de Brucea javanica . Acest ulei conține mulți quasinoizi, și anume brusatol, care este unul dintre cei mai importanți quasinoizi izolați din B. javanica [ 162 ]. Efectele anti-cancer promițătoare ale Brusatol au fost raportate în multe tipuri de cancer, inclusiv cancerul colorectal [ 162 ], pulmonar [ 163 ] și cancerul pancreatic [ 164 ]. În plus, efectul citotoxic al brusatolului a fost testat și pe celulele epiteliale gastrice normale și sa dovedit a fi netoxic [ 164 ]. Când BJO îmbogățit cu brusatol (BE-BJO) la 610 mg/kg, BJO obișnuit (BJO) la 2000 mg/kg și BJO fără brusatol (BF-BJO) la 2000 mg/kg a fost injectat intraperitoneal la șoarecii Kunming masculi cu H22 implantat. hepatoame celulare, s-a constatat că creșterea tumorii a fost suprimată la acei șoareci tratați cu BE-BJO. În plus, analiza ARN a arătat că BE-BJO a indus în mod semnificativ expresia miRNA-29b și p53. BJO și BE-BJO au diminuat Bcl-2 și au îmbunătățit expresia Bax, Bad, citosol citocrom-c, caspaza-3 scindată, caspaza-9 scindată și PARP pentru a induce apoptoza xenogrefelor H22 la șoareci. Pe baza acestor constatări, este clar că brusatol (un compus activ major din BJO) exercită o moarte semnificativă a celulelor HCC [ 56 ]. În mod benefic, s-a observat, de asemenea, că administrarea a 2000 mg/mL din fiecare dintre BJO, BE-BJO și BF-BJO pe cale orală timp de 14 zile nu s-a dovedit toxică pentru șoarecii Kunming, deoarece nu au existat decese sau manifestări comportamentale de toxicitate. Speculăm că terapia combinată cu brusatol izolat din Fructus Bruceae , administrată cu terapiile existente pentru HCC, va îmbunătăți durata de viață a pacientului pe baza efectelor observate în cercetările anterioare efectuate prin care cisplatina administrată în prezența brusatolului a demonstrat o opțiune terapeutică sinergică pentru cancerul de colon. 162 ].
Artemisinina este un compus de lactonă sesquiterpenic găsit în pelin dulce ( Artemisia annua ), un tip de plantă cu flori care este originară din Asia temperată și părți ale Americii de Nord. Aceste plante medicinale sunt adesea folosite în medicina tradițională chineză pentru tratamentul diferitelor boli, inclusiv febră și hemoroizi, deși potențialul clinic al acestor plante cu flori se extinde mult mai departe decât utilizarea lor actuală. Compușii de artemisinină izolați din Artemisia au arătat de mult timp proprietăți anticancerigene împotriva diferitelor tipuri de cancer [ 165 ], iar aceste efecte anticancerigene se extind și la HCC. S-a demonstrat că derivații de artemisinină inhibă proliferarea celulelor HepG2 și Huh-7 (când sunt administrați la 100 pg/mL) prin blocarea căilor de semnalizare PI3K/AKT și mTOR în aceste linii celulare HCC. Aceste căi reglează atât apoptoza, cât și angiogeneza în HCC. Blocarea unor astfel de căi controlează potențialul proliferativ și angiogenic al celulelor HCC. S-a demonstrat, de asemenea, că artemisinina reglează în jos proteinele anti-apoptotice XIAP și survivin, precum și că crește expresia caspazei-3 și PARP scindate. Aceste acțiuni duc la creșterea apoptozei Huh7 și HepG2 [ 57 ]. În plus, sferoizii tumorali tratați cu derivați de artemisinină au prezentat apoptoză și necroză a celulelor tumorale la rate mai mari decât sferoizii de control al HCC netratați cu derivați de artemisinină, indicând potențialul citotoxic eficient al artemisininei împotriva HCC. În plus, artemisinina s-a dovedit a fi eficientă în inhibarea capacităților invazive și migratoare ale celulelor HCC, așa cum se arată când celulele HCC tratate cu artemisinină (la 12,5 µM) au prezentat motilitate minimă în comparație cu celulele de control HCC [58 ] . Rezultatele in vivo au arătat și potențialul terapeutic al artemisininei. S-a constatat că șoarecii masculi atimici nuzi BALB/c care au fost implantați ortotopic cu țesuturi tumorale HCC și apoi tratați intragastric cu artemisinină zilnic (la 50 mg/kg și la 100 mg/kg timp de 4 săptămâni) s-au dovedit a avea mai puține tumori care au metastazat la plămâni. , în comparație cu șoarecii martor netratați. În plus, când a fost testată cu un test de aderență celulară, artemisinina a crescut semnificativ aderența celulă-celulă a celulelor HCC, indicând în continuare efectele prohibitive ale artemisininei asupra capacității HCC de a metastaza [58 ] .
Paclitaxelul este un diterpenoid izolat din arborele Pacific Yew, care este utilizat în prezent ca agent citotoxic împotriva tumorilor solide [ 128 ]. Principala sa metodă de acțiune implică țintirea celulelor în diviziune pentru a induce citotoxicitatea. Paclitaxelul a demonstrat activitate care acționează împotriva tumorilor în cancerele pulmonare, capului, gâtului și esofagiei, precum și asupra remisiilor induse [ 166 ]. Acest fitochimic a avut succes în studiile clinice și este acum un tratament aprobat de FDA pentru cancerul de sân, cancerul pulmonar, cancerul ovarian și sarcomul Kaposi [ 166 ]. Într-un studiu clinic care a explorat potențialul de a utiliza paclitaxel pentru tratamentul HCC, s-a constatat că există o îmbunătățire a simptomelor legate de tumoră. Sunt necesare mai multe cercetări pentru a explora posibilitățile de utilizare a paclitaxelului pentru a trata HCC.
Ginsenoside Rg3 este o saponină de ginseng obținută din planta de ginseng Panax care s-a dovedit că atenuează căile de semnalizare privind Akt/eNOS dependentă de VEGF [ 167 ]. Mai mult, ginsenozidul Rg3 reduce fosforilarea STAT3 rezultată din condițiile hipoxice, fosforilarea ERK 1/2, JNK și scade expresia VEGF [ 168 ]. S-a demonstrat că ginsenozidul Rg3 inhibă activarea formării vaselor microtumorale la șoareci atunci când este administrat la o doză de 10 mg/kg o dată pe zi și a contribuit la contracția tumorii și la creșterea supraviețuirii animalelor [169 ] . Un studiu care a evaluat farmacocinetica Ginsenoside Rg3 a constatat că acesta a fost bine tolerat de oameni atunci când este administrat intramuscular la o doză cuprinsă între 10-60 mg o dată la două zile [170 ] .
3.2. Fenolii
Fenolii reprezintă o mare parte din toți metaboliții secundari produși de plante; peste 8000 de fenoli au fost identificați în natură până acum [ 171 , 172 , 173 ]. Aceste substanțe fitochimice sunt compuse dintr-un inel aromatic cu una sau mai multe grupe hidroxil și se găsesc într-o gamă largă de plante, inclusiv diferite legume, fructe și leguminoase [172 , 173 ] . Fenolii pot fi clasificați în continuare în compuși, cum ar fi flavonoide, catechine, stilbene și xantone [ 172 , 173 ].
3.2.1. Flavonoide
Flavonoidele sunt prezente în fructe, legume, cereale, ceaiuri și vinuri, constituind o parte importantă și esențială a dietei umane [ 174 ]. Acești compuși au fost recunoscuți pentru că posedă proprietăți antiinflamatorii, antioxidante, antibacteriene și antivirale [ 174 , 175 ]. Deloc surprinzător, diferitele flavonoide au demonstrat și potențialul ca compuși anticancerigen.
Citrus bergamia , cunoscută și sub numele de bergamotă, este un copac originar din regiunea Calabria italiană. Se știe că sucul din citricele acestui pom este compus în mare parte din flavonoide, cele mai abundente flavonoide din suc fiind eriocitrina, neoeriocitrina, naringina și neohesperidina [59 ] . Atunci când sunt extrase și testate individual, s-a demonstrat că aceste flavonoide au efecte anticancerigene asupra liniilor celulare de carcinom hepatocelular [ 176 , 177 , 178 ]. În plus, atunci când a fost testată pe o linie de celule hepatice normale (L02) la doze de până la 75 µM, eriocitrina a cauzat citotoxicitate scăzută [ 177 ]. Mai mult, așa cum arată Ferlazzo și colab., efectul anticancer al acestor flavonoide persistă chiar și atunci când compușii sunt administrați în totalitate. În cadrul studiului, tratamentul celulelor HepG2 cu suc de bergamotă 10% (care conține toate flavonoidele menționate mai sus) a scăzut rata de proliferare și a indus efect pro-apoptotic. Un total de 10% din sucul de bergamotă a indus oprirea ciclului celular în celulele HepG2 în faza G2. Această oprire a ciclului celular a coincis cu expresia crescută a proteinelor p53 și p21, ceea ce poate explica mecanismul prin care a avut loc oprirea ciclului celular. Expunerea la flavonoidele găsite în sucul de bergamotă a dus, de asemenea, la o expresie crescută a genelor legate de apoptoză, cum ar fi Bcl-2, caspaza-8, caspaza-9, caspaza-3, PARP, receptorul TNF (domeniul efector al morții), NF-kB , și IkB. În același timp, genele anti-apoptotice (cum ar fi BIRC8 și Bcl-2L2) s-au dovedit a fi reglate în jos. Translocarea nucleară NF-kB a fost, de asemenea, găsită scăzută. Prin urmare, flavonoidele din sucul de bergamotă pot activa atât căile apoptotice intrinseci mitocondriale, cât și cele extrinseci mediate de Fas [ 60 ].
Oroxylin A este un alt flavonoid, care s-a dovedit promițător ca o nouă opțiune potențială de tratament pentru HCC. Acest flavonoid este izolat din rădăcina unei plante asemănătoare cu menta numită Scutellariae radix și a fost folosit de mii de ani în medicina chineză pentru diferite afecțiuni de sănătate. Sa demonstrat că orxilina A la concentrații de 12,5, 25 și 50 pM reduce generarea de lactat și glucoză în celulele HepG2 în condiții hipoxice. De asemenea, a inhibat expresia HIF-1α, care este o proteină cheie de reglare a metabolismului energetic al celulelor tumorale în condiții hipoxice. Această proteină ajută la supraviețuirea celulelor tumorale în condiții hipoxice și glicolitice. Prin suprimarea HIF-1α, țintele sale din aval (PDK1, LDHA și HK II) au fost de asemenea inhibate [ 61 ]. Cu toate acestea, s-a constatat, de asemenea, că oroxilina A suprimă generarea de ATP în celulele hepatice normale (L02) atunci când este administrată în doze de până la 50 µM [ 61 ]. Alte studii au identificat că oroxilina A a crescut activarea HNF-4α în liniile celulare HepG2 și SMMC-7721. HNF-4α este un regulator în diferențierea hepatocitelor și un regulator pozitiv al HIF-1α [ 62 , 179 , 180 ]. Oroxylin A crește, de asemenea, raportul PKM1/PKM2, conducând la activarea HNF-4α și diferențierea ulterioară a celulelor HepG2 și în celulele SMMC-7721 la concentrații de 8 și respectiv 20 µM [62 ] . Diferențierea acestor celule împiedică proliferarea lor necontrolată și permite trecerea în celule funcționale nemaligne ale ficatului.
Quercetina este un flavonoid care se găsește în principal în fructe și legume, cum ar fi varză, fructe de pădure, mere, struguri roșii, broccoli, cireșe, ridichi, ceapă și în ceai și vin roșu [64 ] . Quercetina prezintă efecte de reglare asupra semnalizării protein kinazei C mediate de ROS internă și externă, prevenind deteriorarea oxidativă în organism prin activitatea antioxidantă și inducerea apoptozei în celulele canceroase [63 , 64 ] . În celulele HepG2, administrarea de quercetină duce la suprareglarea p53 și Bax [ 65 , 66 ]. Un alt studiu a constatat că quercetina a inhibat glicoliza în celulele Bel-7402 și SMMC-7721 la o doză de 50 µM prin reducerea nivelurilor de HK-2, care este o enzimă cheie implicată în glicoliză care este supraexprimată în HCC [67 , 181 , 182 ] . În plus, inhibarea glicolitică a fost, de asemenea, asociată cu o expresie redusă a mTOR și Akt fosforilate. Quercetina suprimă căile AKT/mTOR în celulele SMMC-7721 și Bel-7402 într-o manieră dependentă de doză (când este testată la concentrații de 12,5, 25 și 50 pM) prin reducerea nivelurilor de proteină HK-2. Tratamentul cu quercetină a inhibat, de asemenea, creșterea tumorilor xenogrefei HCC la șoarecii nuzi de sex feminin în vârstă de 5 săptămâni atunci când a fost administrat intraperitoneal la 50 mg/kg de două ori pe zi timp de 18 zile. Capacitatea quercetinei de a inhiba creșterea tumorilor s-a dovedit a fi datorită capacității sale de a reduce nivelurile de expresie a HK-2 [ 67 ].
Un flavonoid natural, fisetina, este prezent în multe fructe și legume, cum ar fi căpșuni, mere, castraveți, struguri, ceapă și curmali [ 183 ]. Fisetin și-a demonstrat capacitățile sale anti-oxidative, anti-metastatice și apoptotice pe diferite tipuri de cancer, inclusiv HCC [ 184 , 185 , 186 , 187 ]. Fisetin a redus în funcție de doză viabilitatea celulelor și clonogenitatea celulelor HepG2, a oprit ciclul celular în faza G2/M și a indus modificări morfologice, atunci când sunt administrate la 25, 50 și 100 pM. Fisetina a indus moartea celulară prin două mecanisme: apoptoză și necroptoză. Un număr crescut de celule apoptotice a fost identificat prin colorarea DAPI și vizualizarea cromatinei strâns condensate, ceea ce indică faptul că celulele intră într-o stare apoptotică. În plus, modificările morfologice, inclusiv condensarea nucleelor, sângerarea membranei, corpurile apoptotice și contracția celulară au confirmat moartea celulelor apoptotice în celulele HCC. Apoptoza a fost determinată și prin colorarea cu anexină V/PI, inițierea acesteia având loc prin exprimarea crescută a TNFα și expresia scăzută a NF-kB [ 68 ]. Fisetin a crescut expresia TNFα și IKκB și a scăzut expresia NF-kB, pNF-kB și pIKκB. Fisetin a redus, de asemenea, expresia Bcl2, în timp ce a crescut nivelurile de Bax, caspază-3 și PARP în celulele HepG2 [ 68 ]. Mai mult, fisetina a indus expresia crescută a RIPK1, pRIPK1, RIPK3, pRIPK3 și MLKL a confirmat că moartea celulelor a avut loc și prin necroptoză.
Glabridina este un tip de flavonoid clasificat în continuare ca izoflavonă. Acest compus este extras din rădăcinile plantei de lemn dulce și este folosit atât ca îndulcitor, cât și ca cosmetic natural. Pe lângă utilizările sale mai pietonale, acest compus posedă și caracteristici anti-cancer promițătoare. S-a demonstrat că glabridina inhibă atât căile de semnalizare TGF-p cât și SMAD2 în liniile celulare HepG2 și Huh-7 (la o doză de 20 pM). De asemenea, s-a dovedit că vizează SMAD2-3′ UTR, care a afectat expresia SMAD2 [ 69 ]. În plus, atunci când a fost administrat la celule hepatice normale (L02 la o doză de 10 și 20 µM) nu a fost observat niciun efect asupra viabilității celulelor [ 69 ]. Glabridina a scăzut semnificativ capacitatea invazivă a celulelor Huh-7 și SK-Hep-1 într-o manieră dependentă de doză (10, 20 și 40 pM). Acest efect s-ar putea datora, de asemenea, reglării în jos a proteinelor care facilitează migrarea, cum ar fi MMP-9 și MMP-1, din cauza unei suprareglări a inhibitorului tisular al MMP-urilor. Studiile in vivo au demonstrat suprimarea formării tumorii cu tratamentul cu glabridină într-un model de xenogrefă. S-a constatat că șoarecii masculi nuzi BALB/c în vârstă de 6 săptămâni xenogrefați SK-Hep-1 au redus semnificativ volumul tumorilor după ce au fost tratați cu glabridină intraperitoneal la 10 mg/kg timp de 28 de zile, în comparație cu șoarecii martor xenogrefați netratați. Dovezile din aceste studii sugerează o posibilă utilizare viitoare a glabridinei ca agent chimiopreventiv pentru metastaza HCC [ 70 ].
Genisteina, un izoflavonoid prezent în boabele de soia [ 71 ], s-a dovedit a avea efecte preventive asupra celulelor Bel 7402. Genisteina a inhibat creșterea celulelor tumorale și a indus oprirea ciclului celular în fazele G0/G1 și G2/M atunci când a fost tratată la o concentrație de 10 µg/mL [ 71 ]. S-a descoperit că șoarecii masculi BALB/c nuzi în vârstă de șase săptămâni, când au fost xenogrefați cu celule Bel 7402, au avut o invazie redusă după tratamentul intraperitoneal cu genisteina (50 mg/kg o dată pe zi timp de 15 zile), demonstrând că genisteina poate inhiba metastaza in vivo. [ 71 ]. În linia de celule de cancer hepatic Hep3B, s-a dovedit că genisteina (127,6 µM) funcționează în combinație cu gefitinib (9,8 µM), un inhibitor EGFR [ 72 ]. În plus, la șoarecii C57BL/6 care au suferit inducția chimică a HCC și care au fost hrăniți cu genisteina prin mâncare (80 mg/kg/zi timp de 5 luni), a existat o reducere remarcabilă a incidenței HCC în comparație cu șoarecii care au fost hrăniți doar cu mâncare obișnuită. De asemenea, s-a demonstrat că genisteina crește semnificativ fosfo-AMPK în extractele totale de țesut hepatic de la șoarecii tratați și în celulele Hep3B la concentrații de tratament de 1 și 5 pM. În plus, promovează, de asemenea, apoptoza și suprimă răspunsurile pro-inflamatorii, combătând afectarea ficatului [ 188 ].
Luteolina și kaempferolul sunt tetrahidroxiflavone naturale care se găsesc în multe fructe, legume și ierburi medicinale. Deși luteolina și kaempferolul sunt raportate ca molecule puternic antioxidante, s-a dovedit că au un efect citotoxic asupra carcinomului hepatocelular la concentrații de 12 mM pentru luteolină și 20 mM pentru kaempferol, prin generarea de ROS și eliberarea ulterioară de citocrom-c din mitocondriile [ 73 ]. Un alt studiu asupra luteolinei a arătat că aceasta a prezentat efect apro-apoptotic asupra liniilor celulare de HCC Huh7 și HepG2 (când a fost administrată la 10 pM). Studiile de xenogrefă HCC au arătat în continuare abilitățile anti-cancer ale luteolinei, atunci când șoarecii xenogrefe tratați cu 50 mg/kg/zi de luteolină au prezentat o dimensiune semnificativ scăzută a tumorii în comparație cu șoarecii de control tratați cu DMSO [74 ] .
Fagopyrum tataricum (L.) Gaertn (hrișcă tartară) este o plantă medicinală tradițională chinezească ale cărei rădăcini sunt bogate într-un compus flavonoid numit tatariside F (TF). TF este selectiv citotoxic pentru celulele de carcinom hepatocelular, așa cum s-a demonstrat atât în experimente in vitro, cât și in vivo. Când celulele H22 HCC au fost expuse la TF (la o concentrație de 10 mg/kg), p53 și Bax s-au dovedit a fi suprareglate dramatic, în timp ce Bcl-2 a fost reglată în jos. Acest efect a fost îmbunătățit atunci când celulele HCC au fost tratate atât cu TF, cât și cu ciclofosfamidă curentă pentru chimioterapie la 25 și, respectiv, 10 mg/kg. Când celulele HCC au fost tratate cu această combinație, s-a constatat că reglarea în sus a p53 și Bax a fost crescută semnificativ față de celulele tratate numai cu ciclofosfamidă. Celulele HCC injectate în șoareci ICR au fost lăsate să formeze tumori și apoi au fost tratate cu TF. Inhibarea semnificativă a greutății și creșterii tumorii a fost observată la șoarecii tratați cu 10 mg/kg de TF în comparație cu șoarecii martor. Acest efect inhibitor al TF a fost întărit atunci când a fost administrat împreună cu ciclofosfamidă, la doze de 25 și 10 mg/kg. Inhibarea creșterii tumorii prin combinarea TF și ciclofosfamidă a fost mai mare decât la șoarecii tratați numai cu ciclofosfamidă la 10 mg/kg. Aceste rezultate sugerează că TF poate avea potențial, nu numai ca terapie de sine stătătoare a cancerului, ci și ca un compus sinergic atunci când este utilizat în combinație cu medicamente anticancer existente pentru a le crește eficacitatea [75 ] .
Pulicaria jaubertii este o plantă comestibilă și aromatică originară din Europa, Asia și Africa, care s-a dovedit că reprimă dezvoltarea cancerului în celulele pulmonare umane și în celulele carcinomului hepatocelular uman, chiar și în comparație cu sorafenib (tratamentul actual de primă linie pentru HCC) [ 76 ]. Din patru glicozide diferite izolate din această plantă, doar una, Pulicaria jaubertii 1 (PJ-1), a arătat un potențial mare de a inhiba expresia mutantă a proteinei K-Ras/B-Raf prin inhibarea competitivă. Foarte important, s-a constatat că această inhibare apare numai în celulele canceroase HepG2 și nu în celulele hepatocitelor normale. Nu a fost observată nicio citotoxicitate atunci când hepatocitele normale au fost tratate cu doze de până la 1,5 mg/ml. Cu toate acestea, aceeași doză a fost eficientă în inducerea citotoxicității în celulele HepG2. În plus, PJ-1 a activat proteina p53 în timp ce a scăzut, de asemenea, nivelurile de expresie a TGF-beta și IL-8 [ 76 ].
Extractele de ceai verde din frunze de jujube (JLGTE) s-au dovedit, de asemenea, potențiale ca terapie împotriva cancerului. Deși compușii bioactivi majori din JLGTE nu au fost încă elucidați [ 189 ], există dovezi că componentele bioactive majore din frunzele de jujube sunt flavonoidele [ 189 ]. Cu toate acestea, JLGTE s-a dovedit a avea efecte anti-proliferative în celulele HepG2 la 10 µg/mL, în timp ce nu afectează linia celulară LO2 a hepatocitelor normale, chiar și atunci când este tratată la aceeași doză de 10 µg/mL. De asemenea, sa demonstrat că JLGTE exercită efecte pro-apoptotice prin activarea protein kinazei activate de AMP (AMPK) [ 77 ].
Icaritina este izolată din planta Epimedii herba , folosită în mod tradițional în remediile pe bază de plante chinezești [ 117 ]. Remediile care implică icaritina au fost utilizate de mult timp, deoarece pare să aibă un rol în modularea imunității. S-a demonstrat că tratamentele cu icaritină modulează celulele natural killer (NK), celulele T și celulele supresoare derivate din mieloid (MDSC) [ 190 ]. Studiile au demonstrat activități anti-proliferative ale icaritinei în celulele canceroase, precum și în celulele stem canceroase [ 191 ]. Cu toate acestea, s-a observat o activitate inhibitorie mică atunci când icaritina a fost administrată la o linie celulară hepatică normală (L02 la doze de până la 20 pM). Într-un studiu clinic, icaritina a arătat o tolerabilitate ridicată, fără evenimente adverse legate de imun la pacienții cu HCC avansat, atunci când a fost administrată la o doză de 600 mg oral de două ori pe zi, precum și la creșterea dozei la 800 mg [117 ] . Icaritina continuă să fie foarte promițătoare pentru viitoarele tratamente pentru HCC, deoarece este în prezent evaluată în două studii de fază III [ 192 ].
Catechine
Frunzele de ceai verde ( C. sinensis ) conțin, de asemenea, mulți compuși, care pot avea potențial ca tratamente pentru cancer. Deși extractul din frunze de ceai verde conține zeci de compuși organici (inclusiv cofeină și metilxantine), polifenolii găsiți în extractul de ceai verde au fost cel mai râvniți pentru natura lor antioxidantă și pentru numeroasele beneficii pentru sănătate. În plus, în ceaiurile verzi, toți acești compuși rămân relativ bine conservați, crescându-le potența și eficacitatea.
Epigallocatechin-3-galatul (EGCG) este unul dintre polifenolii extrași din ceaiul verde care prezintă caracteristici anti-cancer promițătoare. S-a demonstrat că EGCG poate împiedica proliferarea HCC în liniile celulare Hep3B, HepG2, SK-Hep1, HCC-LM3, Huh-7 și SMMC-7721 prin inhibarea ERα36, care este receptorul primar de estrogen pe celulele HCC [ 78 ] . EGCG a inhibat semnificativ proliferarea și a sporit efectele pro-apoptotice în celulele HCC care exprimă niveluri ridicate de ERα36 atunci când a fost tratată cu concentrații de 30 sau 60 ug/mL. În plus, celulele Hep3B cu expresie ridicată a ERα36 au arătat un efect inhibitor dependent de doză din aval datorită expunerii EGCG. Efectele din aval s-au dovedit a fi atât anti-proliferative (prin inhibarea căilor PI3K/Akt și MAPK/ERK), cât și pro-apoptotice (prin bucla de feedback ERα36-EGFR-Her-2 și activarea caspazei 3) [78 ] . Numeroase alte studii susțin capacitatea EGCG de a inhiba creșterea și metastaza HCC. S-a demonstrat că EGCG inhibă acumularea proteinei HIF1 α în celulele HepG2 tratate la 80 µg/mL prin modificarea căilor de semnalizare PI3K/Akt și ERK1/2 [ 79 , 80 ]. Mai mult, s-a raportat că EGCG a indus apoptoza în celulele HCC prin scăderea potențialului membranei mitocondriale și împiedicând proliferarea prin inducerea opririi ciclului celular G0/G1 [79 , 81 ] . De asemenea, s-a demonstrat că EGCG induce apoptoza în celulele Hep3B prin activarea caspazei-9 și a caspazei-3 atunci când este tratată la o doză de 10 pg/mL [79 , 82 ] . Anterior, o semnătură de 186 de gene a fost găsită în țesutul hepatic cirotic pentru a prezice riscul de HCC [ 193 ]. Folosind acești parametri, un studiu care a folosit dietilnitrozamină (DEN), un hepatocarcinogen, pentru a induce HCC la șobolani, a constatat că semnătura genei 186 „prognostic slab” este absentă din profilul de expresie a genei la șobolanii tratați cu EGCG [83 ] . Acesta este un indicator puternic că EGCG poate atenua dezvoltarea HCC. Studii suplimentare in vivo pot fi justificate pentru a susține efectele anticancer ale EGCG. Când șobolanii masculi Sprague Dawley HCC indus de DEN au fost tratați intragastric cu EGCG (la o doză de 25 mg/kg), volumul tumorii lor a scăzut semnificativ, iar ratele de supraviețuire au fost crescute [84 ] . Camellia sinensis (L.) O. Kuntze cv. CFT-1 este o varietate de ceai, care a fost special crescută pentru a avea un conținut ridicat de EGCG. Când șobolanii masculi Wistar cu HCC au fost expuși la infuzii de ceai de CFT-1 prin încorporare în apa de băut timp de 20 de săptămâni, nodulii hepatici au fost semnificativ redusi în dimensiune, număr și incidență în comparație cu șobolanii netratați cu CFT-1 [85] .]. De asemenea, s-a descoperit că EGCG modifică transcriptomul celulelor HCC. Când celulele Hep3B HCC au fost expuse la stimuli care imitau micromediul tumoral și apoi tratate cu EGCG, s-a constatat că expresiile genelor 922 au fost modificate în comparație cu celulele care nu au fost expuse la mimetismul micromediului tumoral [86 ] . Expresia miR483-3p s-a corelat cu scăderea supraviețuirii și creșterea progresiei HCC. Într-un model de șoarece HCC, s-a descoperit că tratamentul EGCG prin încorporarea a 0,1% și 0,5% EGCG în apa potabilă inversează îmbunătățirea indusă de mir483-3p a migrării și invaziei celulelor HCC. În plus, EGCG a putut împiedica modificarea indusă de mir483-3p a expresiei markerilor EMT: E-cadherină și vimentină. EGCG a redus, de asemenea, expresia endogenă a mir483-3p în celulele HCC [ 87 ]. EGCG s-a dovedit, de asemenea, promițător atunci când este utilizat împreună cu alte tratamente. În mod obișnuit, în multe cazuri de cancer, activarea receptorului de prostaglandină EP1 induce migrarea și invazia celulelor, conducând la apariția metastazelor. EGCG, atunci când este combinat cu antagonistul selectiv EP1 ONO-8711, a inhibat eficient viabilitatea celulelor HepG2 și capacitatea de migrare la o rată crescută decât EGCG singur (când este tratat la doze de 12,5, 25, 50 și 100 µg/mL) [88 ] . Aceste constatări aruncă lumină asupra mecanismelor specifice prin care EGCG suprimă creșterea HCC.
3.2.2. Stilbene
Pterostilbenul, un stilbenoid găsit în mod natural înrudit cu resveratrolul ca analog metoxilat, se găsește în mod obișnuit în afine, mai multe tipuri de struguri și lemn de copac. Este considerabil mai biodisponibil datorită celor două grupe metoxi ale sale în comparație cu resveratrolul, ceea ce face ca acesta să devină un compus de interes pentru tratamentul cancerului [ 194 ]. Pterostilbenul are un efect inhibitor asupra aproape tuturor proceselor celulare care duce la metastazare într-o manieră dependentă de apoptoză și independentă de apoptoză. Prezintă o capacitate anti-metastatică prin prevenirea colonizării celulelor tumorale și scăderea tumorilor secundare în organe îndepărtate, care a fost găsită atunci când modelul de șoareci nuzi cu cancer pancreatic au fost tratați cu 100 µg/kg/zi și 500 µg/kg/zi de pterostilben [ 89 ] . Inhibarea creșterii HCC a fost demonstrată atunci când linia de celule canceroase SMMC-7721 a fost tratată cu 50 uM de pterostilben. Inhibarea pare să fie realizată prin reglarea în jos a complexului de proteină 1 asociată cu metastaza (MTA1) și histon deacetilază 1 (HDC1). Destabilizarea acestui complex crește nivelurile de activare a supresoarelor tumorale PTEN în tumori, care în cele din urmă promovează apoptoza în HCC [ 90 ]. Eforturile viitoare pot transforma pterostilbenul într-un medicament complementar pentru a reduce rata metastazelor în HCC.
Resveratrolul este un produs natural care se găsește în mod obișnuit în struguri, arahide și pini cu efecte anti-oxidative, antiinflamatorii și citoprotectoare cunoscute [ 91 ]. Oligomerii de resveratrol s-au dovedit a fi citotoxici pentru celulele HepG2 la 21 µM prin inducerea opririi ciclului celular la G2/M, creșterea ROS intracelular și creșterea activității caspazei-3 [92 ] . În plus, s-a dovedit că resveratrolul inhibă proliferarea, migrarea și invazia HCC prin reglarea în jos a degetului 1 de tip CH inelar asociat membranei (MARCH1), o ubiquitin ligază E3 care se găsește în mod obișnuit a fi puternic reglată în HCC [93 ] . Tratamentul cu resveratrol, atunci când a fost administrat pe cale orală la șobolani cu HCC în doză de 100 mg/kg timp de 2 săptămâni, a restabilit enzimele antioxidante catalaza și glutation peroxidaza la niveluri normale, deoarece acestea sunt de obicei reglate în jos în HCC [94 ] . S-a demonstrat că o dietă cu resveratrol hrănită la șobolani Sprague Dawley (la o doză de 300 mg/kg) reduce producția de oxid azotic în ficat prin reglarea inductibilă a sintetazei de oxid nitric [95 ] . Hexokinaza 2 (HK2) este o enzimă cunoscută că modifică fenotipul metabolic al celulelor pentru a susține creșterea anaerobă. Resveratrolul administrat celulelor HCC-LM3 la o concentrație de 20 pM a inhibat expresia HK2 [ 96 ]. Combinat cu sorafenib, resveratrolul inhibă creșterea celulelor HCC și crește efectul anticancer al medicamentului prin suprimarea crescută a creșterii anaerobe [ 96 ]. În modelele de șobolan HCC induse de DEN, resveratrolul s-a dovedit a fi un tratament eficient împotriva cancerului, indicat de o creștere semnificativă a apoptozei [ 97 ]. Când resveratrolul a fost administrat unui model de șoarece HCC, s-a constatat că creșterea tumorii a fost inhibată printr-o scădere marcată a frecvenței CD8+CD122+Treg în tumori, ganglioni limfatici și splina șoarecelui. Celulele T CD8+ care exprimă IFN-γ în tumori și organele limfoide periferice s-au dovedit, de asemenea, crescute [ 32 ]. Efectele resveratrolului par să fie îmbunătățite atunci când compusul poate fi livrat mai precis la locul cancerului. În plus, efectele anticancer ale resveratrolului au fost îmbunătățite atunci când sunt livrate precis la HCC prin sistemul de livrare a nanoparticulelor decât atunci când sunt administrate în mod tradițional [ 98 ]. Un alt studiu a constatat, de asemenea, un succes similar cu un sistem de livrare nano folosind lipozomi cationici pentru a furniza resveratrol; a redus semnificativ numărul de noduli hepatici [ 99 ].
3.2.3. Xantone
Garcinia mangostana, cunoscută și sub numele de mangostan, este un fruct tropical popular cultivat în țările din Asia de Sud-Est, cunoscut pentru gustul său dulce. S-a raportat că doi compuși unici izolați din G. mangostana au efecte anti-cancer. Sunt două xantone: mangostanaxantona V și mangostanaxantona VI [ 100 ]. S-a descoperit că ambele xantone inhibă potențialul proliferativ al celulelor HepG2 în mod semnificativ. Mai mult, mangostanaxantona V a oprit celulele în faza G0/G1 a ciclului celular, în timp ce mangostanaxantona VI s-a dovedit a avea un impact asupra fazelor G2/M [ 100 ].
Alfa-mangostinul este un alt compus activ de xantonă din Garcinia mangostana . Când a fost administrat la o doză de 5,5 pM, a sensibilizat celulele HepG2 la anoikis [ 101 ], un tip de moarte celulară programată care apare atunci când celulele dependente de ancorare se desprind de matricea lor extracelulară. Acest proces este vital pentru răspândirea metastatică a tumorilor maligne [ 195 ]. S-a descoperit că alfa-mangostinul a sensibilizat foarte mult celulele HepG2 la anoikis prin stimularea mecanismelor pro-apoptotice, cum ar fi inducerea caspazei-9, caspazei-8 și caspazei-3 și a scăzut nivelurile de proteine anti-apoptotice. Alfa-mangostinul a redus semnificativ readerența și migrarea celulelor HepG2 (când a fost administrată la o doză de 2 pM) prin inhibarea MMP-2 și MMP-9, împreună cu suprimarea moleculelor implicate în căile de semnalizare AKT și ERK. Aceste descoperiri sugerează că alfa-mangostinul poate fi un medicament multi-țintă eficient pentru tratarea HCC [ 101 ]. Interesant este că diferitele doze de alfa-mangostin (10, 20, 30, 40 și 50 µM) testate pe celule osteogene umane normale nu au arătat nicio modificare a viabilității celulare, ceea ce indică faptul că alfa-mangostină a avut doar citotoxicitate minimă pentru celulele normale [196] . ].
Acidul Gambogic (GA) este o xantonă izolată din gamboge, o rășină uscată din arborii din genul Garcinia . În mod tradițional, gamboge a fost folosit ca medicament antiinflamator, paraziticid și de detoxifiere de mii de ani în toată Asia de Sud-Est [ 197 ]. Sa demonstrat că acidul Gambogic are un efect anticancerigen asupra celulelor SMMC-7721 prin necroză și proliferări reduse atunci când este administrat la o concentrație de 0,625–5,0 mg/mL și un efect inhibitor crescut asupra telomerazei atunci când este administrat la o doză de 2 mg/mL. [ 104 ]. Într-un alt studiu, GA a indus apoptoza prin stres oxidativ în celulele HepG2 atunci când a fost administrat la o concentrație de 0,75–12 µM [ 105 ]. Celulele maligne supraexprimă izoenzimele tioredoxin reductază (TrxR) pentru creșterea tumorii lor. TrxR funcțional joacă, de asemenea, un rol important în scăderea sarcinii de stres oxidativ în celule. GA inhibă activitatea TrxR1 pentru a induce stresul oxidativ și apoi apoptoza în celulele SMMC-7721 HCC atunci când este administrată la o doză de 3 pM [ 106 ].
3.3. Alcaloizi
Alcaloizii sunt compuși care conțin azot care se găsesc din abundență în multe plante. Alcaloizii au fost folosiți în scopuri terapeutice de mulți ani și sunt bine cunoscuți pentru proprietățile lor curative și medicinale [ 198 ]. Alcaloizii (cum ar fi cofeina) pot fi găsiți din abundență în cafea, al cărei consum este asociat cu un risc scăzut de HCC, precum și cu un risc scăzut de boli hepatice cronice. Un studiu de cohortă prospectiv bazat pe populație pe mai mult de 215.000 de bărbați și femei a constatat că, indiferent de etnie, sex, IMC, statutul de fumat, consumul de alcool sau starea de diabet, participanții care au consumat cafea au avut o incidență comparativ mai mică a carcinomului hepatocelular și a bolii hepatice cronice. [ 199 ]. Într-un studiu cu o perioadă de urmărire de 18 ani, în comparație cu cei care nu consumau cafea, cei care consumau două până la trei căni pe zi au arătat o reducere cu 38% a riscului de a dezvolta HCC. S-a constatat că cei care au băut patru sau mai multe căni pe zi în timpul aceleiași perioade de urmărire au o reducere cu 41% a riscului de CHC [ 199 ]. Într-o meta-analiză, persoanele care au băut cafea au avut mai puține șanse de a dezvolta ciroză sau fibroză hepatică în comparație cu cei care nu consumau cafea. [ 200 ]. Fibroza și ciroza cresc riscul de HCC; prin urmare, căile care conduc la dezvoltarea acestor boli ar putea fi ținte potențiale pentru tratamentul HCC. Efectele antioxidante ale cafelei în ficat sunt asociate cu Nrf-2, un factor de transcripție care se leagă de elementul de răspuns antioxidant (ARE) pentru a promova multe gene antioxidante. Proteinele și enzimele cunoscute a fi reglate de Nrf-2 au arătat o creștere a expresiei pe măsură ce concentrația de cafea a crescut, rezultând o reducere generală a stresului oxidativ [ 201 ]. Interesant, nu s-a descoperit că există un efect de prevenire a chimio-chimioterapiei HCC la persoanele care au băut cafea decofeinizată în comparație cu cei care nu consumă cafea, sugerând că cofeina poate juca un rol important în capacitatea de prevenire a chimio-chimiografică a cafelei [202 , 203 ] . În plus, studii mai specifice privind tratamentul HCC au arătat că terapia combinată de cofeină cu 5-fluorouracil, în comparație cu terapia cu numai 5-fluorouracil, a demonstrat scăderea proliferării celulare și o creștere semnificativă a apoptozei prin reglarea producției intracelulare de ROS [ 41 ] .
Rhizoma coptidis, cunoscut și sub denumirea de fir de aur chinezesc, este o plantă cu flori originară din China ale cărei rădăcini au fost folosite pe scară largă în China de mii de ani pentru a trata multe afecțiuni și boli diferite [204 ] . Recent, au apărut dovezi care sugerează că planta ar putea avea potențial ca o opțiune terapeutică viitoare pentru tratamentul HCC. Berberina, (un alcaloid derivat din izochinolină), este compusul principal găsit în Rhizoma coptidis . Berberina provoacă apoptoză și are efecte anti-proliferative dependente de doză (10, 50 și 100 pM) asupra celulelor HepG2 [ 102 ]. În plus, celulele HepG2 tratate cu berberină 50 µM au avut niveluri reduse de NF-kB-p65, sugerând că berberina poate acționa prin p65 pentru a suprima calea NF-kB [ 102 ].
Neferina este un alcaloid natural de bisbenzilizochinolin derivat din embrionii de semințe verzi ale plantei acvatice înflorite, lotusul indian ( Nelumbo nucifera ). Neferina a indus apoptoza în celulele Hep3B la doze de peste 10 pM prin reglarea în jos a genelor proliferative c-Myc, ciclina D1, D3, CDK4 și E2F-1 și inducerea stresului ER [103 ] . Yoon și colab., au demonstrat că neferina a prezentat, de asemenea, citotoxicitate selectivă și apoptoză față de celulele HCC și nu în celulele hepatice normale THLE-3. Cu celulele hepatice normale, a existat o pierdere minimă a viabilității celulelor chiar și după tratamentul cu 30 pM de neferină.
Planta alcaloid irinotecan provine din planta Camptotheca acuminata . Irinotecanul este un inhibitor al topoizomerazei-1 care interferează cu sinteza ADN-ului provocând rupturi de ADN monocatenar și inhibând enzimele de reparare a ADN-ului [ 205 ]. Potențialul terapeutic al compusului irinotecan a fost demonstrat și este aprobat de FDA pentru cancerul colorectal metastatic [ 205 ]. Într-un studiu clinic, pacienții cu HCC au fost tratați cu irinotecan în doză de 125 mg/m 2 . Deși pacienții au avut un rezultat modest de îmbunătățire după tratament, toxicitatea cu acest compus este o preocupare reală.
3.4. Polizaharide
Polizaharidele sunt lanțuri lungi compuse din zaharuri simple. Ele sunt imperative pentru viață și se găsesc într-o mare varietate de plante, animale și ciuperci.
Compusul activ corelat cu hexoză (AHCC) este un extract polizaharidic de Basidiomycotina derivat din hibridizarea ciupercilor [ 206 ]. S-a demonstrat că AHCC joacă un rol în funcția celulelor imune și în numărul de celule; astfel de celule imunitare includ celule natural killer (NK) și celule T, care poate apăra gazda de infecții și afecțiuni maligne [ 207 ]. Când a fost administrat cu o doză mare de 9 g de AHCC lichid pe zi, nu au fost raportate anomalii semnificative ale parametrilor de laborator la subiecții sănătoși [ 208 ]. Depășirea nivelurilor ridicate de toxicitate de la majoritatea compușilor introduși într-un cadru clinic este întotdeauna un obstacol, dar, așa cum s-a afirmat în studiul anterior, AHCC are o toxicitate potențial scăzută la oameni, prin urmare, făcând o opțiune viabilă ca agent terapeutic. Adăugarea de AHCC la 360 mg·kg -1 ·d -1 cu o doză mică de chimioterapie cu 5-fluorouracil la 10 mg·kg -1 ·d -1 tratament la șoareci a dus la un efect antitumoral mai mare și a provocat mai multă apoptoză în tumoră țesut [ 209 ].
3.5. Extracte întregi
Extractele întregi derivate din plante pot conține mai mult de un fitochimic și sunt foarte promițătoare ca terapii viitoare pentru cancer. Poncirus fructus (PF) este un extract de metanol din fructele uscate imature de Poncirus trifoliata . Acest arbust de foioase crește în mod natural în insulele sud-coreene Jeju și Gaduk, iar culturile din Asia de Est și-au folosit de mult fructele pentru a trata tulburări variind de la gastrointestinale la cardiovasculare [107 ] . Pe lângă faptul că este benefic pentru problemele de sănătate menționate mai sus, PF pare să conțină și proprietăți anti-cancer promițătoare. PF a demonstrat citotoxicitate împotriva celulelor HCC, inducând apoptoza și inhibând semnificativ abilitățile proliferative și de formare de colonii ale Hep3B și Huh7 într-o manieră dependentă de doză atunci când este tratată la doze de 20, 30 și 40 µM in vitro [107 ] . PF pare să induce citotoxicitate în celulele HCC prin calea de apoptoză mediată mitocondrial; PF a indus pierderea potențialului membranei mitocondriale, care, la rândul său, a provocat niveluri ridicate de generare intracelulară de ROS. În plus, PF a inhibat capacitățile migratoare și invazive ale celulelor HCC prin creșterea expresiei markerului epitelial E-cadherină, în timp ce a redus simultan markerii mezenchimali și ai ciclului celular B-catenină, N-cadherină, vimentină, Snail, a-SMA, Pin1 și ciclină. D1 [ 107 ]. Mai mult, PF a redus MMP-2 și MMP-9. Aceste MMP sunt jucători cruciali în capacitatea celulelor tumorale de a invada, deoarece ajută la degradarea membranei bazale. O astfel de atenuare a activității MMP-2 și MMP-9 sugerează că PF poate fi un compus util în prevenirea răspândirii HCC.
Trametes robiniophila, cunoscut și sub numele de Huaier, este o ciupercă care a fost folosită ca medicament tradițional chinezesc de peste 1000 de ani [ 210 ]. Cele mai comune preparate de Huaier includ granule sau extracte apoase. Principalul ingredient activ, proteoglicanul, constă din aminoacizi, polizaharide și apă [ 211 ]. Huaier are efecte anticancer puternice asupra HCC [ 212 ], cancerului de sân [ 213 ] și multe altele. Când au fost administrate celulelor HepG2 HCC la o doză de 15 mg/mL, granulele acestei ciuperci au scăzut proliferarea și migrarea [ 214 ]. În plus, granula Huaier nu a provocat citotoxicitate în celulele hepatocitelor umane normale L-02, indicând că vizează numai celulele HCC [ 214 ]. Într-un studiu clinic controlat randomizat, ingestia unui extract apos din Trametes robiniophila (20 g în 100 ml apă, de 3 ori pe zi timp de 96 de săptămâni) a scăzut semnificativ ratele de recidivă și metastaze ale cancerului, în același timp îmbunătățind ratele de supraviețuire la pacienții care au suferit rezecție hepatică. pentru HCC [ 215 ].
Smallanthus sonchifolius leaf (YLE ) sau Yacón este o specie de margaretă perenă, care a fost cultivată în mod tradițional în nordul și centrul Anzilor din Columbia până în nordul Argentinei pentru rădăcinile sale cu gust dulce. La începutul anilor 2000, YLE a fost cultivat în Myanmar și de atunci a fost folosit ca remediu tradițional pe bază de plante în zonele rurale din Myanmar pentru a trata bolile hepatice. Recent, s-a demonstrat că extractul YLE metanolic de plantă integrală posedă activitate anticanceroasă semnificativă împotriva celulelor HepG2 (cu un IC50 de 58,2 pg/mL). Când a fost administrat celulelor HepG2, YLE a indus semnificativ necroza fără a activa caspazele 3 sau 8 și a indus oprirea ciclului celular în linia celulară. YLE a scăzut, de asemenea, ROS atât în mod dependent de doză, cât și de timp, dând sprijin efectelor sale inhibitoare asupra proliferării celulelor canceroase [ 216 ]. În mod interesant, viabilitatea celulelor HEK 293 non-tumorale nu a fost afectată de YLE (0-100 µg/mL), indicând citotoxicitatea este specifică celulelor HCC [ 216 ].
Cyperus amuricus, cunoscut și sub denumirea de piatră asiatică, este o iarbă originară din China, Japonia, Coreea și Rusia europeană, care a fost folosită în medicina populară orientală ca astringent, pentru vindecarea rănilor, ca diuretic și pentru alte probleme intestinale [217] . ]. Extractul metanolic din plante întregi de C. amuricus a demonstrat efecte citotoxice împotriva celulelor Hep3B la 100–200 µg/mL și a sugerat că moartea celulară indusă de extractul de C. amuricus a avut loc printr-un mecanism apoptotic, bazat pe modificările morfologice asociate cu tratament 24 de ore. Tratamentul celulelor Hep3B a dus la inhibarea CDK-urilor și a ciclinelor, ducând la oprirea ciclului celular la sub-G1. Datele sugerează, de asemenea, că scindarea FADD a procaspazei-8 în caspaza-8 activată poate fi declanșată de C. amuricus . Apoi, scindarea ulterioară reglată de caspaza-8 a lui Bid în tBid și translocarea tBid în mitocondrii poate permite diafonia între căile apoptotice intrinseci și extrinseci. Mai mult, activarea ordonată a caspazelor clivate-8, -9, -3, -7, -6 și a PARP clivate împreună cu alte proteine legate de apoptoză oferă un sprijin suplimentar ideii că proprietățile anticanceroase ale C. amuricus sunt legate de capacitatea sa . pentru a induce apoptoza [ 217 ]. Important, C. amuricus (50–200 µg/mL) nu a avut efecte citotoxice semnificative asupra cheratinocitelor umane necanceroase [ 217 ].
Fraxini-2 provine din planta de vâsc Viscum album L., derivată din frasin, care conține multe substanțe biologic active [ 114 ]. S-a demonstrat că Fraxini-2 are activitate naturală a celulelor ucigașe ca urmare a producției de interferon gamma (INF-γ), a factorului de necroză tumorală-α (TNF-α), a activării monocitelor-macrofage și a celulelor helper și a nivelurilor mai ridicate. de citokine [ 114 ]. Într-un studiu clinic, 12 pacienți cu HCC avansat au primit fraxini-2 prin injecție subcutanată (la o doză de 20 mg o dată pe săptămână timp de două săptămâni) [218 ] . Pacienții au urmat acest regim de tratament timp de diferite perioade, variind de la 2,6 la 36,4 săptămâni. În general, nivelurile de alfa-fetoproteină (AFP) ale pacienților par să se stabilizeze în primele 3-4 săptămâni de tratament cu fraxini-2. Nivelurile ridicate de AFP sunt folosite istoric ca markeri serici pentru prezența HCC [ 219 ], ca atare, nivelurile scăzute de AFP observate după tratamentul cu fraxini-2 sunt rezultate încurajatoare. Dovezile din acest studiu par să indice că ar putea exista un loc pentru AFP ca opțiune de tratament suplimentar în viitor; cu toate acestea, trebuie făcute mai multe studii. Populația totală a acestui studiu a fost foarte mică, rezultatele pentru doar 10 persoane fiind disponibile la sfârșit. În plus, AFP se poate dovedi benefică numai pentru pacienții cu HCC specifici, așa că ar fi necesare studii viitoare care să controleze tratamentele anterioare și caracteristicile bolii pentru a putea trage concluzii convingătoare.
3.6. Combinaţie
Substanțele fitochimice pot câștiga eficacitate datorită efectelor lor sinergice anticancer atunci când sunt combinate cu alți compuși sau fitochimice. Potențialul pe care îl pot avea aceste combinații ca terapii anti-cancer viitoare este promițător și justifică o explorare suplimentară.
Mai mult, acetatul de 1′-acetoxichavicol (ACA) este un ester de acetat derivat din rizomii de Alpinia galangal din familia ghimbirului. Este folosit în mod obișnuit ca medicament tradițional și ca condiment în artele culinare. ACA a indus apoptoza în celulele HepG2 atunci când a fost administrat la 10 -5 M [ 220 ]. ACA s-a dovedit a fi benefic în prevenirea progresului carcinogenezei pe tumorile induse chimic, atunci când este administrat la celulele epiteliale intestinale de șobolan [ 221 ]. De asemenea, s-a descoperit că ACA este un compus anti-cancer benefic atunci când este utilizat împreună cu butiratul de sodiu. Butiratul de sodiu este un acid gras cu lanț scurt produs în principal prin fermentarea bacteriană a fibrelor alimentare despre care s-a demonstrat anterior că are efecte anticancerigene [ 222 , 223 , 224 , 225 ]. Tratamentul combinat cu ACA și butiratul de sodiu a determinat o creștere semnificativă a speciilor reactive de oxigen și a activităților NADPH oxidazei în celulele HepG2 [ 108 ]. Acest tratament combinat a arătat, de asemenea, un efect inhibitor sinergic asupra creșterii HCC, care a fost cel mai puternic atunci când 12,5 pM de ACA a fost combinat cu butirat de sodiu 2 mM. Mai mult, a crescut semnificativ nivelurile de AMPK. Tratamentul combinat ACA plus butirat de sodiu ar putea fi utilizat ca terapie țintită pentru HCC fără a provoca toxicitate hepatică semnificativă [ 108 ]. S-a constatat că indicele de combinație este mai mic de 1, indicând în plus o relație sinergică puternică între ACA și butiratul de sodiu.
S-a demonstrat că acidul betulinic (Bet A) și ginsenozida Rh2 (G-Rh2) induc apoptoza în celulele HepG2. Bet A, un triterpenoid pentaciclic, este derivat din mesteacănii albi. Individual, promovează moartea celulelor pro-apoptotice în diferite tipuri de celule prin calea mitocondrială [ 226 , 227 , 228 ]. G-Rh2 este izolat din rădăcina de ginseng Panax [ 229 , 230 ]. Ginseng a fost folosit în mod tradițional în medicina pe bază de plante în Asia de Est, ingredientul său activ principal fiind ginsenozide. Ginsenozidele au prezentat efecte asupra răspunsului imun, metabolismului și tratamentului și prevenirii cancerului [ 231 , 232 ]. G-Rh2 singur a indus moderat apoptoza în celulele HepG2; totuși, împreună cu Bet A, a avut efecte mai pronunțate asupra proliferării și apoptozei HCC, cel mai puternic la doze de 10 µg/mL de Bet A combinate cu 12,5 µg/mL de G-Rh2. Astfel de procese au fost evidențiate de modificări morfologice caracteristice celulelor apoptotice, inclusiv fragmentare nucleară, blebbing membranar și condensare nucleară [ 109 ]. Indicele de combinație al acestor compuși a fost mai mic de 1, indicând un efect sinergic.
Decoctul Huanglian este compus din patru medicamente tradiționale chinezești pe bază de plante: Coptidis Rhizoma , Zingiberis Rhizoma , Folium Artemisiae Argyi și Mume Fructus . Compusul activ al acestui amestec este alcaloidul clorhidrat de berberină (0,26 mg/mL). Decoctul Huanglian are un efect negativ direct asupra creșterii liniilor celulare de cancer hepatic. Capacitatea lor de migrare și invazie este împiedicată prin inducerea apoptozei și oprirea ciclului celular în faza G2/M. În plus, reglarea în jos a ciclinei-B1 G2/mitotic-specific (CCNB1) activează calea p53 prin reglarea în sus a lui Bax, caspaza-3, caspaza-9, p21 și p53 [110 ] . Decoctul Huanglian arată în continuare efectele sale remarcabile in vivo [ 111 ], prin care creșterea tumorii și angiogeneza într-un model murin xenogrefat s-au dovedit a fi suprimate după tratamentul cu decoct. Se presupune că mecanismul de acțiune este prin atenuarea eEF-2 [ 112 ], care este o enzimă care se dovedește a fi suprareglată în diferite tipuri de cancer [ 233 ].
Medicina tradițională chineză (TCM) include, de asemenea, compuși, cum ar fi Fufang Banmao Capsule și Huaier Granule [ 234 ]. Fufang Banmao are o istorie lungă de utilizare pentru tratarea HCC. Într-un studiu, 320 de pacienți cu HCC au primit Fufang Banmao într-o formă de 0,25 g/capsulă, care urmează să fie administrat oral de două ori pe zi timp de 6 luni. În comparație cu grupul de control din studiu care nu a luat capsula, pacienții care au luat Fufang Banmao au arătat o creștere semnificativă a ratelor de supraviețuire generale [ 235 ]. Huaier Granule este derivat în mod specific dintr-un amestec de produse de fermentație care provin din planta Trametes robiniophila Murr [ 234 ]. Această granulă a arătat, de asemenea, dovezi că are efecte anticancer împotriva HCC, în special prin promovarea apoptozei prin p38-MAPK, inhibarea metastazei și angiogeneza și, de asemenea, provocând oprirea ciclului celular în faza G0/G1 [234 ] . Într-un studiu clinic pe 340 de persoane, s-a descoperit că granula Huaier prelungește supraviețuirea fără progresie a pacienților cu HCC care au suferit anterior ablație termică. Acei pacienți care au luat granula Huaier mai mult de 2 ani prin administrare orală au văzut cele mai multe beneficii [ 116 ]. S-a demonstrat că acești compuși reduc riscul de HCC la pacienții cu hepatită B și reduc riscul de ciroză [ 236 , 237 ]. Studiile au arătat potențialul unor astfel de substanțe fitochimice de a inhiba creșterea celulelor HCC, de a induce moartea celulelor și de a reduce răspunsul inflamator, precum și de a crește activitatea antivirală.
Terapia Jianpi Huayu este un alt MTC care constă din ginseng, atractylodes, tuckahoe, rădăcină de lemn dulce, radix bupleuri , yam, cortex moutan , salvia miltiorrhiza , turmeric și rhizoma zedoariae [ 120 ]. TCM s-a dovedit a fi abundent în efecte terapeutice, inclusiv reducerea toxicității radiațiilor/chimioterapiei, îmbunătățirea calității vieții pacientului și prelungirea supraviețuirii pacientului [ 234 , 238 , 239 ] și Jianpi Huayu pare să nu facă excepție. Terapia Jianpi Huayu a fost raportată a fi benefică pentru pacienții cu HCC într-un studiu clinic atunci când este administrată post-hepatectomie într-un studiu clinic [ 120 ]. În plus, un studiu a încercat să exploreze mecanismul de bază al terapiei Jianpi Huayu și eficacitatea acestuia împotriva HCC. Celulele SMMC-7721 au fost implantate ortotopic în șoareci nuzi în vârstă de 4 până la 7 săptămâni. Odată ce tumorile au fost prezente, șoarecii au primit 24,96 g/kg de compus Jianpi Huayu intragastric timp de 21 de zile, în timp ce șoarecii de control au primit ser fiziologic. Până la sfârșitul experimentului, șoarecii tratați cu Jianpi Huayu au redus semnificativ dimensiunea și greutatea tumorii în comparație cu șoarecii martor hrăniți cu soluție salină. Studiul speculează că suprimarea HCC se poate datora capacității lui Jianpi Huayu de a regla în jos nivelurile miR-602, care sunt de obicei suprareglate la pacienții cu cancer hepatic [ 240 ]. Deși acest studiu a avut rezultate promițătoare, utilizarea unei doze atât de mari de Jianpi Huayu este puțin îngrijorătoare când vine vorba de traducere la ființe umane. Sunt necesare cercetări suplimentare pentru a determina toxicitatea acestui compus la om, pentru a stabili un nivel de dozare maxim sigur. Cu toate acestea, este, de asemenea, important de reținut că nu au existat cazuri de toxicitate în studiul clinic [ 120 ].
Decoctul Jianpi Ligan este alcătuit din mai multe componente TCM care implică Radix Codonopsis , Rhizoma Atractylodis macrocephala , Poria cocos , Radix Glycyrrhizae , Rhizoma Diosscoreae , Rhizoma Pinelliae , Fructus Crataegi , Semen Nelumbinis , Periacarus , Herba Arteca , 1 și Permiscar scoppi . În două studii clinice retrospective, decoctul Jianpi Ligan, administrat împreună cu chemoembolizarea arterială transcateter (TACE) sau cu ablația prin radiofrecvență (RFA), a arătat o supraviețuire globală semnificativ îmbunătățită în comparație cu pacienții care au primit numai tratamente TACE sau RFA [ 121 , 122 ] .
Amestecul Shenqi este un tip de amestec TCM de multe ierburi constând din rădăcină de pseudostarwort, rădăcină de rădăcină de lapte, poria, tubercul alb de atractylodes, rădăcină de rehmannia, dendrobium, rădăcină de bujor alb, rădăcină de angelica chineză, plantă Oldenlandia, lobelia chineză, ghimbir sălbatic și rădăcină de lemn dulce. [ 129 ]. Medicamentul în amestec Shenqi are beneficii precum îmbunătățirea circulației sângelui și eliminarea toxinelor [ 129 ]. Această combinație de plante s-a dovedit a avea succes într-un studiu clinic în care 20 ml (conținând 20 g de amestec brut) au fost administrați oral pacienților (care au primit anterior coagulare la microunde) de trei ori pe zi. Rata de eficacitate a tratamentului în grupul de tratament a fost de 75,00% comparativ cu grupul de control.
Carcinomul hepatocelular este o boală mortală în creștere rapidă, care are puține opțiuni terapeutice disponibile în prezent pacienților. HCC devine și mai letal odată cu producerea metastazelor și, pentru a înrăutăți lucrurile, opțiunile de tratament în acest stadiu se scad și mai mult. Există o cerere mare de a explora rezervoare naturale neexploatate pentru a produce medicamente și opțiuni de tratament pentru pacienții care prea des nu au niciunul. Din fericire, mulți compuși fitochimici arată promițător ca opțiuni terapeutice pentru pacienții cu metastază HCC. În plus, substanțele fitochimice au și un potențial mare ca terapii adjuvante. Mai multe studii au arătat siguranța, eficacitatea, fezabilitatea și mecanismul de acțiuni ale fitochimicelor care susțin încorporarea lor în chimioterapiile curente.
Cu toate acestea, este important să luăm în considerare deficiențele și limitările care vin împreună cu multe dintre aceste procese de investigație. Adesea, studiile in vitro se pot dovedi a fi mai înșelătoare decât lămuritoare. Experimentele de cultură celulară nu pot replica complexitățile unui întreg organism biologic și, din acest motiv, există posibilitatea ca, deși se vede că un compus are un efect in vitro, poate exista cu totul alt efect observat în modelele in vivo. Cu toate acestea, în cele mai grave cazuri, poate exista de fapt o toxicitate severă în organism, chiar dacă nu a fost observată o astfel de toxicitate în liniile celulare. În ceea ce privește testarea in vivo, este important să rețineți că multe dintre aceste cercetări folosesc rozătoarele ca modele biologice pentru a explora efectele potențiale ale noilor compuși, deoarece aceștia sunt ușor de manevrat și accesibili. Acest lucru în sine are limitările sale, deoarece rozătoarele au metabolism și caracteristici diferite față de o ființă umană. Încă o dată, chiar dacă un compus arată promițător in vitro și in vivo, este posibil să nu poată fi transpus eficient la oameni.
Există, de asemenea, obstacolul puterii statistice și al mărimii populației. Pentru multe dintre studiile clinice discutate aici, numărul de pacienți testați a fost destul de mic. Populațiile mai mici limitează puterea studiului clinic, deoarece rezultatele (chiar dacă sunt foarte pozitive) nu pot fi aplicate în siguranță unei populații mari, pe baza rezultatelor și a efectelor secundare experimentate doar de un număr mic de oameni. Studiile clinice mici sunt ele însele fundamente pentru studii clinice mai mari, prin care mai mulți oameni implicați pot ajuta cu adevărat oamenii de știință și medicii să ajungă la o concluzie mai puternică din punct de vedere statistic.
Dozarea devine o problemă serioasă și atunci când se ia în considerare translabilitatea multora dintre acești compuși. Deși pot exista efecte pozitive observate în studiile preclinice, efectele se pot datora probabil distribuirii unor doze mari pe celule sau animale, care nu ar putea fi translabile la om. Încă o dată, toxicologia devine o preocupare reală atunci când se încearcă aducerea de noi tratamente și medicamente în clinici din studii preclinice. Cel mai bine ar fi să folosiți cea mai mică doză cu beneficii maxime, dar dozele eficiente utilizate în diferite studii diferă drastic. Este bine cunoscut faptul că niciun produs fitochimic nu este la fel de activ. Eficacitatea și potența fitochimicelor depind de câțiva parametri diferiți, cum ar fi biodisponibilitatea medicamentului, absorbția, clearance-ul etc. Unele dintre ele ar putea fi necesare în cantități mari pentru a produce un efect benefic, în timp ce altele ar putea fi necesare doar în cantități foarte mici. Unele substanțe fitochimice sunt eficiente în doze mari și asta s-ar putea datora faptului că ingredientul activ ar putea fi în concentrații scăzute. Prin urmare, se așteaptă ca dozele eficiente să varieze de la fitochimic la fitochimic.
În plus, există o lipsă de tehnici de standardizare când vine vorba de acești compuși. Există, de exemplu, metode diferite de extracție, întreținere, depozitare și administrare a acestor substanțe fitochimice. Toate aceste variații pot avea un efect profund asupra activității compușilor și pot explica de ce un laborator de cercetare poate vedea rezultate semnificative și altul nu. Până nu există o practică mai standardizată de extracție și manipulare pentru fiecare compus, rezultatele pot fi foarte variabile și, prin urmare, nesigure. În plus, din cauza lipsei de standardizare, unii compuși pot conține, din neatenție, toxine, care ar putea denatura rezultatele prin cauzarea unor daune care nu sunt rezultatul compusului în sine. Este esențial să se acorde atenție calității compusului fitochimic care este testat. Din aceste motive, rămân multe obstacole între experimentarea de bază și aplicarea clinică reală.
Oricum, procesele științifice pe care le folosim pentru a face noi descoperiri sunt adesea suficient de eficiente, așa cum demonstrează progresele zilnice în știință și medicină, folosind aceleași abordări ca cele discutate în studiile analizate aici. Deși sunt necesare mult mai multe informații pentru multe dintre aceste substanțe fitochimice înainte ca acestea să devină tratamente clinice cu drepturi depline, fundația pentru această ramură a medicinei pare foarte puternică.
În această revizuire, am discutat efectele fitochimice asupra HCC din studiile preclinice și clinice și am găsit rezultate încurajatoare și promițătoare. Substanțele fitochimice raportate în această revizuire au fost clasificate în mai multe categorii, cum ar fi lipide, polifenoli, alcaloizi, polizaharide, extracte întregi și combinații fitochimice (Figura 3). Susținem cu tărie că este nevoie de mai multă atenție științifică asupra fitochimicelor pentru a continua evaluarea efectelor lor benefice și combaterea HCC. De o importanță deosebită este necesitatea de a extinde domeniile de biodisponibilitate, farmacocinetică și farmacodinamică a multor dintre acești compuși. Mai multe investigații asupra modului în care fitochimicele pot fi utilizate ca medicamente eficiente împotriva cancerului ar putea fi cheia pentru a reduce decalajul dintre studiile preclinice și studiile clinice pentru multe dintre aceste fitochimice. Aproape 80% dintre compuși nu au reușit să progreseze în studiile clinice din cauza lipsei de informații privind eficacitatea, biodisponibilitatea și toxicitatea neprevăzută pentru celulele normale [ 241 ]. Deși rămân obstacole mari, substanțele fitochimice pot fi utilizate fie ca terapie alternativă, fie ca terapie integrativă împreună cu chimioterapiile HCC existente. Utilizarea modelelor predictive non/preclinice și a tehnologiei organ-on-a chip care seamănă mai precis cu fiziologia umană este crucială pentru a evalua farmacocinetica, farmacodinamia, eficacitatea și siguranța fitochimicelor terapeutice. În concluzie, fitochimicele au un mare potențial ca opțiuni de tratament pentru carcinomul hepatocelular.
Dorim să mulțumim Departamentului de Medicină Moleculară și Translațională, Texas Tech University Health Sciences Center El Paso pentru sprijinul acordat acestei cercetări.
Conceptualizare, RS; anchetă, SR, KS, TM-G., MG, SK; scriere—pregătirea proiectului original, SR, KS, TM-G., MG, SK și RS; scriere — revizuire și editare, SR, KS, TM-G., MG, SK, RS, JR, AY, MS și SP; vizualizare, SR, MG, SK și RS, supraveghere, RS; administrarea proiectului, RS Toți autorii au citit și au fost de acord cu versiunea publicată a manuscrisului.
Sung H., Ferlay J., Siegel RL, Laversanne M., Soerjomataram I., Jemal A., Bray F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021; 71 :209–249. doi: 10.3322/caac.21660. [ PubMed ] [ CrossRef ] [ Google Scholar ]2.
Sagnelli E., Macera M., Russo A., Coppola N., Sagnelli C. Variații epidemiologice și etiologice în carcinomul hepatocelular. Infecţie. 2020; 48 :7–17. doi: 10.1007/s15010-019-01345-y. [ PubMed ] [ CrossRef ] [ Google Scholar ]3.
Bray F., Ferlay J., Soerjomataram I., Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimations of incident and mortality worldwide for 36 cancers in 185 states. CA Cancer J. Clin. 2018; 68 :394–424. doi: 10.3322/caac.21492. [ PubMed ] [ CrossRef ] [ Google Scholar ]4.
Sarveazad A., Agah S., Babahajian A., Amini N., Bahardoust M. Predictorii ratei de supraviețuire la 5 ani la pacienții cu carcinom hepatocelular. J. Res. Med. Sci. 2019; 24:86 . doi: 10.4103/jrms.JRMS_1017_18. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]5.
Sun W., Fu S., Wu S., Tu R. Dovezi în creștere ale metastazei legate de microARN exosomal ale carcinomului hepatocelular. Biomed Res. Int. 2020; 2020 :4501454. doi: 10.1155/2020/4501454. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]6.
Keating GM Sorafenib: O revizuire în carcinomul hepatocelular. Țintă Oncol. 2017; 12 :243–253. doi: 10.1007/s11523-017-0484-7. [ PubMed ] [ CrossRef ] [ Google Scholar ]7.
de Martel C., Maucort-Boulch D., Plummer M., Franceschi S. Contribuția relativă la nivel mondial a virusurilor hepatitei B și C în carcinomul hepatocelular. hepatologie. 2015; 62 :1190–1200. doi: 10.1002/hep.27969. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]8.
Chen DS Virusul hepatitei C în boala hepatică cronică și carcinomul hepatocelular în Taiwan. Printesa Takamatsu Symp. 1995; 25 :27–32. [ PubMed ] [ Google Scholar ]9.
Nordenstedt H., White DL, El-Serag HB Modelul în schimbare al epidemiologiei în carcinomul hepatocelular. Săpa. Ficat Dis. 2010; 42 ((Supliment 3)):S206–S214. doi: 10.1016/S1590-8658(10)60507-5. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]11.
Gentile D., Donadon M., Lleo A., Aghemo A., Roncalli M., di Tommaso L., Torzilli G. Tratamentul chirurgical al hepatocolangiocarcinomului: o revizuire sistematică. Cancer de ficat. 2020; 9 :15–27. doi: 10.1159/000503719. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]12.
Wong TC, Lo CM Strategii de rezecție pentru carcinomul hepatocelular. Semin Ficat Dis. 2013; 33 :273–281. doi: 10.1055/s-0033-1351782. [ PubMed ] [ CrossRef ] [ Google Scholar ]13.
Lau WY, Leung TW, Lai BS, Liew CT, Ho SK, Yu SC, Tang AM Chimioimunoterapie sistemică preoperatorie și rezecție secvențială pentru carcinom hepatocelular nerezecabil. Ann. Surg. 2001; 233 :236–241. doi: 10.1097/00000658-200102000-00013. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]14.
Mazzaferro V., Regalia E., Doci R., Andreola S., Pulvirenti A., Bozzetti F., Montalto F., Ammatuna M., Morabito A., Gennari L. Transplant de ficat pentru tratamentul carcinoamelor hepatocelulare mici la pacientii cu ciroza. N. Engl. J. Med. 1996; 334 :693–699. doi: 10.1056/NEJM199603143341104. [ PubMed ] [ CrossRef ] [ Google Scholar ]15.
Spallanzani A., Orsi G., Andrikou K., Gelsomino F., Rimini M., Riggi L., Cascinu S. Lenvatinib ca terapie pentru carcinomul hepatocelular nerezecabil. Expert Rev. Anticancer Ther. 2018; 18 :1069–1076. doi: 10.1080/14737140.2018.1524297. [ PubMed ] [ CrossRef ] [ Google Scholar ]16.
Llovet JM, Ricci S., Mazzaferro V., Hilgard P., Gane E., Blanc JF, de Oliveira AC, Santoro A., Raoul JL, Forner A., et al. Sorafenib în carcinomul hepatocelular avansat. N. Engl. J. Med. 2008; 359 :378–390. doi: 10.1056/NEJMoa0708857. [ PubMed ] [ CrossRef ] [ Google Scholar ]17.
Rimassa L., Pressiani T., Merle P. Systemic Treatment Options in Hepatocellular Carcinoma. Cancer de ficat. 2019; 8 :427–446. doi: 10.1159/000499765. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]18.
Zheng A., Chevalier N., Calderoni M., Dubuis G., Dormond O., Ziros PG, Sykiotis GP, Widmann C. Screeningul la nivel de genom CRISPR/Cas9 identifică KEAP1 ca genă de sensibilitate la sorafenib, lenvatinib și regorafenib în carcinomul hepatocelular. Oncotarget. 2019; 10 :7058–7070. doi: 10.18632/oncotarget.27361. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]19.
Ao J., Chiba T., Shibata S., Kurosugi A., Qiang N., Ma Y., Kan M., Iwanaga T., Sakuma T., Kanzaki H., et al. Achiziția de fenotipuri asemănătoare mezenchimatoase și supraproducția de factori angiogenici în celulele de carcinom hepatocelular rezistent la lenvatinib. Biochim. Biophys. Res. comun. 2021; 549 :171–178. doi: 10.1016/j.bbrc.2021.02.097. [ PubMed ] [ CrossRef ] [ Google Scholar ]20.
Tang W., Chen Z., Zhang W., Cheng Y., Zhang B., Wu F., Wang Q., Wang S., Rong D., Reiter FP și colab. Mecanismele rezistenței la sorafenib în carcinomul hepatocelular: baze teoretice și aspecte terapeutice. Semnal. Transduct. Ţintă. Acolo. 2020; 5:87 . doi: 10.1038/s41392-020-0187-x. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]21.
Raoul JL, Gilabert M., Oziel-Taïeb S., Giovannini M. Tratament paliativ pentru carcinomul hepatocelular. Pr. Prat. 2013; 63 :233–236. [ PubMed ] [ Google Scholar ]22.
Habib A., Desai K., Hickey R., Thornburg B., Lewandowski R., Salem R. Terapia locoregională a carcinomului hepatocelular. Clin. Ficat Dis. 2015; 19 :401–420. doi: 10.1016/j.cld.2015.01.008. [ PubMed ] [ CrossRef ] [ Google Scholar ]23.
Sangro B., Iñarrairaegui M., Bilbao JI Radioembolization for hepatocellular carcinom. J. Hepatol. 2012; 56 :464–473. doi: 10.1016/j.jhep.2011.07.012. [ PubMed ] [ CrossRef ] [ Google Scholar ]24.
Helgason CM Comentariu asupra semnificației pentru neurologia modernă a Papirusului chirurgical al secolului al XVII-lea î.Hr. Poate sa. J. Neurol. Sci. 1987; 14 :560–563. [ PubMed ] [ Google Scholar ]25.
Cragg GM, Grothaus PG, Newman DJ Impactul produselor naturale asupra dezvoltării de noi agenți anti-cancer. Chim. Rev. 2009; 109 :3012–3043. doi: 10.1021/cr900019j. [ PubMed ] [ CrossRef ] [ Google Scholar ]26.
Zhou DD, Luo M., Shang A., Mao QQ, Li BY, Gan RY, Li HB Componente alimentare antioxidante pentru prevenirea și tratarea bolilor cardiovasculare: efecte, mecanisme și studii clinice. Oxid. Med. Cell Longev. 2021; 2021 :6627355. doi: 10.1155/2021/6627355. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]27.
Graf BL, Raskin I., Cefalu WT, Ribnicky DM Terapeutice derivate din plante pentru tratamentul sindromului metabolic. Curr. Opinează. Investig. Droguri. 2010; 11 :1107–1115. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]28.
Sharifi-Rad M., Lankatillake C., Dias DA, Docea AO, Mahomoodally MF, Lobine D., Chazot PL, Kurt B., Tumer TB, Moreira AC, et al. Impactul compușilor naturali asupra tulburărilor neurodegenerative: de la preclinic la farmacoterapie. J. Clin. Med. 2020; 9 :1061. doi: 10.3390/jcm9041061. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]29.
Zhou T., Ye L., Bai Y., Sun A., Cox B., Liu D., Li Y., Liotta D., Snyder JP, Fu H. și colab. Autofagie și apoptoză în carcinomul hepatocelular indus de EF25-(GSH)2: Un nou analog de curcumină. Plus unu. 2014; 9 :e107876. doi: 10.1371/journal.pone.0107876. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]30.
Zhao Z., Malhotra A., Seng WY Curcumina modulează carcinomul hepatocelular prin reducerea expresiei UNC119. J. Environ. Pathol. Toxicol. Oncol. 2019; 38 :195–203. doi: 10.1615/JEnvironPatholToxicolOncol.2019029549. [ PubMed ] [ CrossRef ] [ Google Scholar ]31.
Zheng M., Chen R., Zhong H., Lin Q., Wang X., Zhao Z., Xie L. Efecte secundare ale resveratrolului în celulele HepG2: Pten redus și expresie crescută a mRNA bcl-xl. Mol. Med. Rep. 2012; 6 :1367–1370. doi: 10.3892/mmr.2012.1077. [ PubMed ] [ CrossRef ] [ Google Scholar ]32.
Zhang Q., Huang H., Zheng F., Liu H., Qiu F., Chen Y., Liang CL, Dai Z. Resveratrolul exercită efecte antitumorale prin scăderea CD8. Oncoimunologie. 2020; 9 :1829346. doi: 10.1080/2162402X.2020.1829346. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]33.
Wu L., Li J., Liu T., Li S., Feng J., Yu Q., Zhang J., Chen J., Zhou Y., Ji J. și colab. Quercetina prezintă efect anti-tumoral în celulele carcinomului hepatocelular LM3 prin abrogarea căii de semnalizare JAK2/STAT3. Cancer Med. 2019; 8 :4806–4820. doi: 10.1002/cam4.2388. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]34.
Zhang S., Yang Y., Liang Z., Duan W., Yang J., Yan J., Wang N., Feng W., Ding M., Nie Y., și colab. Inhibarea mediată de silibină a semnalizării Notch exercită activitate antitumorală în celulele carcinomului hepatocelular uman. Plus unu. 2013; 8 :e83699. doi: 10.1371/journal.pone.0083699. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]35.
Rawat D., Shrivastava S., Naik RA, Chhonker SK, Mehrotra A., Koiri RK O prezentare generală a produselor naturale din plante în tratamentul carcinomului hepatocelular. Agenti anticancerigen Med. Chim. 2018; 18 :1838–1859. doi: 10.2174/1871520618666180604085612. [ PubMed ] [ CrossRef ] [ Google Scholar ]36.
Watanabe S., Kitade Y., Masaki T., Nishioka M., Satoh K., Nishino H. Efectele licopenului și Sho-saiko-to asupra hepatocarcinogenezei într-un model de șobolan de cancer hepatic spontan. Nutr. Cancer. 2001; 39 :96–101. doi: 10.1207/S15327914nc391_13. [ PubMed ] [ CrossRef ] [ Google Scholar ]37.
Thomas CE, Luu HN, Wang R., Adams-Haduch J., Jin A., Koh WP, Yuan JM Asociația dintre consumul alimentar de roșii și riscul de carcinom hepatocelular: Studiul de sănătate chinezesc din Singapore. Epidemiol de cancer. Biomark. Prev. 2020; 29 :1430–1435. doi: 10.1158/1055-9965.EPI-20-0051. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]38.
Xia H., Liu C., Li CC, Fu M., Takahashi S., Hu KQ, Aizawa K., Hiroyuki S., Wu G., Zhao L. și colab. Pulberea dietetică de roșii inhibă carcinomul hepatocelular promovat de dieta cu conținut ridicat de grăsimi cu modificarea microbiotei intestinale la șoarecii care nu au enzime de scindare carotenoide. Cancer Prev. Res. 2018; 11 :797–810. doi: 10.1158/1940-6207.CAPR-18-0188. [ PubMed ] [ CrossRef ] [ Google Scholar ]39.
Zhou RS, Wang XW, Sun QF, Ye ZJ, Liu JW, Zhou DH, Tang Y. Efectele anticanceroase ale Emodinului asupra celulei HepG2: Dovezi din analiza bioinformatică. Biomed Res. Int. 2019; 2019 :3065818. doi: 10.1155/2019/3065818. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]40.
Subramaniam A., Shanmugam MK, Ong TH, Li F., Perumal E., Chen L., Vali S., Abbasi T., Kapoor S., Ahn KS, et al. Emodina inhibă creșterea și induce apoptoza într-un model de carcinom hepatocelular ortotopic prin blocarea activării STAT3. Br. J. Pharmacol. 2013; 170 :807–821. doi: 10.1111/bph.12302. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]41.
Wang Z., Gu C., Wang X., Lang Y., Wu Y., Wu X., Zhu X., Wang K., Yang H. Cofeina sporește efectul antitumoral al 5-fluorouracilului prin creșterea producerea de specii reactive de oxigen în carcinomul hepatocelular. Med. Oncol. 2019; 36:97 . doi: 10.1007/s12032-019-1323-8. [ PubMed ] [ CrossRef ] [ Google Scholar ]42.
Saraswati S., Alhaider A., Abdelgadir AM, Tanwer P., Korashy HM Phloretin atenuează activitatea STAT-3 și depășește rezistența la sorafenib, vizând inhibarea mediată de SHP-1 a căii STAT3 și Akt/VEGFR2 în carcinomul hepatocelular. Celulă comun. Semnal. 2019; 17 :127. doi: 10.1186/s12964-019-0430-7. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]43.
Luo H., Wang YJ, Chen J., Liu JQ Studiu asupra efectului floretinei asupra inhibării feotipului malign al celulelor BEL-7402. Zhong Yao Cai. 2008; 31 :1019–1021. [ PubMed ] [ Google Scholar ]44.
Newman DJ, Cragg GM Produsele naturale ca surse de medicamente noi în ultimii 25 de ani. J. Nat. Prod. 2007; 70 :461–477. doi: 10.1021/np068054v. [ PubMed ] [ CrossRef ] [ Google Scholar ]45.
Karimi A., Majlesi M., Rafieian-Kopaei M. Herbal versus synthetic drugs; credințe și fapte. J. Nefrofarmacol. 2015; 4 :27–30. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]46.
Mondal A., Guria T., Maity TK, Bishayee A. Un nou acid gras tetraenoic izolat din Amaranthus spinosus inhibă proliferarea și induce apoptoza celulelor canceroase ale ficatului uman. Int. J. Mol. Sci. 2016; 17 :1604. doi: 10.3390/ijms17101604. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]47.
Dasgupta A., Dey D., Ghosh D., Lai TK, Bhuvanesh N., Dolui S., Velayutham R., Acharya K. Astrakurkurone, un sesquiterpenoid din ciuperca comestibilă sălbatică, vizează celulele canceroase hepatice prin modularea Bcl-2 proteinele familiei. Viața IUBMB. 2019; 71 :992–1002. doi: 10.1002/iub.2047. [ PubMed ] [ CrossRef ] [ Google Scholar ]48.
Kim DK, Baek JH, Kang CM, Yoo MA, Sung JW, Chung HY, Kim ND, Choi YH, Lee SH, Kim KW Activitatea apoptotică a acidului ursolic se poate corela cu inhibarea inițierii replicării ADN-ului. Int. J. Cancer. 2000; 87 :629–636. doi: 10.1002/1097-0215(20000901)87:5<629::AID-IJC2>3.0.CO;2-P. [ PubMed ] [ CrossRef ] [ Google Scholar ]49.
Kim GH, Kan SY, Kang H., Lee S., Ko HM, Kim JH, Lim JH Acidul ursolic suprimă biosinteza colesterolului și exercită efecte anti-cancer în celulele de carcinom hepatocelular. Int. J. Mol. Sci. 2019; 20 :4767. doi: 10.3390/ijms20194767. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]50.
Shyu MH, Kao TC, Yen GC Acidul oleanolic și acidul ursolic induc apoptoza în celulele de carcinom hepatocelular uman HuH7 printr-o cale dependentă de mitocondrie și reglarea în jos a XIAP. J. Agric. Food Chim. 2010; 58 :6110–6118. doi: 10.1021/jf100574j. [ PubMed ] [ CrossRef ] [ Google Scholar ]51.
Yang L., Zhang XY, Li K., Li AP, Yang WD, Yang R., Wang P., Zhao ZH, Cui F., Qin Y. și colab. Protopanaxadiolul inhibă tranziția epitelial-mezenchimală a carcinomului hepatocelular prin țintirea căii STAT3. Moartea celulară Dis. 2019; 10 :630. doi: 10.1038/s41419-019-1733-8. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]52.
Lou L., Ye W., Chen Y., Wu S., Jin L., He J., Tao X., Zhu J., Chen X., Deng A. și colab. Ardipusilozida inhibă supraviețuirea, invazia și metastaza celulelor de carcinom hepatocelular uman. Fitomedicina. 2012; 19 :603–608. doi: 10.1016/j.phymed.2012.01.003. [ PubMed ] [ CrossRef ] [ Google Scholar ]53.
Durmaz I., Guven EB, Ersahin T., Ozturk M., Calis I., Cetin-Atalay R. Celulele canceroase hepatice sunt sensibile la moartea celulară indusă de Lanatoside C, independent de statutul lor PTEN. Fitomedicina. 2016; 23 :42–51. doi: 10.1016/j.phymed.2015.11.012. [ PubMed ] [ CrossRef ] [ Google Scholar ]54.
Ko H., Huh G., Jung SH, Kwon H., Jeon Y., Park YN, Kim YJ Diospyros kaki frunzele inhibă EMT mediată de semnalizare HGF/Met și caracteristicile stemness în carcinomul hepatocelular. Food Chim. Toxicol. 2020; 142 :111475. doi: 10.1016/j.fct.2020.111475. [ PubMed ] [ CrossRef ] [ Google Scholar ]55.
Liao W., Fan L., Zheng Z., Liu H., Deng H., Li M., Liu F., Yang A. Ziyuglycoside II exercită efecte antiproliferative și antimetastazice asupra celulelor carcinomului hepatocelular. Medicamente anticancerigene. 2020; 31 :819–827. doi: 10.1097/CAD.0000000000000918. [ PubMed ] [ CrossRef ] [ Google Scholar ]56.
Wang T., Dou Y., Lin G., Li Q., Nie J., Chen B., Xie J., Su Z., Zeng H., Chen J., și colab. Efectul anti-carcinom hepatocelular al uleiului de Brucea javanica la șoarecii purtători de tumori ascitice: detectarea brusatolului și rolul său. Biomed Pharma. 2021; 134 :111122. doi: 10.1016/j.biopha.2020.111122. [ PubMed ] [ CrossRef ] [ Google Scholar ]57.
Jung KH, Rumman M., Yan H., Cheon MJ, Choi JG, Jin X., Park S., Oh MS, Hong SS O fracțiune de acetat de etil a Artemisia capillaris (ACE-63) a indus apoptoza și anti-angiogeneza prin inhibarea semnalizării PI3K/AKT în carcinomul hepatocelular. Phytother. Res. 2018; 32 :2034–2046. doi: 10.1002/ptr.6135. [ PubMed ] [ CrossRef ] [ Google Scholar ]58.
Weifeng T., Feng S., Xiangji L., Changqing S., Zhiquan Q., Huazhong Z., Peining Y., Yong Y., Mengchao W., Xiaoqing J. și colab. Artemisinina inhibă invazia și metastaza in vitro și in vivo a celulelor de carcinom hepatocelular uman. Fitomedicina. 2011; 18 :158–162. doi: 10.1016/j.phymed.2010.07.003. [ PubMed ] [ CrossRef ] [ Google Scholar ]59.
Delle Monache S., Sanità P., Trapasso E., Ursino MR, Dugo P., Russo M., Ferlazzo N., Calapai G., Angelucci A., Navarra M. Mecanisms underlying anti-tumoral effects of Citrus Suc de Bergamia. Plus unu. 2013; 8 :e61484. doi: 10.1371/journal.pone.0061484. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]60.
Ferlazzo N., Cirmi S., Russo M., Trapasso E., Ursino MR, Lombardo GE, Gangemi S., Calapai G., Navarra M. NF-κB mediază efectele antiproliferative și proapoptotice ale sucului de bergamotă în celulele HepG2 . Life Sci. 2016; 146 :81–91. doi: 10.1016/j.lfs.2015.12.040. [ PubMed ] [ CrossRef ] [ Google Scholar ]61.
Dai Q., Yin Q., Wei L., Zhou Y., Qiao C., Guo Y., Wang X., Ma S., Lu N. Oroxylin A reglează metabolismul glucozei ca răspuns la stresul hipoxic cu implicarea de factor-1 inductibil de hipoxie în celulele HepG2 de hepatom uman. Mol. Carcinog. 2016; 55 :1275–1289. doi: 10.1002/mc.22369. [ PubMed ] [ CrossRef ] [ Google Scholar ]62.
Wei L., Dai Y., Zhou Y., He Z., Yao J., Zhao L., Guo Q., Yang L. Oroxylin A activează PKM1/HNF4 alfa pentru a induce diferențierea hepatomului și a bloca progresia cancerului. Moartea celulară Dis. 2017; 8 :e2944. doi: 10.1038/cddis.2017.335. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]63.
Anand David AV, Arulmoli R., Parasuraman S. Overviews of Biological Importance of Quercetin: A Bioactive Flavonoid. Pharmacogn. Rev. 2016; 10 :84–89. doi: 10.4103/0973-7847.194044. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]64.
Xu D., Hu MJ, Wang YQ, Cui YL Activitățile antioxidante ale quercetinei și complexele sale pentru aplicare medicinală. Molecule. 2019; 24 :1123. doi: 10.3390/molecules24061123. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]65.
Maurya AK, Vinayak M. Acțiunea anticancerigenă a quercetinei prin reglarea în jos a fosfatidilinozitol 3-kinazei (PI3K) și a proteinei kinazei C (PKC) prin inducerea p53 în linia celulară de carcinom hepatocelular (HepG2). Mol. Biol. Rep. 2015; 42 :1419–1429. doi: 10.1007/s11033-015-3921-7. [ PubMed ] [ CrossRef ] [ Google Scholar ]66.
Maurya AK, Vinayak M. Modularea semnalizării PKC și inducerea apoptozei prin suprimarea speciilor reactive de oxigen și a receptorului 1 al factorului de necroză tumorală (TNFR1): Rolul cheie al quercetinei în prevenirea cancerului. Tumor Biol. 2015; 36 :8913–8924. doi: 10.1007/s13277-015-3634-5. [ PubMed ] [ CrossRef ] [ Google Scholar ]67.
Wu H., Pan L., Gao C., Xu H., Li Y., Zhang L., Ma L., Meng L., Sun X., Qin H. Quercetina inhibă proliferarea HCC dependent de glicoliză Celule prin reducerea Hexokinazei 2 și a căii Akt-mTOR. Molecule. 2019; 24 :1993. doi: 10.3390/molecules24101993. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]68.
Sundarraj K., Raghunath A., Panneerselvam L., Perumal E. Fisetin, un fitopolifenol, vizează moartea celulelor apoptotice și necroptotice în celulele HepG2. Biofactori. 2020; 46 :118–135. doi: 10.1002/biof.1577. [ PubMed ] [ CrossRef ] [ Google Scholar ]69.
Jiang F., Mu J., Wang X., Ye X., Si L., Ning S., Li Z., Li Y. Efectul represiv al miR-148a asupra căii semnalului TGF beta-SMAD este implicat în inhibarea indusă de glabridină a proprietăților asemănătoare celulelor stem canceroase în celulele de carcinom hepatocelular. Plus unu. 2014; 9 :e96698. doi: 10.1371/journal.pone.0096698. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]70.
Hsieh MJ, Lin CW, Yang SF, Chen MK, Chiou HL Glabridin inhibă migrarea și invazia prin inhibarea transcripțională a metaloproteinazei matriceale 9 prin modularea activității NF-kB și AP-1 în celulele canceroase hepatice umane. Br. J. Pharmacol. 2014; 171 :3037–3050. doi: 10.1111/bph.12626. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]71.
Gu Y., Zhu CF, Iwamoto H., Chen JS Genisteina inhibă potențialul invaziv al carcinomului hepatocelular uman prin modificarea ciclului celular, apoptoza și angiogeneza. Lumea J. Gastroenterol. 2005; 11 :6512–6517. doi: 10.3748/wjg.v11.i41.6512. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]72.
Tong Y., Wang M., Huang H., Zhang J., Huang Y., Chen Y., Pan H. Efectele inhibitoare ale genisteinei în combinație cu gefitinib asupra liniei celulare de carcinom hepatocelular Hep3B. Exp. Acolo. Med. 2019; 18 :3793–3800. doi: 10.3892/etm.2019.8027. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]73.
Seydi E., Salimi A., Rasekh HR, Mohsenifar Z., Pourahmad J. Selective Cytotoxicity of Luteolin and Kaempferol on Cancerous Hepatocytes Obtained from Rat Model of Hepatocellular Carcinoma: Implication of ROS-Mediated Mitochondrial Targeting. Nutr. Cancer. 2018; 70 :594–604. doi: 10.1080/01635581.2018.1460679. [ PubMed ] [ CrossRef ] [ Google Scholar ]74.
Yang PW, Lu ZY, Pan Q., Chen TT, Feng XJ, Wang SM, Pan YC, Zhu MH, Zhang SH MicroRNA-6809-5p mediază efectele anticancer induse de luteolină împotriva hepatomului prin țintirea flotilinei 1. Fitomedicină . 2019; 57 :18–29. doi: 10.1016/j.phymed.2018.10.027. [ PubMed ] [ CrossRef ] [ Google Scholar ]75.
Peng W., Hu C., Shu Z., Han T., Qin L., Zheng C. Activitatea antitumorală a tatarizidei F izolate din rădăcinile Fagopyrum tataricum (L.) Gaertn împotriva carcinomului hepatocelular H22 prin reglarea în sus a p53. Fitomedicina. 2015; 22 :730–736. doi: 10.1016/j.phymed.2015.05.003. [ PubMed ] [ CrossRef ] [ Google Scholar ]76.
Abd El Maksoud AI, Taher RF, Gaara AH, Abdelrazik E., Keshk OS, Elawdan KA, Morsy SE, Salah A., Khalil H. Selective Regulation of B-Raf Dependent K-Ras/Mitogen-Activated Protein by Natural Inhibitori multi-kinaze care apar în celulele canceroase. Față. Oncol. 2019; 9 :1220. doi: 10.3389/fonc.2019.01220. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]77.
Liu HX, Xu MQ, Li SP, Tian S., Guo MX, Qi JY, He CJ, Zhao XS Extractele de ceai verde din frunze de jujube inhibă celulele carcinomului hepatocelular prin activarea AMPK. Oncotarget. 2017; 8 :110566–110575. doi: 10.18632/oncotarget.22821. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]78.
Chen J., Chen L., Lu T., Xie Y., Li C., Jia Z., Cao J. ERα36 este o țintă eficientă a epigalocatechin-3-galat în carcinomul hepatocelular. Int. J. Clin. Exp. Pathol. 2019; 12 :3222–3234. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]79.
Bimonte S., Albino V., Piccirillo M., Nasto A., Molino C., Palaia R., Cascella M. Epigallocatechin-3-gallate in the prevention and treatment of hepatocellular carcinom: Experimental findings and translational perspectives. Drug Des. Devel. Acolo. 2019; 13 :611–621. doi: 10.2147/DDDT.S180079. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]80.
Zhang Q., Tang X., Lu Q., Zhang Z., Rao J., Le AD Extractul de ceai verde și (-)-epigallocatechin-3-galatul inhibă hipoxia și acumularea de proteine HIF-1alfa induse de ser și Expresia VEGF în celulele de carcinom cervical uman și hepatom. Mol. Cancer Ther. 2006; 5 :1227–1238. doi: 10.1158/1535-7163.MCT-05-0490. [ PubMed ] [ CrossRef ] [ Google Scholar ]81.
Zhang Y., Owusu L., Duan W., Jiang T., Zang S., Ahmed A., Xin Y. Efecte antimetastatice și diferențiale asupra expresiei proteice a epigallocatechin-3-galat în celulele carcinomului hepatocelular HCCLM6. Int. J. Mol. Med. 2013; 32 :959–964. doi: 10.3892/ijmm.2013.1446. [ PubMed ] [ CrossRef ] [ Google Scholar ]82.
Chen L., Ye HL, Zhang G., Yao WM, Chen XZ, Zhang FC, Liang G. Inhibarea autofagiei contribuie la interacțiunea sinergică dintre EGCG și doxorubicină pentru a ucide celulele Hep3B de hepatom. Plus unu. 2014; 9 :e85771. doi: 10.1371/journal.pone.0085771. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]83.
Sojoodi M., Wei L., Erstad DJ, Yamada S., Fujii T., Hirschfield H., Kim RS, Lauwers GY, Lanuti M., Hoshida Y., et al. Galatul de epigalocatechină induce senescența celulelor stelate hepatice și atenuează dezvoltarea carcinomului hepatocelular. Cancer Prev. Res. 2020; 13 :497–508. doi: 10.1158/1940-6207.CAPR-19-0383. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]84.
Tang Y., Cao J., Cai Z., An H., Li Y., Peng Y., Chen N., Luo A., Tao H., Li K. Galatul de epigalocatechină induce efecte chimiopreventive asupra șobolanilor cu dietilnitrozamină -cancer hepatic indus prin inhibarea ciclului de diviziune celulară 25A. Mol. Med. Rep. 2020; 22 :3873–3885. doi: 10.3892/mmr.2020.11463. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]85.
Liao S., Lin J., Liu J., Chen T., Xu M., Zheng J. Chemoprevention of elite tea soi CFT-1 bogat în EGCG împotriva cancerului hepatic indus chimic la șobolani. Științe alimentare Nutr. 2019; 7 :2647–2665. doi: 10.1002/fsn3.1121. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]86.
Agioutantis PC, Kotsikoris V., Kolisis FN, Loutrari H. Analiza datelor ARN-seq a celulelor de carcinom hepatocelular stimulat tratate cu galat de epigalocatechină și fisetin dezvăluie genele țintă și mecanismele de acțiune. Calculator. Struct. Biotehnologia. J. 2020; 18 :686–695. doi: 10.1016/j.csbj.2020.03.006. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]87.
Kang Q., Tong Y., Gowd V., Wang M., Chen F., Cheng KW Administrarea orală de soluție EGCG echivalentă cu dozele zilnice realizabile ale băutorilor obișnuiți de ceai suprimă în mod eficient metastaza indusă de miR483-3p a celulelor carcinomului hepatocelular în soareci. Funcție alimentară. 2021; 12 :3381–3392. doi: 10.1039/D1FO00664A. [ PubMed ] [ CrossRef ] [ Google Scholar ]88.
Yang H., Wang M., Sun H., Zhu S., Jin J. Efectul sinergetic al antagonistului receptorului EP1 și al (-)-epigallocatechin-3-galat în carcinomul hepatocelular. Farmacologie. 2019; 104 :267–275. doi: 10.1159/000502076. [ PubMed ] [ CrossRef ] [ Google Scholar ]89.
Chen RJ, Kuo HC, Cheng LH, Lee YH, Chang WT, Wang BJ, Wang YJ, Cheng HC Activitățile apoptotice și nonapoptotice ale pterostilbenului împotriva cancerului. Int. J. Mol. Sci. 2018; 19 :287. doi: 10.3390/ijms19010287. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]90.
Qian YY, Liu ZS, Yan HJ, Yuan YF, Levenson AS, Li K. Pterostilbene inhibă complexul MTA1/HDAC1 ducând la acetilarea PTEN în carcinomul hepatocelular. Biomed Pharmacother. 2018; 101 :852–859. doi: 10.1016/j.biopha.2018.03.022. [ PubMed ] [ CrossRef ] [ Google Scholar ]91.
Malhotra A., Bath S., Elbarbry F. O abordare a sistemului de organe pentru a explora acțiunile antioxidante, antiinflamatorii și citoprotectoare ale resveratrolului. Oxid Med. Cell Longev. 2015; 2015 :803971. doi: 10.1155/2015/803971. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]92.
Aja I., Ruiz-Larrea MB, Courtois A., Krisa S., Richard T., Ruiz-Sanz JI Screening of Natural Stilbene Oligomers from. Antioxidanți. 2020; 9 :469. doi: 10.3390/antiox9060469. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]93.
Dai H., Li M., Yang W., Sun X., Wang P., Wang X., Su J., Hu X., Zhao M. Resveratrolul inhibă progresia malignă a carcinomului hepatocelular prin reglarea indusă de MARCH1 de semnalizare PTEN/AKT. Îmbătrânire. 2020; 12 :11717–11731. doi: 10.18632/aging.103338. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]94.
Rawat D., Chhonker SK, Naik RA, Koiri RK Modularea enzimelor antioxidante, SIRT1 și NF-κB de către resveratrol și nicotinamidă în carcinomul hepatocelular indus de alcool-aflatoxină B1. J. Biochim. Mol. Toxicol. 2021; 35 :e22625. doi: 10.1002/jbt.22625. [ PubMed ] [ CrossRef ] [ Google Scholar ]95.
Bishayee A., Barnes KF, Bhatia D., Darvesh AS, Carroll RT Resveratrolul suprimă stresul oxidativ și răspunsul inflamator în hepatocarcinogeneza de șobolan inițiată de dietilnitrozamină. Cancer Prev. Res. 2010; 3 :753–763. doi: 10.1158/1940-6207.CAPR-09-0171. [ PubMed ] [ CrossRef ] [ Google Scholar ]96.
Dai W., Wang F., Lu J., Xia Y., He L., Chen K., Li J., Li S., Liu T., Zheng Y., și colab. Prin reducerea hexokinazei 2, resveratrolul induce apoptoza în celulele HCC dependente de glicoliză aerobă și inhibă creșterea tumorii la șoareci. Oncotarget. 2015; 6 :13703–13717. doi: 10.18632/oncotarget.3800. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]97.
Karabekir SC, Özgörgülü A. Posibile efecte protectoare ale resveratrolului în carcinomul hepatocelular. Iranul. J. Basic Med. Sci. 2020; 23 :71–78. doi: 10.22038/IJBMS.2019.36821.8774. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]98.
Rahman M., Almalki WH, Afzal O., Alfawaz Altamimi AS, Kazmi I., Al-Abbasi FA, Choudhry H., Alenezi SK, Barkat MA, Beg S., et al. Nanoparticule lipidice solide cationice de resveratrol pentru tratamentul carcinomului hepatocelular: optimizare sistematică, caracterizare in vitro și investigații preclinice. Int. J. Nanomed. 2020; 15 :9283–9299. doi: 10.2147/IJN.S277545. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]99.
Jagwani S., Jalalpure S., Dhamecha D., Jadhav K., Bohara R. Evaluarea farmacocinetică și farmacodinamică a lipozomilor cationici încărcați cu resveratrol pentru țintirea carcinomului hepatocelular. ACS Biomater. Sci. ing. 2020; 6 :4969–4984. doi: 10.1021/acsbiomaterials.0c00429. [ PubMed ] [ CrossRef ] [ Google Scholar ]100.
Mohamed GA, Al-Abd AM, El-Halawany AM, Abdallah HM, Ibrahim SRM Noi xantoni și constituenți citotoxici din coji de fructe de Garcinia mangostana împotriva liniilor celulare de cancer hepatocelular, mamar și colorectal uman. J. Etnofarmacol. 2017; 198 :302–312. doi: 10.1016/j.jep.2017.01.030. [ PubMed ] [ CrossRef ] [ Google Scholar ]101.
Wudtiwai B., Pitchakarn P., Banjerdpongchai R. Alpha-mangostin, un compus activ din Garcinia mangostana, abrogă rezistența la anoikis în celulele carcinomului hepatocelular uman. Toxicol. In vitro. 2018; 53 :222–232. doi: 10.1016/j.tiv.2018.09.003. [ PubMed ] [ CrossRef ] [ Google Scholar ]102.
Li M., Zhang M., Zhang ZL, Liu N., Han XY, Liu QC, Deng WJ, Liao CX Inducerea apoptozei de către berberină în celulele HepG2 de carcinom hepatocelular prin reglarea în jos a NF-kB. Oncol. Res. 2017; 25 :233–239. doi: 10.3727/096504016X14742891049073. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]103.
Yoon JS, Kim HM, Yadunandam AK, Kim NH, Jung HA, Choi JS, Kim CY, Kim GD. și răspunsul anti-angiogenic. Fitomedicina. 2013; 20 :1013–1022. doi: 10.1016/j.phymed.2013.03.024. [ PubMed ] [ CrossRef ] [ Google Scholar ]104.
Guo QL, You QD, Wu ZQ, Yuan ST, Zhao L. Acizii gambogici generali au inhibat creșterea celulelor hepatomului uman SMMC-7721 in vitro și la șoareci nuzi. Acta Pharmacol. Păcat. 2004; 25 :769–774. [ PubMed ] [ Google Scholar ]105.
Yan F., Wang M., Li J., Cheng H., Su J., Wang X., Wu H., Xia L., Li X., Chang HC și colab. Acidul Gambogenic a indus apoptoza dependentă de mitocondrii și s-a referit la fosfo-Erk1/2 și fosfo-p38 MAPK în celulele HepG2 de hepatom uman. Mediul. Toxicol. Pharmacol. 2012; 33 :181–190. doi: 10.1016/j.etap.2011.12.006. [ PubMed ] [ CrossRef ] [ Google Scholar ]106.
Duan D., Zhang B., Yao J., Liu Y., Sun J., Ge C., Peng S., Fang J. Acidul Gambogic induce apoptoza în celulele carcinomului hepatocelular SMMC-7721 prin țintirea tioredoxinei reductazei citosolice. Radic liber. Biol. Med. 2014; 69 :15–25. doi: 10.1016/j.freeradbiomed.2013.12.027. [ PubMed ] [ CrossRef ] [ Google Scholar ]107.
Munakarmi S., Chand L., Shin HB, Hussein UK, Yun BS, Park HR, Jeong YJ Efectele anticanceroase ale Poncirus fructus asupra carcinomului hepatocelular prin reglarea apoptozei, migrației și invaziei. Oncol. Rep. 2020; 44 :2537–2546. doi: 10.3892/or.2020.7790. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]108.
Kato R., Matsui-Yuasa I., Azuma H., Kojima-Yuasa A. Efectul sinergic al acetatului de 1′-acetoxicavicol și al butiratului de sodiu asupra morții celulelor de carcinom hepatocelular uman. Chim. Biol. Interacționa. 2014; 212 :1–10. doi: 10.1016/j.cbi.2014.01.010. [ PubMed ] [ CrossRef ] [ Google Scholar ]109.
Li Q., Li Y., Wang X., Fang X., He K., Guo X., Zhan Z., Sun C., Jin YH Tratamentul concomitent cu ginsenozid Rh2 și acid betulinic induce sinergie apoptoza la om celulele canceroase în asociere cu activarea îmbunătățită a capzazei-8, translocarea bax și eliberarea citocromului c. Mol. Carcinog. 2011; 50 :760–769. doi: 10.1002/mc.20673. [ PubMed ] [ CrossRef ] [ Google Scholar ]110.
Li M., Shang H., Wang T., Yang SQ, Li L. Decoctul Huanglian suprimă creșterea celulelor de carcinom hepatocelular prin reducerea. Lumea J. Gastroenterol. 2021; 27 :939–958. doi: 10.3748/wjg.v27.i10.939. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]111.
Huang J., Guo W., Cheung F., Tan HY, Wang N., Feng Y. Integrarea Farmacologiei de rețea și a modelelor experimentale pentru a investiga eficacitatea Coptidis și Scutellaria care conține decoct Huanglian Jiedu asupra carcinomului hepatocelular. A.m. J. Chin. Med. 2020; 48 :161–182. doi: 10.1142/S0192415X20500093. [ PubMed ] [ CrossRef ] [ Google Scholar ]112.
Wang N., Feng Y., Tan HY, Cheung F., Hong M., Lao L., Nagamatsu T. Inhibarea factorului eucariotic de alungire-2 conferă suprimarea tumorii printr-o formulare pe bază de plante decoct Huanglian-Jiedu în hepatocelular uman carcinom. J. Etnofarmacol. 2015; 164 :309–318. doi: 10.1016/j.jep.2015.02.025. [ PubMed ] [ CrossRef ] [ Google Scholar ]113.
Matsui Y., Uhara J., Satoi S., Kaibori M., Yamada H., Kitade H., Imamura A., Takai S., Kawaguchi Y., Kwon AH, et al. Prognostic îmbunătățit al pacienților cu carcinom hepatocelular postoperator atunci când sunt tratați cu alimente funcționale: un studiu de cohortă prospectiv. J. Hepatol. 2002; 37 :78–86. doi: 10.1016/S0168-8278(02)00091-0. [ PubMed ] [ CrossRef ] [ Google Scholar ]114.
Mabed M., El-Helw L., Shamaa S. Studiu de fază II al viscum fraxini-2 la pacienții cu carcinom hepatocelular avansat. Br. J. Cancer. 2004; 90 :65–69. doi: 10.1038/sj.bjc.6601463. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]115.
Zhou B., Yan Z., Liu R., Shi P., Qian S., Qu X., Zhu L., Zhang W., Wang J. Studiu prospectiv al chimioembolizării arteriale transcateter (TACE) cu Ginsenoside Rg3 versus Doar TACE pentru tratamentul pacienților cu carcinom hepatocelular avansat. Radiologie. 2016; 280 :630–639. doi: 10.1148/radiol.2016150719. [ PubMed ] [ CrossRef ] [ Google Scholar ]116.
Wang Z., Yu XL, Zhang J., Cheng ZG, Han ZY, Liu FY, Dou JP, Kong Y., Dong XJ, Zhao QX și colab. Granula Huaier previne reapariția carcinomului hepatocelular în stadiu incipient după ablația termică: un studiu de cohortă. J. Etnofarmacol. 2021; 281 :114539. doi: 10.1016/j.jep.2021.114539. [ PubMed ] [ CrossRef ] [ Google Scholar ]117.
Fan Y., Li S., Ding X., Yue J., Jiang J., Zhao H., Hao R., Qiu W., Liu K., Li Y. și colab. Icaritin, moleculă mică de modulare imuno-prima din clasă, în carcinomul hepatocelular avansat: rezultate preliminare de siguranță, supraviețuire durabilă și biomarkeri imunitari. BMC Cancer. 2019; 19 :279. doi: 10.1186/s12885-019-5471-1. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]118.
O’Reilly EM, Stuart KE, Sanz-Altamira PM, Schwartz GK, Steger CM, Raeburn L., Kemeny NE, Kelsen DP, Saltz LB Un studiu de fază II al irinotecanului la pacienții cu carcinom hepatocelular avansat. Cancer. 2001; 91 :101–105. doi: 10.1002/1097-0142(20010101)91:1<101::AID-CNCR13>3.0.CO;2-K. [ PubMed ] [ CrossRef ] [ Google Scholar ]119.
Brandi G., Biasco G., Mirarchi MG, Golfieri R., Di Paolo A., Borghi A., Fanello S., Derenzini E., Agostini V., Giampalma E., et al. Un studiu de fază I al perfuziei arteriale hepatice continue cu irinotecan la pacienții cu carcinom hepatocelular local avansat. Săpa. Ficat Dis. 2011; 43 :1015–1021. doi: 10.1016/j.dld.2011.08.005. [ PubMed ] [ CrossRef ] [ Google Scholar ]120.
Zhong C., Li HD, Liu DY, Xu FB, Wu J., Lin XM, Guo RP Studiu clinic al hepatectomiei combinate cu Terapia Jianpi Huayu pentru carcinomul hepatocelular. Pac asiatic. J. Cancer Prev. 2014; 15 :5951–5957. doi: 10.7314/APJCP.2014.15.14.5951. [ PubMed ] [ CrossRef ] [ Google Scholar ]121.
Tang CW, Zhu M., Feng WM, Bao Y., Zheng YY Medicina chineză pe bază de plante, decoctul Jianpi Ligan, îmbunătățește prognosticul carcinomului hepatocelular nerezecabil după chemoembolizarea transarterială: un studiu retrospectiv. Drug Des. Devel. Acolo. 2016; 10 :2461–2466. doi: 10.2147/DDDT.S113295. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]122.
Tang C., Feng W., Qin L., Bao Y. Chinese Herbal Medicine, Jian Pi Li Gan Decocct, Improvement Survival of Nonresecable Hepatocellular Cancer After Radiofrequency Ablation: Un studiu retrospectiv. Integr. Cancer Ther. 2018; 17 :431–436. doi: 10.1177/1534735417722223. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]123.
Wang J., Luo J., Yin X., Huang W., Cao H., Wang G., Zhou J. Jiedu Granule combinate cu chemoembolizare arterială transcateter și radiochirurgie Gamma Knife în tratarea carcinomului hepatocelular cu trombul tumoral venei portal. Biomed Res. Int. 2019; 2019 :4696843. doi: 10.1155/2019/4696843. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]124.
Zhai XF, Liu XL, Shen F., Fan J., Ling CQ Medicina tradițională pe bază de plante previne reapariția postoperatorie a carcinomului hepatocelular mic: un studiu controlat randomizat. Cancer. 2018; 124 :2161–2168. doi: 10.1002/cncr.30915. [ PubMed ] [ CrossRef ] [ Google Scholar ]125.
Lau WY, Lai EC, Leung TW, Yu SC Adjuvant intra-arterial lipiodol marcat cu iod-131 pentru carcinomul hepatocelular rezecabil: o actualizare prospectivă a studiului randomizat privind supraviețuirea la 5 și 10 ani. Ann. Surg. 2008; 247 :43–48. doi: 10.1097/SLA.0b013e3181571047. [ PubMed ] [ CrossRef ] [ Google Scholar ]126.
Lintia-Gaultier A., Perret C., Ansquer C., Eugène T., Kraeber-Bodéré F., Frampas E. Injecție intra-arterială de Lipiodol marcat cu 131I pentru carcinom hepatocelular avansat: o experiență de 7 ani. Nucl. Med. comun. 2013; 34 :674–681. doi: 10.1097/MNM.0b013e32836141a0. [ PubMed ] [ CrossRef ] [ Google Scholar ]127.
Zhou B., Shan H., Zhu KS, Jiang ZB, Guan SH, Meng XC, Zeng XC Chemoembolizare cu lobaplatină amestecată cu ulei iodat pentru carcinom hepatocelular recurent nerezecabil după transplantul hepatic ortotopic. J. Vasc. Interv. Radiol. 2010; 21 :333–338. doi: 10.1016/j.jvir.2009.11.006. [ PubMed ] [ CrossRef ] [ Google Scholar ]128.
Strumberg D., Erhard J., Harstrick A., Klaassen U., Müller C., Eberhardt W., Wilke H., Seeber S. Studiu de fază I al unei perfuzii săptămânale de 1 oră de paclitaxel la pacienții cu carcinom hepatocelular nerezecabil . EURO. J. Cancer. 1998; 34 :1290–1292. doi: 10.1016/S0959-8049(98)00054-9. [ PubMed ] [ CrossRef ] [ Google Scholar ]129.
Lin JJ, Jin CN, Zheng ML, Ouyang XN, Zeng JX, Dai XH Studiu clinic privind tratamentul carcinomului hepatocelular primar prin amestec Shenqi combinat cu coagularea cu microunde. Bărbie. J. Integr. Med. 2005; 11 :104–110. doi: 10.1007/BF02836465. [ PubMed ] [ CrossRef ] [ Google Scholar ]130.
Huang Z., Wang Y., Chen J., Wang R., Chen Q. Efectul injecției Xiaoaiping asupra carcinomului hepatocelular avansat la pacienți. J. Tradit. Bărbie. Med. 2013; 33 :34–38. doi: 10.1016/S0254-6272(13)60097-7. [ PubMed ] [ CrossRef ] [ Google Scholar ]131.
Chay WY, Tham CK, Toh HC, Lim HY, Tan CK, Lim C., Wang WW, Choo SP Coriolus versicolor (Yunzhi) Utilizare ca terapie la pacienții cu carcinom hepatocelular avansat cu funcție hepatică deficitară sau care nu sunt apți pentru terapia standard . J. Altern. Completa. Med. 2017; 23 :648–652. doi: 10.1089/acm.2016.0136. [ PubMed ] [ CrossRef ] [ Google Scholar ]132.
Sarker U., Oba S. Nutraceutice, pigmenți antioxidanti și fitochimice în frunzele speciilor de buruieni Amaranthus spinosus și Amaranthus viridis. Sci. Rep. 2019; 9 :20413. doi: 10.1038/s41598-019-50977-5. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]133.
Sharma J., Gairola S., Gaur RD, Painuli RM Tratamentul icterului cu plante medicinale în comunitățile indigene din regiunea sub-himalaya din Uttarakhand, India. J. Etnofarmacol. 2012; 143 :262–291. doi: 10.1016/j.jep.2012.06.034. [ PubMed ] [ CrossRef ] [ Google Scholar ]134.
Mondal A., Guria T., Maity TK Un nou ester al acidului gras dintr-un extract metanolic din întreaga plantă de Amaranthus spinosus și activitatea sa inhibitoare a α-glucozidazei. Farmacă. Biol. 2015; 53 :600–604. doi: 10.3109/13880209.2014.935863. [ PubMed ] [ CrossRef ] [ Google Scholar ]135.
Amuthan A., Chogtu B., Bairy KL, Prakash M. Evaluarea activității diuretice a Amaranthus spinosus Linn. extract apos la șobolani Wistar. J. Etnofarmacol. 2012; 140 :424–427. doi: 10.1016/j.jep.2012.01.049. [ PubMed ] [ CrossRef ] [ Google Scholar ]136.
Chaudhary MA, Imran I., Bashir S., Mehmood MH, Rehman NU, Gilani AH Evaluarea activităților de modulare intestinală și bronhodilatatoare ale Amaranthus spinosus Linn. Complement BMC. Altern. Med. 2012; 12 :166. doi: 10.1186/1472-6882-12-166. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]137.
Mondal A., Maity TK Activitatea antibacteriană a unui nou acid gras (14E, 18E, 22E, 26E)-metil nonacosa-14, 18, 22, 26 tetraenoat izolat din Amaranthus spinosus. Farmacă. Biol. 2016; 54 :2364–2367. doi: 10.3109/13880209.2016.1155628. [ PubMed ] [ CrossRef ] [ Google Scholar ]138.
Leung WT, Lau WY, Ho S., Chan M., Leung N., Lin J., Ho KC, Metreweli C., Johnson PJ, Li AK Radioterapia internă selectivă cu iod-131-Lipiodol intra-arterial în carcinom hepatocelular inoperabil. J. Nucl. Med. 1994; 35 :1313–1318. [ PubMed ] [ Google Scholar ]139.
Mani JS, Johnson JB, Hosking H., Ashwath N., Walsh KB, Neilsen PM, Broszczak DA, Naiker M. Potențialul antioxidant și terapeutic al plantelor australiene selectate: o revizuire. J. Etnofarmacol. 2021; 268 :113580. doi: 10.1016/j.jep.2020.113580. [ PubMed ] [ CrossRef ] [ Google Scholar ]140.
Arnesen JA, Kildegaard KR, Cernuda Pastor M., Jayachandran S., Kristensen M., Borodina I. Strains Engineered for the Production of Terpenoids. Față. Bioing. Biotehnologia. 2020; 8 :945. doi: 10.3389/fbioe.2020.00945. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]141.
Jiang M., Wu Z., Guo H., Liu L., Chen S. A Review of Terpenes from Marine-Derived Fungi: 2015–2019. Mar. Droguri. 2020; 18 :321. doi: 10.3390/md18060321. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]142.
Hillier SG, Lathe R. Terpene, hormoni și viață: regulă izoprenă revizuită. J. Endocrinol. 2019; 242 :R9–R22. doi: 10.1530/JOE-19-0084. [ PubMed ] [ CrossRef ] [ Google Scholar ]143.
Firn RD, Jones CG Produse naturale – un model simplu pentru a explica diversitatea chimică. Nat. Prod. Rep. 2003; 20 :382–391. doi: 10.1039/b208815k. [ PubMed ] [ CrossRef ] [ Google Scholar ]144.
Thoppil RJ, Bishayee A. Terpenoizii ca potențiali agenți chimiopreventivi și terapeutici în cancerul de ficat. Lumea J. Hepatol. 2011; 3 :228–249. doi: 10.4254/wjh.v3.i9.228. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]145.
Dasgupta A., Acharya K. Ciuperci: O resursă emergentă pentru terpenoide terapeutice. 3 Biotehnologie. 2019; 9 :369. doi: 10.1007/s13205-019-1906-2. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]146.
Sullivan R., Smith JE, Rowan NJ Ciuperci medicinale și terapia cancerului: traducerea unei practici tradiționale în medicina occidentală. Perspectivă. Biol. Med. 2006; 49 :159–170. doi: 10.1353/pbm.2006.0034. [ PubMed ] [ CrossRef ] [ Google Scholar ]147.
Standish LJ, Wenner CA, Sweet ES, Bridge C., Nelson A., Martzen M., Novack J., Torkelson C. Trametes versicolor mushroom immune therapy in cancer breast. J. Soc. Integr. Oncol. 2008; 6 :122–128. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]148.
Gao QH, Wu CS, Wang M. Fructul jujube (Ziziphus jujuba Mill.): O revizuire a cunoștințelor actuale despre compoziția fructelor și beneficiile pentru sănătate. J. Agric. Food Chim. 2013; 61 :3351–3363. doi: 10.1021/jf4007032. [ PubMed ] [ CrossRef ] [ Google Scholar ]150.
Liu J. Farmacologia acidului oleanolic și acidului ursolic. J. Etnofarmacol. 1995; 49 :57–68. doi: 10.1016/0378-8741(95)90032-2. [ PubMed ] [ CrossRef ] [ Google Scholar ]151.
Yan SL, Huang CY, Wu ST, Yin MC Acidul oleanolic și acidul ursolic induc apoptoza în patru linii celulare de cancer hepatic uman. Toxicol. In vitro. 2010; 24 :842–848. doi: 10.1016/j.tiv.2009.12.008. [ PubMed ] [ CrossRef ] [ Google Scholar ]152.
Yang Y., Qiu S., Qian L., Tian Y., Chen Y., Bi L., Chen W. OCF poate reprima metastaza tumorii prin inhibarea tranziției epitelial-mezenchimale implicate în calea PTEN/PI3K/AKT în plămân celule canceroase. Plus unu. 2017; 12 :e0174021. doi: 10.1371/journal.pone.0174021. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]153.
Wojcikowski K., Gobe G. Studii pe animale asupra ierburilor medicinale: predictibilitate, conversie de doză și valoare potențială. Phytother. Res. 2014; 28 :22–27. doi: 10.1002/ptr.4966. [ PubMed ] [ CrossRef ] [ Google Scholar ]154.
Xiong J., Cheng G., Tang H., Zhen HN, Zhang X. Ardipusilloside I induce apoptoza în celulele de glioblastom uman printr-o cale de semnalizare FasL/Fas independentă de caspază-8. Mediul. Toxicol. Pharmacol. 2009; 27 :264–270. doi: 10.1016/j.etap.2008.11.008. [ PubMed ] [ CrossRef ] [ Google Scholar ]155.
Talevi A., Quiroga AMP ADME Processes in Pharmaceutical Sciences. Springer; Cham, Elveția: 2018. [ Google Scholar ]156.
Raj Marshall G., Raveendran R. Introducere în Bazele Farmacologiei și Toxicologiei. Farmacologie generală și moleculară: principiile acțiunii medicamentului. Volumul 1 Springer; Singapore: 2019. [ Google Scholar ]157.
Reddy D., Kumavath R., Ghosh P., Barh D. Lanatoside C induce oprirea ciclului celular G2/M și suprimă creșterea celulelor canceroase prin atenuarea căilor de semnalizare MAPK, Wnt, JAK-STAT și PI3K/AKT/mTOR. Biomolecule. 2019; 9 :792. doi: 10.3390/biom9120792. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]158.
Chen LI, Guo Y., Alsaif G., Gao Y. Total Flavonoids Isolated from. Anticancer Res. 2020; 40 :5201–5210. doi: 10.21873/anticanres.14523. [ PubMed ] [ CrossRef ] [ Google Scholar ]159.
Zhu AK, Zhou H., Xia JZ, Jin HC, Wang K., Yan J., Zuo JB, Zhu X., Shan T. Apoptoza indusă de Ziyuglycoside II în celulele BGC-823 de carcinom gastric uman prin reglarea Bax/ Expresia Bcl-2 și calea de activare a caspazei-3. Braz. J. Med. Biol. Res. 2013; 46 :670–675. doi: 10.1590/1414-431X20133050. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]160.
Zhu X., Wang K., Zhang K., Huang B., Zhang J., Zhang Y., Zhu L., Zhou B., Zhou F. Ziyuglycoside II inhibă creșterea carcinomului mamar uman MDA-MB- 435 prin oprirea ciclului celular și inducerea apoptozei prin calea dependentă de mitocondrii. Int. J. Mol. Sci. 2013; 14 :18041–18055. doi: 10.3390/ijms140918041. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]161.
Lkhagvasuren K., Kim JK Ziyuglycoside II induce apoptoza dependentă de caspaze și independentă de caspaze în celulele canceroase de colon umane. Toxicol. In vitro. 2019; 59 :255–262. doi: 10.1016/j.tiv.2019.04.028. [ PubMed ] [ CrossRef ] [ Google Scholar ]162.
Chen HM, Lai ZQ, Liao HJ, Xie JH, Xian YF, Chen YL, Ip SP, Lin ZX, Su ZR Efectul antitumoral sinergic al brusatolului combinat cu cisplatină asupra celulelor cancerului colorectal. Int. J. Mol. Med. 2018; 41 :1447–1454. doi: 10.3892/ijmm.2018.3372. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]163.
Vartanian S., Ma TP, Lee J., Haverty PM, Kirkpatrick DS, Yu K., Stokoe D. Application of Mass Spectrometry Profiling to Establish Brusatol as an inhibitor of Global Protein Synthesis. Mol. Celulă. Proteom. 2016; 15 :1220–1231. doi: 10.1074/mcp.M115.055509. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]164.
Lu Z., Lai ZQ, Leung AWN, Leung PS, Li ZS, Lin ZX Explorarea brusatolului ca nou adjuvant anti-cancer pancreatic: evaluare biologică și studii mecanice. Oncotarget. 2017; 8 :84974–84985. doi: 10.18632/oncotarget.17761. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]165.
Zhao Q., Li P., Jiang J., Hu P. Pharmacokinetics of Single Ascending Does and Multiple Dozes of 20(S)-Ginsenoside Rg3 in Chinese Healthy Volunteers. EURO. J. Drug Metab. Farmacokineta. 2016; 41 :845–853. doi: 10.1007/s13318-015-0304-3. [ PubMed ] [ CrossRef ] [ Google Scholar ]171.
Ganesan K., Xu B. A Critical Review on Polyphenols and Health Benefits of Black Soybeans. Nutrienți. 2017; 9 :455. doi: 10.3390/nu9050455. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]172.
Dai J., Mumper RJ Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecule. 2010; 15 :7313–7352. doi: 10,3390/molecules15107313. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]173.
Anantharaju PG, Gowda PC, Vimalambike MG, Madhunapantula SV O privire de ansamblu asupra rolului fenolicilor dietetici pentru tratamentul cancerelor. Nutr. J. 2016; 15:99 . doi: 10.1186/s12937-016-0217-2. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]174.
Kikuchi H., Yuan B., Hu X., Okazaki M. Activitatea chimiopreventivă și anticanceroasă a flavonoidelor și posibilitatea acesteia de utilizare clinică prin combinarea cu agenți chimioterapeutici convenționali. A.m. J. Cancer Res. 2019; 9 :1517–1535. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]175.
Abotaleb M., Samuel SM, Varghese E., Varghese S., Kubatka P., Liskova A., Büsselberg D. Flavonoids in Cancer and Apoptosis. cancere. 2018; 11:28 . doi: 10.3390/cancers11010028. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]176.
Banjerdpongchai R., Wudtiwai B., Khaw-On P., Rachakhom W., Duangnil N., Kongtawelert P. Hesperidin din semințele de citrice induce apoptoza celulelor hepatocelulare a carcinomului uman HepG2 atât prin căile mitocondriale, cât și ale receptorilor morții. Tumor Biol. 2016; 37 :227–237. doi: 10.1007/s13277-015-3774-7. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]177.
Wang Z., Zhang H., Zhou J., Zhang X., Chen L., Chen K., Huang Z. Eriocitrina din lămâie suprimă proliferarea celulelor de carcinom hepatocelular uman prin inducerea apoptozei și oprirea ciclului celular. Cancer Chemother. Pharmacol. 2016; 78 :1143–1150. doi: 10.1007/s00280-016-3171-y. [ PubMed ] [ CrossRef ] [ Google Scholar ]178.
Xie D., Yuan P., Wang D., Jin H., Chen H. Efectele naringinei asupra expresiei miR-19b și apoptozei celulare în carcinomul hepatocelular uman. Oncol. Lett. 2017; 14 :1455–1459. doi: 10.3892/ol.2017.6278. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]179.
Kuo CJ, Conley PB, Chen L., Sladek FM, Darnell JE, Crabtree GR O ierarhie transcripțională implicată în specificarea tipului de celule de mamifere. Natură. 1992; 355 :457–461. doi: 10.1038/355457a0. [ PubMed ] [ CrossRef ] [ Google Scholar ]180.
Saha SK, Parachoniak CA, Ghanta KS, Fitamant J., Ross KN, Najem MS, Gurumurthy S., Akbay EA, Sia D., Cornella H., et al. IDH mutant inhibă HNF-4α pentru a bloca diferențierea hepatocitelor și pentru a promova cancerul biliar. Natură. 2014; 513 :110–114. doi: 10.1038/nature13441. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]181.
Lincet H., Icard P. Cum favorizează enzimele glicolitice proliferarea celulelor canceroase prin funcții nemetabolice? Oncogene. 2015; 34 :3751–3759. doi: 10.1038/onc.2014.320. [ PubMed ] [ CrossRef ] [ Google Scholar ]182.
Mayer D., Klimek F., Rempel A., Bannasch P. Expresia hexokinazei în preneoplazia și neoplazia ficatului. Biochim. Soc. Trans. 1997; 25 :122–127. doi: 10.1042/bst0250122. [ PubMed ] [ CrossRef ] [ Google Scholar ]183.
Arai Y., Watanabe S., Kimira M., Shimoi K., Mochizuki R., Kinae N. Aporturile dietetice de flavonoli, flavone și izoflavone de către femeile japoneze și corelația inversă dintre aportul de quercetină și concentrația plasmatică a colesterolului LDL. J. Nutr. 2000; 130 :2243–2250. doi: 10.1093/jn/130.9.2243. [ PubMed ] [ CrossRef ] [ Google Scholar ]184.
Hanneken A., Lin FF, Johnson J., Maher P. Flavonoidele protejează celulele epiteliale pigmentare ale retinei umane de moartea indusă de stresul oxidativ. Investig. Oftalmol. Vis. Sci. 2006; 47 :3164–3177. doi: 10.1167/iovs.04-1369. [ PubMed ] [ CrossRef ] [ Google Scholar ]185.
Salmela AL, Pouwels J., Varis A., Kukkonen AM, Toivonen P., Halonen PK, Perälä M., Kallioniemi O., Gorbsky GJ, Kallio MJ. punct de control. Carcinogeneza. 2009; 30 :1032–1040. doi: 10.1093/carcin/bgp101. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]186.
Khan N., Afaq F., Syed DN, Mukhtar H. Fisetin, un flavonoid alimentar nou, provoacă apoptoza și oprirea ciclului celular în celulele LNCaP ale cancerului de prostată uman. Carcinogeneza. 2008; 29 :1049–1056. doi: 10.1093/carcin/bgn078. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]187.
Youns M., Abdel Halim Hegazy W. Fisetina flavonoidă naturală inhibă proliferarea celulară a celulelor canceroase hepatice, colorectale și pancreatice prin modularea căilor multiple de semnalizare. Plus unu. 2017; 12 :e0169335. doi: 10.1371/journal.pone.0169335. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]188.
Lee SR, Kwon SW, Lee YH, Kaya P., Kim JM, Ahn C., Jung EM, Lee GS, An BS, Jeung EB și colab. Aportul alimentar de genisteine suprimă carcinomul hepatocelular prin apoptoză mediată de AMPK și antiinflamație. BMC Cancer. 2019; 19 :6. doi: 10.1186/s12885-018-5222-8. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]189.
Zhang L., Liu P., Li L., Huang Y., Pu Y., Hou X., Song L. Identificarea și activitatea antioxidantă a flavonoidelor extrase din Xinjiang Jujube. Molecule. 2018; 24 :122. doi: 10.3390/molecules24010122. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]190.
Zhou J., Wu J., Chen X., Fortenbery N., Eksioglu E., Kodumudi KN, Pk EB, Dong J., Djeu JY, Wei S. Icariin și derivatul său, TIC, exercită antiinflamator, efecte anti-tumorale și modulează funcțiile celulelor supresoare derivate mieloide (MDSC). Int. Imunofarmacol. 2011; 11 :890–898. doi: 10.1016/j.intimp.2011.01.007. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]191.
Zhao H., Guo Y., Li S., Han R., Ying J., Zhu H., Wang Y., Yin L., Han Y., Sun L. și colab. Un nou agent anti-cancer Icaritin suprimă inițierea carcinomului hepatocelular și creșterea malignă prin calea IL-6/Jak2/Stat3. Oncotarget. 2015; 6 :31927–31943. doi: 10.18632/oncotarget.5578. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]192.
Luo XY, Wu KM, He XX Progrese în dezvoltarea de medicamente pentru carcinomul hepatocelular: studii clinice și potențiale ținte terapeutice. J. Exp. Clin. Cancer Res. 2021; 40 :172. doi: 10.1186/s13046-021-01968-w. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]193.
Nakagawa S., Wei L., Song WM, Higashi T., Ghoshal S., Kim RS, Bian CB, Yamada S., Sun X., Venkatesh A., et al. Prevenirea moleculară a cancerului hepatic în ciroză prin analiza transcriptomului de organ și inhibarea căii acidului lisofosfatidic. Celula canceroasă. 2016; 30 :879–890. doi: 10.1016/j.ccell.2016.11.004. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]194.
McCormack D., McFadden D. Pterostilbene and cancer: Current review. J. Surg. Res. 2012; 173 :e53–e61. doi: 10.1016/j.jss.2011.09.054. [ PubMed ] [ CrossRef ] [ Google Scholar ]195.
Paoli P., Giannoni E., Chiarugi P. Anoikis molecular pathways and its role in cancer progression. Biochim. Biophys. Acta. 2013; 1833 :3481–3498. doi: 10.1016/j.bbamcr.2013.06.026. [ PubMed ] [ CrossRef ] [ Google Scholar ]196.
Yang S., Zhou F., Dong Y., Ren F. α-Mangostin induce apoptoza în celulele de osteosarcom uman prin stresul reticulului endoplasmatic mediat de ROS prin calea WNT. Transplant de celule. 2021; 30 :9636897211035080. doi: 10.1177/09636897211035080. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]197.
Panthong A., Norkaew P., Kanjanapothi D., Taesotikul T., Anantachoke N., Reutrakul V. Activități antiinflamatorii, analgezice și antipiretice ale extractului de gamboge din Garcinia hanburyi Hook f. J. Etnofarmacol. 2007; 111 :335–340. doi: 10.1016/j.jep.2006.11.038. [ PubMed ] [ CrossRef ] [ Google Scholar ]198.
Qiu S., Sun H., Zhang AH, Xu HY, Yan GL, Han Y., Wang XJ Natural alcaloids: Basic aspects, biological roles, and future perspectives. Bărbie. J. Nat. Med. 2014; 12 :401–406. doi: 10.1016/S1875-5364(14)60063-7. [ PubMed ] [ CrossRef ] [ Google Scholar ]199.
Setiawan VW, Wilkens LR, Lu SC, Hernandez BY, Le Marchand L., Henderson BE Asociația aportului de cafea cu incidență redusă a cancerului hepatic și a decesului din cauza bolii hepatice cronice în cohorta multietnică din SUA. Gastroenterologie. 2015; 148 :118–125. doi: 10.1053/j.gastro.2014.10.005. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]200.
Liu F., Wang X., Wu G., Chen L., Hu P., Ren H., Hu H. Consumul de cafea scade riscurile de fibroză hepatică și ciroză: o meta-analiză. Plus unu. 2015; 10 :e0142457. doi: 10.1371/journal.pone.0142457. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]201.
Cavin C., Marin-Kuan M., Langouët S., Bezençon C., Guignard G., Verguet C., Piguet D., Holzhäuser D., Cornaz R., Schilter B. Inducerea apărării celulare mediate de Nrf2 și alterarea activităților de fază I ca mecanisme ale efectelor chimioprotectoare ale cafelei în ficat. Food Chim. Toxicol. 2008; 46 :1239–1248. doi: 10.1016/j.fct.2007.09.099. [ PubMed ] [ CrossRef ] [ Google Scholar ]202.
Petrick JL, Freedman ND, Graubard BI, Sahasrabuddhe VV, Lai GY, Alavanja MC, Beane-Freeman LE, Boggs DA, Buring JE, Chan AT și colab. Consumul de cafea și riscul de carcinom hepatocelular și colangiocarcinom intrahepatic în funcție de sex: proiectul de grupare a cancerului de ficat. Epidemiol de cancer. Biomark. Prev. 2015; 24 :1398–1406. doi: 10.1158/1055-9965.EPI-15-0137. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]203.
Bamia C., Lagiou P., Jenab M., Trichopoulou A., Fedirko V., Aleksandrova K., Pischon T., Overvad K., Olsen A., Tjønneland A., et al. Cafeaua, ceaiul și cafeaua decofeinizată în relație cu carcinomul hepatocelular într-o populație europeană: studiu de cohortă prospectiv, multicentric. Int. J. Cancer. 2015; 136 :1899–1908. doi: 10.1002/ijc.29214. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]204.
Wang H., Mu W., Shang H., Lin J., Lei X. Efectele antihiperglicemice ale Rhizoma Coptidis și mecanismul de acțiuni: O revizuire a revizuirilor sistematice și a cercetării farmacologice. Biomed Res. Int. 2014; 2014 :798093. doi: 10.1155/2014/798093. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]205.
Reyhanoglu G., Smith T. Irinotecan. Editura StatPearls; Treasure Island, FL, SUA: 2021. [ Google Scholar ]206.
Matsushita K., Kuramitsu Y., Ohiro Y., Obara M., Kobayashi M., Li YQ, Hosokawa M. Terapia combinată a compusului activ corelat cu hexoză plus UFT reduce semnificativ metastaza adenocarcinomului mamar la șobolan. Medicamente anticancerigene. 1998; 9 :343–350. doi: 10.1097/00001813-199804000-00008. [ PubMed ] [ CrossRef ] [ Google Scholar ]207.
Shin MS, Park HJ, Maeda T., Nishioka H., Fujii H., Kang I. The Effects of AHCC®, a Standardized Extract of Cultured. J. Immunol. Res. 2019; 2019 :3758576. doi: 10.1155/2019/3758576. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]208.
Spierings EL, Fujii H., Sun B., Walshe T. Un studiu de fază I al siguranței suplimentului nutrițional, compus activ corelat cu hexoză, AHCC, la voluntari sănătoși. J. Nutr. Sci. Vitaminol. 2007; 53 :536–539. doi: 10.3177/jnsv.53.536. [ PubMed ] [ CrossRef ] [ Google Scholar ]209.
Cao Z., Chen X., Lan L., Zhang Z., Du J., Liao L. Compusul activ corelat cu hexoză potențează efectele antitumorale ale 5-fluorouracilului în doză mică prin modularea funcției imune în hepatomul 22 tumoral- purtător de șoareci. Nutr. Res. Practică. 2015; 9 :129–136. doi: 10.4162/nrp.2015.9.2.129. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]210.
Pan J., Yang C., Jiang Z., Huang J. Murr: O medicină tradițională chineză cu efecte anti-tumorale puternice. Managementul cancerului. Res. 2019; 11 :1541–1549. doi: 10.2147/CMAR.S193174. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]211.
Zhang T., Wang K., Zhang J., Wang X., Chen Z., Ni C., Qiu F., Huang J. Huaier extractul apos inhibă creșterea celulelor stem de cancer colorectal parțial prin reglarea în jos a Wnt/β -calea cateninei. Oncol. Lett. 2013; 5 :1171–1176. doi: 10.3892/ol.2013.1145. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]212.
Hu Z., Yang A., Su G., Zhao Y., Wang Y., Chai X., Tu P. Huaier limitează potențialul proliferativ și invaziv al celulelor SKHEP-1 ale hepatomului uman, parțial prin scăderea Lamin B1 și creșterea NOV . Sci. Rep. 2016; 6 :31298. doi: 10.1038/srep31298. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]213.
Qi W., Sun M., Kong X., Li Y., Wang X., Lv S., Ding X., Gao S., Cun J., Cai C. și colab. Extractul Huaier face sinergie cu tamoxifenul pentru a induce autofagia și apoptoza în celulele canceroase de sân ER-pozitive. Oncotarget. 2016; 7 :26003–26015. doi: 10.18632/oncotarget.8303. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]214.
Shan L., Li Y., Jiang H., Tao Y., Qian Z., Li L., Cai F., Ma L., Yu Y. Huaier limitează potențialul de proliferare și migrație al celulelor de carcinom hepatocelular parțial prin scăderea Da-proteina asociată 1. J. Cancer. 2017; 8 :4087–4097. doi: 10.7150/jca.21018. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]215.
Chen Q., Shu C., Laurence AD, Chen Y., Peng BG, Zhen ZJ, Cai JQ, Ding YT, Li LQ, Zhang YB și colab. Efectul granulelor Huaier asupra recurenței după rezecția curativă a HCC: un studiu clinic multicentric, randomizat. Intestin. 2018; 67 :2006–2016. doi: 10.1136/gutjnl-2018-315983. [ PubMed ] [ CrossRef ] [ Google Scholar ]216.
Pham HH, Seong YA, Oh CW, Kim GD Medicamentul pe bază de plante Cyperus amuricus inhibă proliferarea celulelor Hep3B ale carcinomului hepatocelular uman prin inducerea apoptozei și oprirea în faza ciclului celular G0/G1. Int. J. Oncol. 2016; 49 :2046–2054. doi: 10.3892/ijo.2016.3698. [ PubMed ] [ CrossRef ] [ Google Scholar ]218.
Lee RT, Yang P., Alahmadi A., McQuade J., Yuan E., Difeo A., Narla G., Kaseb A. Extract de vâsc Viscum Fraxini-2 pentru tratamentul carcinomului hepatocelular avansat: o serie de cazuri. Caz Rep. Oncol. 2021; 14 :224–231. doi: 10.1159/000511566. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]219.
Galle PR, Foerster F., Kudo M., Chan SL, Llovet JM, Qin S., Schelman WR, Chintharlapalli S., Abada PB, Sherman M., et al. Biologia și semnificația alfa-fetoproteinei în carcinomul hepatocelular. Ficat Int. 2019; 39 :2214–2229. doi: 10.1111/liv.14223. [ PubMed ] [ CrossRef ] [ Google Scholar ]220.
Ohnishi H., Asamoto M., Tujimura K., Hokaiwado N., Takahashi S., Ogawa K., Kuribayashi M., Ogiso T., Okuyama H., Shirai T. Inhibarea proliferării celulare de către nobiletin, o dietă fitochimic, asociat cu apoptoza și expresia genelor caracteristice, dar lipsa efectului asupra hepatocarcinogenezei timpurii la șobolan in vivo. Cancer Sci. 2004; 95 :936–942. doi: 10.1111/j.1349-7006.2004.tb03180.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]221.
Yaku K., Matsui-Yuasa I., Azuma H., Kojima-Yuasa A. Acetatul de 1′-acetoxichavicol îmbunătățește activitățile enzimatice de fază II prin creșterea nivelului intranuclear de Nrf2 și a nivelului citosolic p21. A.m. J. Chin. Med. 2011; 39 :789–802. doi: 10.1142/S0192415X11009196. [ PubMed ] [ CrossRef ] [ Google Scholar ]222.
Wu JT, Archer SY, Hinnebusch B., Meng S., Hodin RA Tranzitorie vs hiperacetilarea prelungită a histonelor: Efecte asupra creșterii celulelor cancerului de colon, diferențiere și apoptoză. A.m. J. Physiol. Test gastro-intestinal. Ficat Physiol. 2001; 280 :G482–G490. doi: 10.1152/ajpgi.2001.280.3.G482. [ PubMed ] [ CrossRef ] [ Google Scholar ]223.
Haga A., Manning AM, Hanlon KA, Huschtscha LI, Hart D., Paraskeva C. Butiratul de sodiu induce apoptoza în liniile celulare tumorale colonice umane într-o cale independentă de p53: Implicații pentru posibilul rol al fibrelor alimentare în prevenire a cancerului de intestin gros. Int. J. Cancer. 1993; 55 :498–505. doi: 10.1002/ijc.2910550329. [ PubMed ] [ CrossRef ] [ Google Scholar ]224.
Gleave ME, Sato N., Sadar M., Yago V., Bruchovsky N., Sullivan L. Analog de butirat, izobutiramida, inhibă creșterea tumorii și timpul până la progresia independentă de androgeni în modelul tumoral LNCaP de prostată umană. J. Cell Biochem. 1998; 69 :271–281. doi: 10.1002/(SICI)1097-4644(19980601)69:3<271::AID-JCB5>3.0.CO;2-O. [ PubMed ] [ CrossRef ] [ Google Scholar ]225.
Yamamoto H., Fujimoto J., Okamoto E., Furuyama J., Tamaoki T., Hashimoto-Tamaoki T. Suprimarea creșterii carcinomului hepatocelular prin butirat de sodiu in vitro și in vivo. Int. J. Cancer. 1998; 76 :897–902. doi: 10.1002/(SICI)1097-0215(19980610)76:6<897::AID-IJC21>3.0.CO;2-Z. [ PubMed ] [ CrossRef ] [ Google Scholar ]226.
Pisha E., Chai H., Lee IS, Chagwedera TE, Farnsworth NR, Cordell GA, Beecher CW, Fong HH, Kinghorn AD, Brown DM Descoperirea acidului betulinic ca inhibitor selectiv al melanomului uman care funcționează prin inducerea apoptozei . Nat. Med. 1995; 1 :1046–1051. doi: 10.1038/nm1095-1046. [ PubMed ] [ CrossRef ] [ Google Scholar ]227.
Fulda S., Jeremias I., Pietsch T., Debatin KM Acidul betulinic: Un nou agent chimioterapeutic în tratamentul tumorilor neuroectodermale. Klin Padiatr. 1999; 211 :319–322. doi: 10.1055/s-2008-1043808. [ PubMed ] [ CrossRef ] [ Google Scholar ]228.
Zuco V., Supino R., Righetti SC, Cleris L., Marchesi E., Gambacorti-Passerini C., Formelli F. Selective cytotoxicity of betulinic acid on tumor cell lines, but not on normal cells. Rac Lett. 2002; 175 :17–25. doi: 10.1016/S0304-3835(01)00718-2. [ PubMed ] [ CrossRef ] [ Google Scholar ]229.
Tatsuka M., Maeda M., Ota T. Efectul anticancerigen și creșterea potențialului metastatic al celulelor BALB/c 3T3 de către ginsenozid Rh(2) Jpn. J. Cancer Res. 2001; 92 :1184–1189. doi: 10.1111/j.1349-7006.2001.tb02138.x. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]230.
Nakata H., Kikuchi Y., Tode T., Hirata J., Kita T., Ishii K., Kudoh K., Nagata I., Shinomiya N. Efectele inhibitorii ale ginsenozidei Rh2 asupra creșterii tumorii la șoarecii nuzi purtători de oameni celule de cancer ovarian. Jpn. J. Cancer Res. 1998; 89 :733–740. doi: 10.1111/j.1349-7006.1998.tb03278.x. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]231.
Odashima S., Ohta T., Kohno H., Matsuda T., Kitagawa I., Abe H., Arichi S. Controlul exprimării fenotipice a celulelor de melanom B16 cultivate prin glicozide vegetale. Cancer Res. 1985; 45 :2781–2784. [ PubMed ] [ Google Scholar ]232.
Zhu JH, Takeshita T., Kitagawa I., Morimoto K. Suprimarea formării schimburilor de cromatide surori prin concentrații scăzute de ginsenozidă Rh2 în limfocitele din sângele uman. Cancer Res. 1995; 55 :1221–1223. [ PubMed ] [ Google Scholar ]233.
Song Y., Sun B., Hao L., Hu J., Du S., Zhou X., Zhang L., Liu L., Gong L., Chi X. și colab. Expresia crescută a factorului de alungire eucariotic 2 este implicată în proliferarea și invazia carcinomului cu celule scuamoase pulmonare. Oncotarget. 2016; 7 :58470–58482. doi: 10.18632/oncotarget.11298. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]234.
Liu X., Li M., Wang X., Dang Z., Yu L., Jiang Y., Yang Z. Efectele terapiei adjuvante de medicină tradițională chineză asupra supraviețuirii pe termen lung la pacienții cu carcinom hepatocelular. Fitomedicina. 2019; 62 :152930. doi: 10.1016/j.phymed.2019.152930. [ PubMed ] [ CrossRef ] [ Google Scholar ]235.
Liu Y., Li Y., Wang X., Huang Y., Zhang Q., Shi K., Ran C., Hou J. Capsule, a Traditional Chinese Medicinal Formulation, Enhances the Supravious of Patients with Hepatocellular Carcinom and Vp3-4 Tromboză tumorală venoasă portală în curs de tratament de susținere. J. Altern. Completa. Med. 2020; 26 :956–965. doi: 10.1089/acm.2019.0334. [ PubMed ] [ CrossRef ] [ Google Scholar ]236.
Tsai TY, Livneh H., Hung TH, Lin IH, Lu MC, Yeh CC Asociații între medicina pe bază de plante chineză prescrisă și riscul de carcinom hepatocelular la pacienții cu hepatită cronică B: un studiu de cohortă la nivel național. BMJ Open. 2017; 7 :e014571. doi: 10.1136/bmjopen-2016-014571. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]237.
Tsai FJ, Cheng CF, Chen CJ, Lin CY, Wu YF, Li TM, Chuang PH, Wu YC, Lai CH, Liu X. și colab. Efectele terapiei cu plante medicinale chinezești asupra supraviețuirii și a rezultatelor hepatice la pacienții cu infecție cu virusul hepatitei C din Taiwan. Fitomedicina. 2019; 57 :30–38. doi: 10.1016/j.phymed.2018.09.237. [ PubMed ] [ CrossRef ] [ Google Scholar ]238.
Qi F., Zhao L., Zhou A., Zhang B., Li A., Wang Z., Han J. Avantajele utilizării medicinei tradiționale chineze ca terapie adjuvantă în întregul curs de tratament al cancerului în loc de numai stadiul terminal al cancerului. Biosci. Tendințe. 2015; 9 :16–34. doi: 10.5582/bst.2015.01019. [ PubMed ] [ CrossRef ] [ Google Scholar ]239.
Jingjing H., Hongna H., Xiaojiao W., Yan G., Yuexue Z., Yueqiang H. Pilula Bie Jia Jian îmbunătățește ameliorarea celulelor stem mezenchimale osoase în progresia carcinomului hepatocelular. J. Nat. Med. 2021 doi: 10.1007/s11418-021-01548-4. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]240.
Huang Y., Zhou C., Wen H., Chen Y., Xie Y., Lan X., Lin J., Huang X., Mo Y., Yang C. și colab. Formula Jianpi-Huayu inhibă dezvoltarea carcinomului hepatocelular prin reglarea expresiei miR-602, care vizează. Integr. Cancer Ther. 2020; 19 :1534735419900804. doi: 10.1177/1534735419900804. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]241.
Maru GB, Hudlikar RR, Kumar G., Gandhi K., Mahimkar MB Înțelegerea mecanismelor moleculare de prevenire a cancerului prin fitochimice dietetice: de la modele experimentale la studii clinice. Lumea J. Biol. Chim. 2016; 7 :88–99. doi: 10.4331/wjbc.v7.i1.88. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
Articole de
la Cancers sunt furnizate aici prin amabilitatea
Multidisciplinary Digital Publishing Institute (MDPI)
Cancerele ginecologice sunt printre principalele cauze de mortalitate asociată cancerului la nivel mondial. În timp ce numărul cazurilor este în creștere, abordările terapeutice actuale nu sunt suficient de eficiente. Există efecte secundare considerabile, precum și tipuri rezistente la tratament. În plus, toate acestea fac tratamentul complicat pentru cazurile afectate. Prin urmare, pentru a îmbunătăți eficacitatea procesului de tratament și calitatea vieții pacienților, căutarea unor noi tratamente adjuvante este foarte justificată. Curcumina, un compus natural promițător, este înzestrat cu numeroase potențiale terapeutice, inclusiv efecte anticancerigene semnificative. Recent, diverse investigații au demonstrat efectele anticancer ale curcuminei și ale analogilor săi noi asupra cancerelor ginecologice. În plus, noi formulări de curcumină au avut ca rezultat efecte favorabile suplimentare. Această revizuire discută aceste studii și evidențiază posibilele mecanisme de bază ale efectelor observate.
fundal
S-a demonstrat că compuși naturali precum curcumina, epigalocatechin galat (EGCG), quercetina și resveratrolul modulează unele mecanisme genetice și epigenetice, care pot crește sensibilitatea celulelor canceroase la agenții convenționali și astfel inhibă creșterea tumorii [1 , 2 ] . Curcumina, principalul ingredient bioactiv al turmericului, s-a dovedit că posedă diferite potențiale terapeutice, cum ar fi antitumoral [ 3 , 4 ], anti-aterosclerotic [ 5 , 6 ], anti-microbian [ 7 ], anti-oxidant [ 8 ] ] și antiinflamatoare [ 9 , 10 , 11 ]. O mare varietate de ținte moleculare au fost raportate pentru curcumină, inclusiv unii factori de transcripție, modulatori genici, kinaze, factori de creștere și receptori de membrană celulară [ 12 , 13 , 14 , 15 , 16 ]. Datorită funcțiilor sale diverse și pleiotrope, un număr tot mai mare de studii s-au concentrat pe curcumină, în special în afecțiunile maligne.
Numeroase investigații au arătat că curcumina acționează ca un adjuvant benefic și agent chimiopreventiv pentru cancer [ 17 ]. În special, studiile pe oameni au demonstrat tolerabilitatea, siguranța și capacitatea anticancerigenă a acestui compus pentru oameni [ 13 , 14 ]. Rezultatul promițător al acestor studii a contribuit la creșterea speranței pentru un viitor mai luminos în tratamentul cancerului.
Cancerele ginecologice includ o creștere nelimitată și anormală a celulelor, care se dezvoltă în sistemul reproducător feminin, cum ar fi afecțiunile maligne endometriale, ovariene, cervicale, peritoneale primare, vulvare și vaginale. Potrivit rapoartelor, peste 1 milion de pacienți au fost nou diagnosticați și peste 580 000 de decese au avut loc din cauza cancerelor endometriale, de col uterin și ovar în 2018 [15 ] . În timp ce cancerele endometriale și cervicale sunt diagnosticate în stadii incipiente, cancerul ovarian este de obicei diagnosticat în stadii mai avansate, când tratamentul este mai dificil [ 16 , 18 ]. În ciuda tuturor evoluțiilor în terapia cancerului și a apariției unor tratamente noi, aceste tipuri de cancer au încă rate considerabile de mortalitate. Prin urmare, este foarte necesară introducerea de noi agenți care pot îmbunătăți rezultatul terapeutic pentru acești pacienți afectați. Aici, în această revizuire, ne concentrăm pe dovezile actuale pentru eficacitatea curcuminei și a formulărilor sale noi în cancerele ginecologice.
Rolul curcuminei în cancerele ginecologice
În ceea ce privește cancerele ginecologice, s-a demonstrat că curcumina modifică efectele factorilor de risc în cursul progresiei cancerului și încă de la început. Unii dintre acești factori de risc cunoscuți sunt obezitatea, fumatul, estrogenul și infecțiile cu virusul papiloma uman (HPV) [ 19 , 20 ]. Interesant, s-a demonstrat că curcumina reduce sintetizarea estrogenului, deteriorarea ADN-ului derivat din estrogen, inflamația în țesutul adipos și inhibă expresia oncoproteinei E6 și E7 în HPV [19 ] .
Conform studiilor in vitro, curcumina poate preveni invazia și migrarea celulelor de carcinom endometrial (EC) și joacă un rol anti-metastatic datorită reducerii producției și funcționării metaloproteinazelor matriceale-2 și -9 (MMP-2 și MMP-). 9). Degradarea matricei extracelulare tumorale de către aceste enzime este o posibilă cauză a metastazelor și invaziei cancerului miometrului la ganglionii limfatici în CE de tip II. S-a demonstrat că curcumina suprimă calea de semnalizare ERK și scade expresia acestor enzime [ 21 ].
S-a raportat că curcumina induce apoptoza în celulele canceroase ovariane, prin calea independentă de P53, dar similar cu celulele P53 de tip sălbatic. Celulele HEY tratate cu curcumină au prezentat clivaj poli (ADP-riboză) polimerază-1, fragmentare ADN, fragmentare nucleară și condensare, indicând inducerea apoptozei celulare. Mai mult, curcumina este capabilă să induce apoptoza atât prin mecanisme intrinseci, cât și prin mecanisme extrinseci. Expresia regulatorilor antiapoptotici, supraviețuirea și Bcl-2, a fost scăzută în urma creșterii activității proteinei p38, protein kinazelor activate de mitogen (MAPK). Moartea celulelor anticanceroase cauzată de curcumină a fost raportată că are loc și prin suprimarea semnalizării Akt pentru supraviețuire în diferite celule de cancer ovarian [ 22 ]. Expresia activatorului plasminogenului de tip urokinază (uPA) a fost blocată de curcumină în linia celulară de cancer ovarian uman extrem de invazivă, HRA, care are un rol în metastaza cancerului. uPA este o serin protează exprimată prin căile Src-MAPK/ERK-AP-1 și Src-MAPK/ERK-PI3K/Akt-NF-kB ca răspuns la TGF-B1, unde curcumina inhibă formarea complexului AP-1 [ 22 ] . În consecință, curcumina poate îmbunătăți rezultatul în cancerele ginecologice avansate care împiedică invazia și progresia cancerului [ 23 ]. În secțiunile următoare, discutăm investigațiile recente privind terapia cu curcumină pentru cancerul endometrial, cervical și ovarian.
Curcumină și cancer endometrial
Cancerul endometrial (CE), cea mai frecventă tumoare malignă a tractului genital la femei, apare la femeile aflate în peri- și postmenopauză [ 24 , 25 ]. Se estimează că au fost diagnosticate peste 50.000 de cazuri noi, iar aproximativ 8.500 de cazuri au fost expirate din cauza acestei malignități în SUA în 2014. Recent, din cauza schimbărilor în stilul de viață, cancerele ginecologice provoacă mortalități în creștere [ 26 ] . Majoritatea persoanelor afectate sunt diagnosticate precoce și, de obicei, vindecate doar prin intervenție chirurgicală sau în combinație cu radioterapia [ 27 ]. Cu toate acestea, rata mortalității acestui cancer a cunoscut o creștere mai accentuată decât incidența sa în ultimele decenii [ 28 , 29 , 30 , 31 ]. În ciuda tuturor dezvoltării strategiilor terapeutice pentru această malignitate, pacienții în stadiu terminal se confruntă cu un prognostic prost, iar rata de supraviețuire la 5 ani este de până la 25-45% [26 ] . În plus, abordările terapeutice actuale nu sunt eficiente pentru 15% dintre pacienții care prezintă fenotip agresiv [ 27 ].
S-a raportat că într-o investigație in vivo de 30 de zile , administrarea zilnică intraperitoneală a 50 mg/kg curcumină a scăzut de cinci ori volumul tumorii în comparație cu cel la animalele tratate cu vehicul [ 32 ]. Sirohi și colegii săi au efectuat, de asemenea, experimente in vitro utilizând atât celulele HEC-1B, cât și celulele Ishikawa și au indicat că curcumina își prezintă efectele anti-migratorii prin creșterea expresiei Slit2 [ 32 ]. Inducerea Slit2 a reglat în jos expresia proteinelor migratoare, inclusiv metaloproteazele matriceale MMP2/9, factorul 1 derivat din celulele stromale (SDF-1) și CXCR4 (receptorul de chemokine 4) în celulele EC [32 ] . Acest studiu a indicat că curcumina suprimă creșterea tumorii și inhibă proliferarea celulelor EC [ 32 ]. De asemenea, mediază apoptoza indusă de ROS și prezintă efecte inhibitoare asupra migrării celulelor Hec-1B și Ishikawa prin reglarea descendentă indusă de Slit-2 a MMP-2 și -9, SDF-1 și CXCR4 [32 ] .
Receptorii de androgeni (AR) sunt factori de transcripție nucleari dependenți de ligand care au fost legați de diferite tipuri de tumori, inclusiv tumori endometriale, hepatice, vezicii urinare și de prostată [33 ] . S-a demonstrat că curcumina, dependent de doză și timp, crește apoptoza și blochează proliferarea liniei celulare EC RL-952 [ 34 ]. Curcumina a vizat, de asemenea, calea Wnt, care joacă un rol crucial în proliferarea și progresia tumorii și, ulterior, a declinat expresia AR în celulele canceroase [ 34 ]. Într-o cercetare efectuată de Chen și colab. [ 21 ], s-a demonstrat că curcumina inhibă invazia și migrarea celulelor de carcinom endometrial. În plus, a scăzut activitatea proteinazei și expresia metaloproteinazei matricei (MMP)-2 și -9 [ 21 ]. Curcumina în combinație cu inhibitorul ERK U0126 a redus sinergic expresia MMP-2/-9, conducând la inhibarea invaziei celulelor canceroase [ 35 ]. Sun și colab. a ilustrat că, pe lângă suprimarea invaziei celulelor EC prin reglarea în jos a MMP-2, curcumina inhibă și proliferarea lor [ 35 ]. Feng şi colab. a dezvăluit că curcumina suprimă apoptoza și proliferarea celulelor de carcinom endometrial uman prin reglarea în jos a expresiei AR prin calea Wnt [ 34 ]. Tabelul 1 prezintă datele disponibile despre efectele curcuminei în terapia EC.Tabelul 1 Studii efectuate asupra curcuminei și terapiei EC
Cancerul de col uterin este a treia cea mai răspândită malignitate și a patra cauză principală de mortalitate legată de cancer la femei [ 44 ]. În zilele noastre, incidența s-a declanșează femelele mai tinere [ 45 ]. Pacienții care sunt încă în stadii incipiente au un prognostic bun și răspund cu succes la tratamentele convenționale, inclusiv chimioradierea și/sau intervenția chirurgicală [ 46 ]. În timp ce aceste terapii standard ar putea provoca leziuni grave ale funcțiilor vaginale și ovariene [ 47 ], pacienții cu angajare extrapelvină au o rată de supraviețuire la 5 ani de 17%, iar la femeile cu boală recurentă această rată este mai mică de 5% [48 ] . Infecția cu virusul papiloma uman (HPV) este pasul inițial în majoritatea cazurilor de cancer de col uterin [ 49 ]. Pe lângă infecția cronică cu HPV, carcinogenul de fum (benzo[a]piren) și fumatul de țigară sunt considerați factori de risc majori legați de cancerul de col uterin [ 50 , 51 ].
Curcumina a evidențiat efecte chimioterapeutice și chimiopreventive dependente de concentrație într-un număr de investigații [ 52 ]. Curcumina exercită efecte citotoxice în celulele cancerului de col uterin într-o abordare dependentă de timp și doză, în special în celulele infectate cu HPV [ 53 ]. S-a confirmat că, prin suprimarea selectivă a activității proteinei activatoare 1 (AP-1), curcumina reglează în jos transcripția HPV18, care inversează dinamica expresiei fra-1 și c-fos în celulele cancerului de col uterin [53 ] . Funcția de supresie mai mare a curcuminei împotriva cancerului de col uterin se datorează inhibării căii mitocondriale, activității iNOS, COX-2 și ciclinei D1 și căilor de semnalizare ERK și Ras, precum și acțiunii telomerazei [54 , 55 ] . În mod remarcabil, curcumina acționează prin țintirea mai multor căi de semnalizare, făcând ca proliferarea celulelor canceroase de col uterin să revină la normal. De asemenea, mediază modificări importante ale proteinelor asociate tumorilor corelate cu metabolismul celular, ciclul celular și carcinogenitatea în celulele HeLa [ 56 ].
El și colegii de muncă au raportat că atât terapia cu curcumină-fotodinamică (PDT) cât și terapia duală antiplachetă (DAPT), un blocant al receptorului Notch, sunt capabile să inducă apoptoza și să blocheze proliferarea celulelor Me180 de cancer de col uterin [57 ] . Mai mult, DAPT are efecte sinergice asupra curcuminei-PDT în tratamentul cancerului de col uterin, care este asociat în principal cu reglarea NF-κB și Notch-1 [57 ] . Ghasemi şi colab. a efectuat recent un studiu in vitro pentru a evalua mecanismele probabile legate de efectele anticanceroase ale curcuminei asupra liniei celulare de cancer de col uterin [ 58 ]. Ei au dezvăluit că curcumina suprimă proliferarea și invazia celulelor canceroase de col uterin prin afectarea căilor Wnt/β-catenina și NF-kB [ 58 ]. Shang și colegii au raportat că 13 μM de curcumină induce moartea celulelor în celulele HeLa prin inducerea deteriorării ADN-ului și a condensării cromatinei [ 59 ]. S-a demonstrat că curcumina în combinație cu ultrasunetele exercită efecte mai benefice împotriva celulelor canceroase de col uterin [ 60 ]. Carr şi colab. a indicat că această combinație a îmbunătățit apoptoza în celulele SiHa sau HeLa [ 60 ]. Curcumina singură a cauzat mai puțină necroză în comparație cu terapia combinată. Ei au arătat capacitatea ultrasunetelor de a crește eficacitatea curcuminei [ 60 ]. Recent, s-a raportat că curcumina crește nivelurile intracelulare de ROS în celulele canceroase de col uterin, dar nu și în celulele epiteliale sănătoase [ 61 ]. Acest efect duce la inhibarea stresului ER și la restabilirea parțială a viabilității în celulele canceroase tratate cu curcumină [ 61 ]. În mod colectiv, aceste observații arată că curcumina promovează apoptoza mediată de stresul ER în celulele canceroase de col uterin prin creșterea generării de ROS specific tipului de celule [ 61 ]. Conform raportului Yoysungnoen-Chintana, curcumina în doze mari (1.000 și 1.500 mg/kg) inhibă angiogeneza, precum și creșterea tumorii la șoarecii implantați cu CaSki, posibil induse prin reglarea în jos a expresiei EGFR, COX- 2 și VEGF [ 62 ]. Tabelul 2 prezintă constatările disponibile privind terapia cu curcumină pentru cancerul de col uterin in vivo și in vitro .Tabelul 2 Investigații efectuate privind tratamentul cancerului de col uterin cu curcumină
În lumea occidentală, cancerul ovarian duce la cea mai mare rată a mortalității dintre toate cancerele ginecologice. Persoanele afectate sunt de obicei diagnosticate prea târziu în stadii avansate, când cancerul s-a extins la suprafețele peritoneale [ 96 , 97 , 98 ]. Chirurgia este cea mai eficientă terapie pentru stadiile avansate și este continuată prin chimioterapie cu paclitaxel și carboplatin. După trei cicluri de chimioterapie, intervenția chirurgicală citoreductivă se face alternativ [ 96 , 97 , 98 ].
Curcumina reduce doza necesară de radiații și cisplatină în suprimarea creșterii celulelor canceroase ovariane rezistente la cisplatină [ 99 ]. Curcumina, în combinație cu cantități mici de cisplatină, promovează, de asemenea, apoptoza [ 99 ]. Scăderea rezistenței la cisplatină în celulele cancerului ovarian de către curcumină se datorează probabil prin reglarea transferului indus de vezicule extracelulare de miR-214 și 3 exprimat matern (MEG3) [ 100 ]. Acest polifenol are efecte citotoxice împotriva celulelor OVCAR-3 rezistente la platină, care pot fi îmbunătățite semnificativ de compusul Y15 (tetraclorhidrat de 1, 2, 4, 5-benzen tetraamină) [101 ] . Mai mult, scade numărul și dimensiunea tumorilor ovariene prin inhibarea semnalizării STAT3 și NF-κB și inducerea căii factorului nuclear eritroid 2/hemoxigenază1 (Nrf2/HO-1) [102 ] . În plus, curcumina induce oprirea ciclului celular G2/M în celulele canceroase ovariane rezistente la cisplatină prin creșterea apoptozei și fosforilării p53 prin activarea caspazei-3 urmată de degradarea poli (ADP-riboză) polimerază-1 (PARP) [103 ] .
În liniile celulare de cancer ovarian uman A2780 și SK-OV-3, curcumina este capabilă să induce apoptoza, precum și autofagia protectoare prin suprimarea căii AKT/mTOR/p70S6K, demonstrând impactul sinergic al curcuminei și suprimarea autofagiei [ 104 ]. Curcumina inhibă activitatea ATPazei de calciu Sarco/reticulului endoplasmatic (SERCA) conducând la dereglarea homeostaziei Ca 2+ ; și, prin urmare, contribuie la apoptoza în celulele canceroase ovariane [ 105 ]. După cum sa discutat anterior, curcumina are efecte anti-proliferative și poate limita creșterea tumorii cu acest potențial. Shi și colegii săi au demonstrat, pe lângă inducerea apoptozei, 40 μM de curcumină a suprimat creșterea celulelor canceroase ovariane [ 106 ]. Lin şi colab. a arătat că, pe lângă inducerea apoptozei și efectele anti-proliferative, curcumina suprimă angiogeneza în cancerul ovarian in vivo și in vitro [ 107 ]. Mai mult, curcumina este capabilă să suprima reglarea în sus a Aquaporinei 3 mediată de factorul de creștere endotelial (EGF) și migrarea celulelor în celulele canceroase ovariane CaOV3 prin impactul său inhibitor asupra activării EGFR și AKT/ERK [108 ] . Într-o investigație in vitro, Xiaoling și colab. a raportat că curcumina inhibă metastaza și invazia celulelor umane de cancer ovarian SKOV3 prin suprimarea expresiei CXCR4 și CXCL-12 [ 109 ]. Wahl și colegii săi au ilustrat că tratamentul combinat al ligandului care induce apoptoza legată de Apo2 (Apo2L)/TNF și curcumină (5-15 μM) duce la creșterea inducerii morții celulelor apoptotice [ 110 ]. Ei au mai afirmat că, deoarece combinația menționată este capabilă să activeze atât căile de apoptoză intrinsecă, cât și extrinsecă, pot depăși chimiorezistența la medicamentele chimioterapeutice convenționale [ 110 ]. Sferoizii cancerului ovarian epitelial (EOC) au un rol cheie în dezvoltarea chimiorezistenței [ 111 ]. A fost efectuată o cercetare pentru a evalua efectele curcuminei asupra chimiorezistenței și metastazelor antiperitoneale la sferoizii EOC [ 112 ]. S-a indicat că celulele EOC invazive care formează sferoizi exprimă un nivel ridicat de aldehidă dehidrogenază 1 membru al familiei A1, un marker de celule stem canceroase, care a fost reglat în mod semnificativ în jos de curcumină [112 ] . Curcumina a crescut semnificativ sensibilitatea sferoizilor EOC la cisplatină și a abolit capacitatea lor de formare a sferei [ 112 ]. În plus, curcumina a inhibat creșterea sferoizilor EOC preexistenți și, de asemenea, a suprimat invazia lor la monostraturile mezoteliale și aderența lor la matricea extracelulară [ 112 ]. Tabelul 3rezumă datele actuale privind efectele terapeutice ale curcuminei asupra cancerului ovarian in vivo și in vitro .Tabelul 3 Studii recente privind tratamentul cu curcumina pentru cancerul ovarian
Efectele clinice anticancer ale curcuminei nu au fost încă pe deplin documentate, în ciuda efectelor sale potențiale. Solubilitatea slabă în apă este una dintre principalele restricții în aplicarea curcuminei (doar aproximativ 11 ng/ml), limitând astfel posibil efectele sale benefice [ 150 ]. Alte dezavantaje sunt instabilitatea în medii alcaline și neutre și biodisponibilitatea scăzută din cauza metabolismului și eliminării rapide [ 151 , 152 ].
Administrarea intravenoasă, peritoneală și orală a curcuminei duce la formarea metaboliților glucuronidului în ficat și la excreția prin bilă în tractul gastrointestinal [ 153 , 154 ]. Într-un studiu, administrarea zilnică de curcumină (3600 mg) la pacienții cu cancer colorectal metastatic, conținutul de curcumină în circulație a fost la nivel nanomolar [ 155 ]. În mod similar, alte constatări au arătat modificări minore în sângele periferic al pacienților cu risc crescut în urma administrării zilnice de curcumină (8000 mg) [ 156 ]. După administrarea curcuminei în doze de 500-8000 mg, nu a existat un nivel măsurabil în sânge și doar o cantitate mică din metaboliții săi a fost măsurată în unitățile de cercetare care au luat 10.000-12.000 mg [157 , 158 ] . Se poate concluziona că potențialele terapeutice ale curcuminei pot fi îmbunătățite prin îmbunătățirea biodisponibilității și solubilității acesteia.
Formulări noi de curcumină în tratamentul cancerelor ginecologice
După cum sa menționat anterior, curcumina are o absorbție scăzută și o biodisponibilitate slabă [ 159 ]; astfel, mai mulți investigatori au încercat să-și îmbunătățească eficacitatea terapeutică și profilul farmacocinetic prin abordări precum dezvoltarea de noi analogi [ 150 , 160 , 161 ]. Modificările structurale ale moleculei de curcumină conduc la numeroase proprietăți benefice pentru tratarea diferitelor boli, inclusiv boli neurodegenerative, diabet și boli cardiovasculare [ 159 ]. Curcumina și analogii săi sunt utilizați pe scară largă ca agenți antioxidanți, antimicrobieni, antiinflamatori și anticancer. Au fost raportate încercări de succes de sintetizare a unor noi analogi ai curcuminei cu bioactivitate îmbunătățită [ 3 , 162 , 163 ]. S-a indicat că curcumina are funcții anticancerigene datorită impactului său asupra diferitelor căi biologice implicate în metastaze, apoptoză, reglarea ciclului celular, tumorigeneză, exprimarea oncogenei și mutageneza [164 , 165 ] . Recent, a fost dezvăluit că unii dintre acești analogi prezintă proprietăți anticancerigene considerabil mai puternice decât curcumina [ 50 , 51 , 52 , 53 ]. Prin urmare, pentru a depăși restricțiile curcuminei, cercetătorii dezvoltă și evaluează un număr tot mai mare de analogi noi de curcumină [ 166 , 167 , 168 ]. Acești analogi au arătat un spectru larg de caracteristici anticanceroase în diferite celule canceroase, cum ar fi colon [ 169 ], sân [ 170 ] și prostată [ 171 ].
Pan și colegii au indicat că, în plasma animalelor tratate cu curcumină, 99% din curcumina administrată este conjugată cu glucuronid [ 172 ]. De asemenea, a fost dezvăluit că metaboliții redusi de curcumină, inclusiv glucuronozidele curcuminei, dihidrocurcumina și tetrahidrocurcumina sunt principalii metaboliți ai curcuminei [ 172 ]. S-a demonstrat că acești metaboliți au un rol crucial în multiple efecte terapeutice, inclusiv anticancer [ 173 ], antiinflamație [ 174 ] și antioxidare [ 175 ]. Cu toate acestea, numeroase investigații au demonstrat că activitatea anticanceroasă a metaboliților curcuminei este mai slabă în comparație cu molecula părinte [ 176 , 177 ]. Astfel, au fost testate mai multe abordări, cum ar fi modificarea structurală și improvizarea moleculelor purtătoare pentru a compensa limitările de biodisponibilitate ale curcuminei [ 176 ]. Pentru creșterea potențialului terapeutic și a biodisponibilității, diferite sisteme de livrare, inclusiv complexe fosfolipide, micele, nanoparticule și lipozomi au fost recomandate recent pentru tratamentul cu curcumină în cancer [ 176 ]. De exemplu, complexele fosfolipide și miceliile îmbunătățesc absorbția gastrointestinală a curcuminei, ducând la concentrații plasmatice mai mari [ 176 ]. S-a dezvăluit că miceliile mixte amfifile încărcate cu curcumină au o biodisponibilitate mai bună, provoacă o acumulare remarcabilă de rodamină și modulează nivelurile de expresie ale IL-6, IL-10 și TNF-α [41 ] . De asemenea, a indus apoptoza și a îmbunătățit absorbția intracelulară conducând la inhibarea creșterii celulelor EC Ishikawa [ 41 ]. Xu și colab. a efectuat o cercetare pentru a evalua eficacitatea curcuminei lipozomale ca tratament pentru cancerul endometrial [ 178 ]. Curcumina lipozomală în funcție de doză a suprimat motilitatea, a indus apoptoza și a inhibat proliferarea celulelor HEC-1 și Ishikawa [ 178 ]. De asemenea, a blocat expresia MMP-9, caspaza-3 și NF-kB. Este important că nu a fost observată nicio toxicitate în modelul de pește-zebră [ 178 ]. Demetoxicurcumina, un nou derivat al curcuminei, a indus apoptoza în celulele canceroase de col uterin in vitro și a scăzut volumul tumorii într-un model in vivo [ 91 ]. Într-o investigație, nanoparticulele de chitosan încărcate cu curcumină au fost utilizate pentru celulele canceroase de col uterin și s-a demonstrat că acest sistem de livrare a îmbunătățit efectele anti-proliferative ale medicamentului împotriva celulelor canceroase și nu a fost toxic pentru celulele normale [90] .]. Du şi colab. a raportat că demetoxicurcumina, un analog de curcumină, a inhibat substratul receptorului de insulină-2 (IRS2) prin reglarea miR-551a, ceea ce duce la împiedicarea creșterii celulelor canceroase ovariane [ 134 ]. În comparație cu curcumina liberă, purtătorii de lipide nanostructurați (NLC) încărcați cu curcumină au arătat rezultate anticancer mai bune și au redus mai eficient supraviețuirea coloniei celulare. Aceste descoperiri sugerează că captarea curcuminei în NLC-uri sporește eficacitatea curcuminei in vitro [ 137 ]. Hosseini şi colab. au demonstrat recent că nanocurcumina dendrozomală (DNC) duce la o moarte celulară mai mare atunci când este combinată cu oxaliplatina. DNC a indus, de asemenea, apoptoza în celulele canceroase ovariane [ 179 ]. De Matos și colab. a indicat că curcumină-nanoemulsia acționează ca un agent fotosensibilizant în PDT, demonstrând capacitatea alternativă potențială pentru leziunile cervicale [ 86 ]. Realizarea de noi formulări, cum ar fi derivații eficienți, precum și utilizarea sistemelor noi de administrare a medicamentelor pot crește semnificativ potențialul anticancer al curcuminei. Sunt necesare mai multe studii experimentale și studii umane pentru a confirma eficacitatea curcuminei și a formulărilor sale noi pentru tratamentul cancerelor ginecologice.
Concluzie și perspective de viitor
Cancerele ginecologice reprezintă o mare varietate de cancere, cum ar fi cancerul ovarian, uterin/endometrului, cervical, trofoblastic gestațional, peritoneal primar, vaginal și vulvar. Unele dintre aceste tipuri de cancer sunt cunoscute ca „ucigaș tăcut”, deoarece pacientul nu este de obicei conștient de prezența tumorii, până când este prea târziu pentru a se vindeca. Pe de altă parte, abordările terapeutice actuale au limite serioase. Prin urmare, dezvoltarea de noi platforme terapeutice ar putea optimiza rezultatul tratamentului pentru aceste tipuri de cancer.
Curcumina este un compus natural bine-cunoscut, care posedă o gamă largă de efecte biologice, cum ar fi activități antiinflamatorii, anticancerigene și antioxidante. Mai multe studii au demonstrat beneficiul potențial al tratamentului cu curcumină atât în monoterapie, cât și în combinație cu medicamentele standard de chimioterapie. Efectele anticanceroase ale curcuminei în cancerele ginecologice se dovedesc a fi legate de activarea căilor autofagice și apoptotice și de inhibarea invaziei și a metastazelor în diferite tumori. Curcumina suprimă genele de reglare majore, cum ar fi NF-κB și țintele sale genice din aval, care joacă un rol semnificativ în controlul invaziei și metastazei celulelor canceroase. Interesant, s-a demonstrat că curcumina îmbunătățește eficacitatea terapiilor actuale prin sensibilizarea celulelor canceroase cu rezistență la chimioradioterapie, care a reprezentat un mare obstacol în tratamentul cancerului. Important, siguranța și tolerabilitatea curcuminei au fost demonstrate în studiile clinice.
Alături de diferite efecte benefice, tratamentul cu curcumină se confruntă cu unele limitări, cum ar fi biodisponibilitatea scăzută . Cu toate acestea, mai multe studii au propus noi formulări de curcumină pentru a depăși această problemă. De asemenea, utilizarea diferiților analogi și sisteme noi de livrare, cum ar fi nanoparticule, lipozomi și micelii, poate îmbunătăți în continuare efectele anticancer ale curcuminei și poate deschide un nou orizont în tratamentul cancerelor ginecologice.
Referințe
Li Y, Kong D, Wang Z, Sarkar FH. Reglarea microARN-urilor de către agenți naturali: un domeniu emergent în cercetarea chimioprevenției și chimioterapiei. Farm Res. 2010;27(6):1027–41.ArticolCASGoogle Academic
Reuter S, Gupta SC, Park B, Goel A, Aggarwal BB. Modificări epigenetice induse de curcumină și alți compuși naturali. Genes Nutr. 2011;6(2):93–108.ArticolCASGoogle Academic
Vallianou NG, Evangelopoulos A, Schizas N, Kazazis C. Potențiale proprietăți anticanceroase și mecanisme de acțiune ale curcuminei. Anticancer Res. 2015;35(2):645–51.CASGoogle Academic
Teymouri M, Pirro M, Johnston TP, Sahebkar A. Curcumina ca un compus cu mai multe fațete împotriva infecției cu virusul papiloma uman și a cancerelor de col uterin: o revizuire a caracteristicilor chimice, celulare, moleculare și preclinice. BioFactors. 2017;43(3):331–46.ArticolCASGoogle Academic
Miriyala S, Panchatcharam M, Rengarajulu P. Cardioprotective effects of curcumin. Adv Exp Med Biol. 2007;595:359–77.ArticolGoogle Academic
Um MY, Hwang KH, Choi WH, Ahn J, Jung CH, Ha TY. Curcumina atenuează moleculele de adeziune și expresia metaloproteinazei matriceale la iepurii hipercolesterolemici. Nutr Res (New York, NY). 2014;34(10):886–93.ArticolCASGoogle Academic
Moghadamtousi SZ, Kadir HA, Hassandarvish P, Tajik H, Abubakar S, Zandi K. O revizuire a activității antibacteriene, antivirale și antifungice a curcuminei. BioMed Res Int. 2014;2014:186864.Google Academic
Sahebkar A, Serban MC, Ursoniu S, Banach M. Efectul curcuminoidelor asupra stresului oxidativ: O revizuire sistematică și meta-analiză a studiilor controlate randomizate. J Alimente funcționale. 2015;18:898–909.ArticolCASGoogle Academic
Panahi Y, Hosseini MS, Khalili N, Naimi E, Simental-Mendia LE, Majeed M, Sahebkar A. Efectele curcuminei asupra concentrațiilor serice de citokine la subiecții cu sindrom metabolic: O analiză post-hoc a unui studiu controlat randomizat. Biomed Pharmacother. 2016;82:578–82.ArticolCASGoogle Academic
Abdollahi E, Momtazi AA, Johnston TP, Sahebkar A. Efectele terapeutice ale curcuminei în bolile inflamatorii și mediate imun: un joc de natură? J Cell Physiol. 2018;233(2):830–48.ArticolCASGoogle Academic
Mollazadeh H, Cicero AFG, Blesso CN, Pirro M, Majeed M, Sahebkar A. Immune modulation by curcumin: The role of interleukin-10. Crit Rev Food Sci Nutr. 2019;59(1):89–101.ArticolCASGoogle Academic
Momtazi AA, Derosa G, Maffioli P, Banach M, Sahebkar A. Rolul microARN-urilor în efectele terapeutice ale curcuminei în bolile non-canceroase. Mol Diagn Ther. 2016;20(4):335–45.ArticolCASGoogle Academic
Gupta SC, Patchva S, Aggarwal BB. Rolurile terapeutice ale curcuminei: lecții învățate din studiile clinice. AAPS J. 2013;15(1):195–218.ArticolCASGoogle Academic
Soleimani V, Sahebkar A, Hosseinzadeh H. Turmericul (Curcuma longa) și constituentul său principal (curcumină) ca substanțe netoxice și sigure: revizuire. Phytother Res. 2018;32(6):985–95.ArticolCASGoogle Academic
Ferlay J, Colombet M, Soerjomataram I, Mathers C, Parkin DM, Pineros M, Znaor A, Bray F. Estimarea incidenței și mortalității globale a cancerului în 2018: surse și metode GLOBOCAN. Int J Cancer. 2019;144(8):1941–53.ArticolCASGoogle Academic
Suh DH, Kim M, Lee KH, Eom KY, Kjeldsen MK, Mirza MR, Kim JW. Progrese majore ale cercetării clinice în cancerul ginecologic în 2017. J Gynecol Oncol. 2018;29(2):e31.ArticolCASGoogle Academic
Kumar G, Mittal S, Sak K, Tuli HS. Mecanismele moleculare care stau la baza potențialului chimiopreventiv al curcuminei: provocări actuale și perspective viitoare. Life Sci. 2016;148:313–28.ArticolCASGoogle Academic
Zaman MS, Chauhan N, Yallapu MM, Gara RK, Maher DM, Kumari S, Sikander M, Khan S, Zafar N, Jaggi M și colab. Nanoformulare de curcumină pentru tratamentul cancerului de col uterin. Sci Rep. 2016;6:20051.ArticolCASGoogle Academic
Maruthur NM, Bolen SD, Brancati FL, Clark JM. Asocierea obezității și screening-ul cancerului de col uterin: o revizuire sistematică și meta-analiză. Obezitatea (Silver Spring, Maryland). 2009;17(2):375–81.ArticolGoogle Academic
Chen Q, Gao Q, Chen K, Wang Y, Chen L, Li XU. Curcumina suprimă migrarea și invazia celulelor de carcinom endometrial uman. Oncol Lett. 2015;10(3):1297–302.ArticolCASGoogle Academic
Watson JL, Greenshields A, Hill R, Hilchie A, Lee PW, Giacomantonio CA, Hoskin DW. Apoptoza indusă de curcumină în celulele de carcinom ovarian este independentă de p53 și implică activarea proteinei kinazei activate de mitogen p38 și reglarea în jos a expresiei Bcl-2 și a supraviețuinței și semnalizarea Akt. Mol Carcinog. 2010;49(1):13–24.ArticolCASGoogle Academic
Momtazi-Borojeni AA, Mosafer J, Nikfar B, Ekhlasi-Hundrieser M, Chaichian S, Mehdizadehkashi A, Vaezi A. Curcumin în Advancing Treatment for Gynecological Cancers with Developed Drug- and Radiotherapy-Associated Resistance. Rev Physiol Biochem Pharmacol. 2019;176:107–29.ArticolCASGoogle Academic
Kajo K, Vallova M, Biro C, Bognar G, Machalekova K, Zavodna K, Galbavy S, Zubor P. Molecular pathology of endometrial carcinom – a review. Cesk Patol. 2015;51(2):65–73.Google Academic
Buhtoiarova TN, Brenner CA, Singh M. Carcinomul endometrial: Rolul biomarkerilor actuali și emergenti în rezolvarea dilemelor clinice persistente. Am J Clin Pathol. 2016;145(1):8–21.ArticolCASGoogle Academic
Chen S, Sun KX, Liu BL, Zong ZH, Zhao Y. Rolul glicogen sintazei kinazei-3beta (GSK-3beta) în carcinomul endometrial: un marker de carcinogeneză, progresie, prognostic și terapie țintă. Oncotarget. 2016;7(19):27538–51.ArticolGoogle Academic
Yeramian A, Garcia V, Bergada L, Domingo M, Santacana M, Valls J, Martinez-Alonso M, Carceller JA, Cussac AL, Dolcet X, et al. Imagistica de bioluminiscență pentru a monitoriza efectele inhibitorului Hsp90 NVP-AUY922 asupra căii NF-kappaB în cancerul endometrial. Mol Imaging Biol. 2016;18(4):545–56.ArticolCASGoogle Academic
Fuso L, Evangelista A, Pagano E, Piovano E, Perotto S, Mazzola S, Bertoldo E, La Porta MR, Rosmino C, Furbatto G, et al. Variație în practica de urmărire a oncologiei ginecologice: atribuită centrelor de cancer sau caracteristicilor pacientului? Un studiu al rețelei regionale de oncologie din Piemont. Tumori. 2011;97(5):551–8.ArticolGoogle Academic
Duong LM, Wilson RJ, Ajani UA, Singh SD, Eheman CR. Tendințe în ratele de incidență a cancerului endometrial în Statele Unite, 1999-2006. J Sănătatea femeilor (2002). 2011;20(8):1157–63.ArticolGoogle Academic
Dewdney SB, Kizer NT, Andaya AA, Babb SA, Luo J, Mutch DG, Schmidt AP, Brinton LA, Broaddus RR, Ramirez NC și colab. Carcinom seros uterin: risc familial crescut pentru tumori maligne asociate linșului. Cancer Prev Res (Philadelphia, Pa). 2012;5(3):435–43.ArticolGoogle Academic
Ueda SM, Kapp DS, Cheung MK, Shin JY, Osann K, Husain A, Teng NN, Berek JS, Chan JK. Tendințe ale caracteristicilor demografice și clinice la femeile diagnosticate cu cancer de corpus și impactul potențial al acestora asupra numărului tot mai mare de decese. Am J Obstet Gynecol. 2008;198(2):218.e211–6.ArticolGoogle Academic
Sirohi VK, Popli P, Sankhwar P, Kaushal JB, Gupta K, Manohar M, Dwivedi A. Curcumina prezintă efect anti-tumoral și atenuează migrarea celulară prin reglarea în jos mediată de Slit-2 a SDF-1 și CXCR4 în celulele de adenocarcinom endometrial. J Nutr Biochem. 2017;44:60–70.ArticolCASGoogle Academic
Davey RA, Grossmann M. Structura receptorului de androgeni, funcția și biologia: de la bancă până la pat. Clin Biochem Rev. 2016;37(1):3–15.Google Academic
Feng W, Yang CX, Zhang L, Fang Y, Yan M. Curcumina promovează apoptoza celulelor de carcinom endometrial uman prin reglarea în jos a expresiei receptorului de androgeni prin calea semnalului Wnt. Eur J Gynaecolog Oncol. 2014;35(6):718–23.CASGoogle Academic
Sun MX, Yu F, Gong ML, Fan GL, Liu CX. Efectele curcuminei asupra rolului MMP-2 în proliferarea și invazia celulelor canceroase endometriale. Eur Rev Med Pharmacol Sci. 2018;22(15):5033–41.Google Academic
Saydmohammed M, Joseph D, Syed V. Curcumina suprimă activarea constitutivă a STAT-3 prin suprareglarea inhibitorului proteic al STAT-3 activat (PIAS-3) în celulele canceroase ovariane și endometriale. J Cell Biochim. 2010a;110(2):447–56.CASGoogle Academic
Patel SK, Jackson L, Warren AY, Arya P, Shaw RW, Khan RN. Un rol pentru canalele de potasiu cu doi pori (K2P) în funcția epitelială endometrială. J Cell Mol Med. 2013;17(1):134–46.ArticolCASGoogle Academic
Liang YJ, Zhang HM, Wu YZ, Hao Q, Wang JD, Hu YL. Efectul inhibitor al letrozolului combinat cu curcumină asupra creșterii carcinomului endometrial xenogrefat la șoarecii nuzi. Chin J Cancer. 2010;29(1):9–14.ArticolCASGoogle Academic
Yu Z, Shah DM. Curcumina reglează în jos expresia Ets-1 și Bcl-2 în celulele HEC-1-A ale carcinomului endometrial uman. Ginecol Oncol. 2007;106(3):541–8.ArticolCASGoogle Academic
Pereira MC, Mohammed R, VAN Otterlo WAL, DE Koning CB, Davids H. Evaluarea efectelor derivaților de aminonaftochinonă în combinație cu curcumină împotriva cancerului de sân ER-pozitiv și a tumorilor asociate. Anticancer Res. 2017;37(12):6749–59.CASGoogle Academic
Kumar A, Sirohi VK, Anum F, Singh PK, Gupta K, Gupta D, Saraf SA, Dwivedi A, Chourasia MK. Apoptoză îmbunătățită, reglare în jos a supraviețuirii și imunochimioterapie asistată prin micelii mixte amfifile încărcate cu curcumină pentru subjugarea cancerului endometrial. Nanomedicina. 2017;13(6):1953–63.ArticolCASGoogle Academic
Xu H, Gong Z, Zhou S, Yang S, Wang D, Chen X, Wu J, Liu L, Zhong S, Zhao J și colab. Curcumina lipozomală care vizează cancerul endometrial prin calea NF-kappaB. Cell Physiol Biochim. 2018;48(2):569–82.ArticolCASGoogle Academic
Tuyaerts S, Rombauts K, Everaert T, Van Nuffel AMT, Amant F. Un studiu de fază 2 pentru a evalua capacitatea imunomodulatoare a unui sistem de livrare bazat pe lecitină a curcuminei în cancerul endometrial. Nutră față. 2018;5:138.ArticolCASGoogle Academic
Wu ES, Jeronimo J, Feldman S. Bariere și provocări ale alternativelor de tratament pentru cancerul de col uterin în stadiu incipient în setări cu resurse inferioare. J Global Oncol. 2017;3(5):572–82.ArticolGoogle Academic
Kessler TA. Cancerul de col uterin: prevenire și depistare precoce. Semin Oncol Nurs. 2017;33(2):172–83.ArticolGoogle Academic
Marth C, Landoni F, Mahner S, McCormack M, Gonzalez-Martin A, Colombo N. Cancerul de col uterin: Ghidurile de practică clinică ESMO pentru diagnostic, tratament și urmărire. Ann Oncol. 2017;28(suppl_4):iv72–83.ArticolCASGoogle Academic
LaVigne AW, Triedman SA, Randall TC, Trimble EL, Viswanathan AN. Cancerul de col uterin în țările cu venituri mici și medii: abordarea barierelor în calea administrării radioterapiei. Gynecol Oncol Rep. 2017;22:16–20.ArticolGoogle Academic
Horner M, Ries L, Krapcho M, Neyman N, Aminou R, Howlader N, Altekruse S, Feuer E, Huang L, Mariotto A. SEER Cancer Statistics Review, 1975-2013, voi. 19. Bethesda: Institutul Național al Cancerului; 2013.Google Academic
zur Hausen H. Papilomavirusuri în cauzarea cancerelor umane – o scurtă relatare istorică. Virologie. 2009;384(2):260–5.ArticolCASGoogle Academic
Maher DM, Bell MC, O’Donnell EA, Gupta BK, Jaggi M, Chauhan SC. Curcumina suprimă oncoproteinele papilomavirusului uman, restabilește proteinele p53, Rb și PTPN13 și inhibă reglarea pozitivă indusă de benzo[a]piren a HPV E7. Mol Carcinog. 2011;50(1):47–57.ArticolCASGoogle Academic
Moore EE, Wark JD, Hopper JL, Erbas B, Garland SM. Rolurile factorilor genetici și de mediu asupra riscului de cancer de col uterin: o revizuire a studiilor clasice pe gemeni. Twin Res Hum Genet. 2012;15(1):79–86.ArticolGoogle Academic
Nardo L, Andreoni A, Bondani M, Masson M, Hjorth Tonnesen H. Studies on curcumin and curcuminoids. XXXIV. Proprietăți fotofizice ale unui analog de curcumină simetric, nesubstituit. J Photochem Photobiol B. 2009;97(2):77–86.ArticolCASGoogle Academic
Prusty BK, Das BC. Activarea constitutivă a factorului de transcripție AP-1 în cancerul de col uterin și suprimarea transcripției papilomavirusului uman (HPV) și a activității AP-1 în celulele HeLa de către curcumină. Int J Cancer. 2005;113(6):951–60.ArticolCASGoogle Academic
Singh M, Singh N. Curcumina contracarează efectul proliferativ al estradiolului și induce apoptoza în celulele canceroase de col uterin. Mol Cell Biochim. 2011;347(1-2):1–11.ArticolCASGoogle Academic
Di Domenico F, Foppoli C, Coccia R, Perluigi M. Antioxidants in cervical cancer: chemopreventive and chemotherapeutic effects of polyphenols. Biochim Biophys Acta. 2012;1822(5):737–47.ArticolCASGoogle Academic
Madden K, Flowers L, Salani R, Horowitz I, Logan S, Kowalski K, Xie J, Mohammed SI. Abordare bazată pe proteomică pentru a elucida mecanismul efectului antitumoral al curcuminei în cancerul de col uterin. Prostaglandine Leukot Essenti acizi grași. 2009;80(1):9–18.ArticolCASGoogle Academic
He G, Mu T, Yuan Y, Yang W, Zhang Y, Chen Q, Bian M, Pan Y, Xiang Q, Chen Z și colab. Efectele căii de semnalizare Notch în cancerul de col uterin prin terapia fotodinamică mediată de curcumină și mecanismele sale posibile in vitro și in vivo. J Cancer. 2019;10(17):4114–22.ArticolCASGoogle Academic
Ghasemi F, Shafiee M, Banikazemi Z, Pourhanifeh MH, Khanbabaei H, Shamshirian A, Moghadam SA, ArefNezhad R, Sahebkar A, Avan A. Curcumina inhibă căile NF-kB și Wnt/β-catenina în celulele cancerului de col uterin. Pathol Res Pract. 2019;215(10):152556.ArticolCASGoogle Academic
Shang HS, Chang CH, Chou YR, Yeh MY, Au MK, Lu HF, Chu YL, Chou HM, Chou HC, Shih YL. Curcumina provoacă leziuni ale ADN-ului și afectează expresia proteinelor asociate în celulele umane de cancer de col uterin HeLa. Oncol Rep. 2016;36(4):2207–15.ArticolCASGoogle Academic
Carr KR, Ioffe YJ, Filippova M, Duerksen-Hughes P, Chan PJ. Tratamentul combinat cu ultrasunete-curcumină al celulelor canceroase de col uterin uman. Eur J Obstet Gynecol Reprod Biol. 2015;193:96–101.ArticolGoogle Academic
Kim B, Kim HS, Jung EJ, Lee JY, Tsang BK, Lim JM, Song YS. Curcumina induce apoptoza mediată de stresul ER prin generarea selectivă de specii reactive de oxigen în celulele cancerului de col uterin. Mol Carcinog. 2016;55(5):918–28.ArticolCASGoogle Academic
Yoysungnoen-Chintana P, Bhattarakosol P, Patumraj S. Activitățile antitumorale și antiangiogenice ale curcuminei în xenogrefele de cancer de col uterin la șoarecii nuzi. Biomed Res Int. 2014;2014:817972.ArticolGoogle Academic
Aedo-Aguilera V, Carrillo-Beltran D, Calaf GM, Munoz JP, Guerrero N, Osorio JC, Tapia JC, Leon O, Contreras HR, Aguayo F. Curcumina scade tranziția epitelialmezenchimală printr-un mecanism Pirindependent în celulele cancerului de col uterin. Oncol Rep. 2019;42(5):2139–48.CASGoogle Academic
Ghasemi F, Shafiee M, Banikazemi Z, Pourhanifeh MH, Khanbabaei H, Shamshirian A, Amiri Moghadam S, ArefNezhad R, Sahebkar A, Avan A și colab. Curcumina inhibă căile NF-kB și Wnt/beta-cateninei în celulele canceroase de col uterin. Pathol Res Pract. 2019;215(10):152556.ArticolCASGoogle Academic
Kumar D, Basu S, Parija L, Rout D, Manna S, Dandapat J, Debata PR. Curcumina și acidul elagic induc sinergic generarea de ROS, deteriorarea ADN-ului, acumularea p53 și apoptoza în celulele de carcinom cervical HeLa. Biomed Pharmacother. 2016;81:31–7.ArticolCASGoogle Academic
Lee JW, Park S, Kim SY, Um SH, Moon EY. Curcumina împiedică efectul antitumoral al vinblastinei prin inhibarea dinamicii microtubulilor și a potențialului membranei mitocondriale în celulele canceroase de col uterin HeLa. Fitomedicina. 2016;23(7):705–13.ArticolCASGoogle Academic
Thacker PC, Karunagaran D. Curcumin și emodin reduc calea de semnalizare a TGF-beta în celulele cancerului de col uterin uman. Plus unu. 2015;10(3):e0120045.ArticolCASGoogle Academic
Dang YP, Yuan XY, Tian R, Li DG, Liu W. Curcumin îmbunătățește apoptoza indusă de paclitaxel a celulelor canceroase de col uterin umane HPV-pozitive prin calea NF-kappaB-p53-caspaze-3. Exp Ther Med. 2015;9(4):1470–6.ArticolCASGoogle Academic
Lewinska A, Adamczyk J, Pajak J, Stoklosa S, Kubis B, Pastuszek P, Slota E, Wnuk M. Scăderea mediată de curcumină a expresiei regiunilor organizatoare nucleolare în celulele cancerului de col uterin (HeLa). Mutat Res Genet Toxicol Environ Mutagen. 2014;771:43–52.ArticolCASGoogle Academic
Roy M, Mukherjee S. Inversarea rezistenței față de cisplatină de către curcumină în celulele cancerului de col uterin. Asian Pac J Cancer Prev. 2014;15(3):1403–10.ArticolGoogle Academic
Debata PR, Castellanos MR, Fata JE, Baggett S, Rajupet S, Szerszen A, Begum S, Mata A, Murty VV, Opitz LM, et al. O nouă cremă vaginală pe bază de curcumină, Vacurin, elimină selectiv celulele umane apuse de cancer de col uterin. Ginecol Oncol. 2013;129(1):145–53.ArticolCASGoogle Academic
Sreekanth CN, Bava SV, Sreekumar E, Anto RJ. Dovezi moleculare pentru eficacitatea chemosensibilizantă a curcuminei lipozomale în chimioterapia cu paclitaxel la modelele de șoarece de cancer de col uterin. Oncogene. 2011;30(28):3139–52.ArticolCASGoogle Academic
Singh M, Singh N. Mecanismul molecular al citotoxicității induse de curcumină în celulele de carcinom cervical uman. Mol Cell Biochim. 2009;325(1-2):107–19.ArticolCASGoogle Academic
Javvadi P, Segan AT, Tuttle SW, Koumenis C. Agentul chemopreventiv curcumina este un radiosensibilizator puternic al celulelor tumorale de col uterin uman prin creșterea producției de specii reactive de oxigen și supraactivarea căii proteine kinazei activate de mitogen. Mol Pharmacol. 2008;73(5):1491–501.ArticolCASGoogle Academic
Murugesan K, Koroth J, Srinivasan PP, Singh A, Mukundan S, Karki SS, Choudhary B, Gupta CM. Efectele nanoparticulelor de argint sintetizate verzi (ST06-AgNPs) folosind derivat de curcumină (ST06) asupra celulelor umane de cancer de col uterin (HeLa) in vitro și modele de șoareci cu tumori EAC. Int J Nanomedicine. 2019;14:5257–70.ArticolCASGoogle Academic
Wang WY, Cao YX, Zhou X, Wei B. Livrarea curcuminei lipozomale modificate cu acid folic pentru terapia țintită a carcinomului cervical. Drug Des Devel Ther. 2019;13:2205–13.ArticolGoogle Academic
Chaudhary M, Kumar N, Baldi A, Chandra R, Babu MA, Madan J. Analog 4-Bromo-4′-cloro pirazoline al curcuminei a crescut activitatea anticancer împotriva cancerului de col uterin uman, celule HeLa: în analiză, sinteză și silico-ghidată citotoxicitate in vitro. J Biomol Struct Dyn. 2020;38(5):1335–53.ArticolCASGoogle Academic
Chaudhary M, Kumar N, Baldi A, Chandra R, Arockia Babu M, Madan J. Chloro și bromo-pirazol curcumină Knoevenagel condensează activitatea anticanceroasă crescută împotriva celulelor canceroase de col uterin uman: proiectare, sinteza, andocare in silico și analiză de citotoxicitate in vitro. J Biomol Struct Dyn. 2020;38(1):200–18.ArticolCASGoogle Academic
Upadhyay A, Yagnik B, Desai P, Dalvi SV. Livrare îmbunătățită mediată de microbule de curcumină către celulele canceroase de col uterin. ACS omega. 2018;3(10):12824–31.ArticolCASGoogle Academic
Liao CL, Chu YL, Lin HY, Chen CY, Hsu MJ, Liu KC, Lai KC, Huang AC, Chung JG. Bisdemetoxicurcumina suprimă migrarea și invazia celulelor HeLa ale cancerului de col uterin uman prin inhibarea căilor NF-kB, MMP-2 și -9. Anticancer Res. 2018;38(7):3989–97.ArticolCASGoogle Academic
He GF, Mu TL, Pan YS, Chen ZH, Xiang Q, Yang WY, Zhang Y, Yuan YL, Sun AP. Efectul inhibitor al DAPT asupra căii de semnalizare Notch în terapia fotodinamică mediată de curcumină pentru xenogrefele de cancer de col uterin la șoarecii nuzi. Zhonghua Yi Xue Za Zhi. 2018;98(19):1511–6.CASGoogle Academic
Kumari P, Rompicharla SVK, Muddineti OS, Ghosh B, Biswas S. Micelele pe bază de poli (lactidă) ancorate cu transferină pentru a îmbunătăți activitatea anticanceroasă a curcuminei în monostraturile celulelor canceroase hepatice și cervicale și sferoizii 3D. Int J Biolog Macromol. 2018;116:1196–213.ArticolCASGoogle Academic
Lin CC, Kuo CL, Huang YP, Chen CY, Hsu MJ, Chu YL, Chueh FS, Chung JG. Demetoxicurcumina suprimă migrarea și invazia celulelor HeLa ale cancerului de col uterin uman prin inhibarea căilor NF-kappaB. Anticancer Res. 2018;38(5):2761–9.CASGoogle Academic
Khan MA, Zafaryab M, Mehdi SH, Ahmad I, Rizvi MMA. Caracterizarea fizico-chimică a nanoparticulelor de chitosan încărcate cu curcumină: Implicații în cancerul de col uterin. Agenți anticancer Med Chem. 2018;18(8):1131–7.ArticolCASGoogle Academic
Gawde KA, Sau S, Tatiparti K, Kashaw SK, Mehrmohammadi M, Azmi AS, Iyer AK. Paclitaxel și curcumină di-fluorurată încărcate în nanoparticule de albumină pentru terapia combinată sinergică țintită a cancerului ovarian și de col uterin. Coloizii Surf B, Biointerfețe. 2018;167:8–19.ArticolCASGoogle Academic
de Matos RPA, Calmon MF, Amantino CF, Villa LL, Primo FL, Tedesco AC, Rahal P. Effect of Curcumin-Nanoemulsion Associated with Photodynamic Therapy in Cervical Carcinoma Cell Lines. Biomed Res Int. 2018;2018:4057959.ArticolCASGoogle Academic
Wang J, Liu Q, Yang L, Xia X, Zhu R, Chen S, Wang M, Cheng L, Wu X, Wang S. Curcumin-Loaded TPGS/F127/P123 Mixed Polymeric Micelles for Cervical Cancer Therapy: Formulation, Characterization, și Evaluare InVitro și InVivo. J Biomed Nanotechnol. 2017;13(12):1631–46.ArticolCASGoogle Academic
Ahmadi F, Ghasemi-Kasman M, Ghasemi S, Gholamitabar Tabari M, Pourbagher R, Kazemi S, Alinejad-Mir A. Inducerea apoptozei în celulele canceroase HeLa printr-o sinteză mediată de ultrasunete a nanoparticulelor de chitosan-alginat-STPP încărcate cu curcumină. Int J Nanomedicine. 2017;12:8545–56.ArticolCASGoogle Academic
Luong D, Kesharwani P, Alsaab HO, Sau S, Padhye S, Sarkar FH, Iyer AK. Micelele polimerice conjugate cu acid folic încărcate cu un analog difluorinat de curcumină pentru țintirea cancerelor de col uterin și ovarian. Coloizii Surf B Biointerfețe. 2017;157:490–502.ArticolCASGoogle Academic
Khan MA, Zafaryab M, Mehdi SH, Ahmad I, Rizvi MM. Caracterizarea și activitatea anti-proliferativă a nanoparticulelor de chitosan încărcate cu curcumină în cancerul de col uterin. Int J Biolog Macromol. 2016;93(Pt A):242–53.ArticolCASGoogle Academic
Yoysungnoen B, Bhattarakosol P, Changtam C, Patumraj S. Efectele tetrahidrocurcuminei asupra creșterii tumorii și semnalizării celulare în xenogrefele de cancer de col uterin la șoareci nuzi. Biomed Res Int. 2016;2016:1781208.ArticolCASGoogle Academic
Yoysungnoen B, Bhattarakosol O, Changtam C, Patumraj S. Efectul tratamentului combinat al tetrahidrocurcuminei și celecoxibului asupra creșterii tumorii induse de celulele cancerului de col uterin și angiogenezei tumorii la șoareci nuzi. J Med Asoc Thai. 2016;99(Suppl 4):S23–31.Google Academic
Yoysungnoen B, Bhattarakosol P, Patumraj S, Changtam C. Efectele tetrahidrocurcuminei asupra factorului 1 alfa inductibil de hipoxie și expresiei factorului de creștere endotelial vascular în angiogeneza indusă de celulele cancerului de col uterin la șoarecii nuzi. Biomed Res Int. 2015;2015:391748.ArticolCASGoogle Academic
Zheng M, Liu S, Guan X, Xie Z. Sinteza într-un singur pas a cadrelor de imidazolat zeolitic la scară nanometrică cu încărcare mare de curcumină pentru tratamentul cancerului de col uterin. Interfețe ACS Appl Mater. 2015;7(40):22181–7.ArticolCASGoogle Academic
Saengkrit N, Saesoo S, Srinuanchai W, Phunpee S, Ruktanonchai UR. Influența lipozomului cationic încărcat cu curcumină asupra activității anticancer pentru terapia cancerului de col uterin. Coloizii Surf B Biointerfețe. 2014;114:349–56.ArticolCASGoogle Academic
Wright AA, Bohlke K, Armstrong DK, Bookman MA, Cliby WA, Coleman RL, Dizon DS, Kash JJ, Meyer LA, Moore KN. Chimioterapia neoadjuvantă pentru cancerul ovarian avansat nou diagnosticat: ghidul de practică clinică al Societății de Oncologie Ginecologică și al Societății Americane de Oncologie Clinică. Ginecol Oncol. 2016;143(1):3–15.ArticolGoogle Academic
Vergote I, Tropé CG, Amant F, Kristensen GB, Ehlen T, Johnson N, Verheijen RH, Van Der Burg ME, Lacave AJ, Panici PB. Chimioterapia neoadjuvantă sau intervenția chirurgicală primară în cancerul ovarian în stadiul IIIC sau IV. N Engl J Med. 2010;363(10):943–53.ArticolCASGoogle Academic
Chang SJ, Bristow RE, Chi DS, Cliby WA. Rolul citoreducției chirurgicale agresive în cancerul ovarian avansat. J Gynecol Oncol. 2015;26(4):336–42.ArticolCASGoogle Academic
Yallapu MM, Maher DM, Sundram V, Bell MC, Jaggi M, Chauhan SC. Curcumina induce chimio/radio-sensibilizare în celulele canceroase ovariane, iar nanoparticulele de curcumină inhibă creșterea celulelor canceroase ovariane. J Ovarian Res. 2010;3(1):11.ArticolCASGoogle Academic
Zhang J, Liu J, Xu X, Li L. Curcumina suprimă dezvoltarea rezistenței la cisplatină parțial prin modularea transferului extracelular mediat de vezicule de MEG3 și miR-214 în cancerul ovarian. Cancer Chemother Pharmacol. 2017;79(3):479–87.ArticolCASGoogle Academic
Levy AS, Rathinavelu A, Coelho N, Ramnot A, Kanagasabai T, Bracho O, Smith R. Evaluation of the efficacy of curcumin and Y15 in platinum resistant ovarian cancer cells. În: AACR; 2018.Google Academic
Sahin K, Orhan C, Tuzcu M, Sahin N, Tastan H, Özercan İH, Güler O, Kahraman N, Kucuk O, Ozpolat B. Eficacitatea chimiopreventivă și antitumorală a curcuminei într-un model de cancer ovarian de găină în dezvoltare spontană. Cancer Prev Res. 2018;11(1):59–67.ArticolCASGoogle Academic
Weir NM, Selvendiran K, Kutala VK, Tong L, Vishwanath S, Rajaram M, Tridandapani S, Anant S, Kuppusamy P. Curcumina induce stoparea G2/M și apoptoza în celulele cancerului ovarian uman rezistente la cisplatină prin modularea Akt și p38 MAPK. Cancer Biol Ther. 2007;6(2):178–84.ArticolCASGoogle Academic
Liu LD, Pang YX, Zhao XR, Li R, Jin CJ, Xue J, Dong RY, Liu PS. Curcumina induce moartea celulelor apoptotice și autofagia protectoare prin inhibarea căii AKT/mTOR/p70S6K în celulele canceroase ovariane umane. Arch Gynecol Obstetr. 2019;299(6):1627–39.ArticolCASGoogle Academic
Seo JA, Kim B, Dhanasekaran DN, Tsang BK, Song YS. Curcumina induce apoptoza prin inhibarea activității Ca2+ ATPazei reticulului sarco/endoplasmatic în celulele canceroase ovariane. Cancer Lett. 2016;371(1):30–7.ArticolCASGoogle Academic
Shi M, Cai Q, Yao L, Mao Y, Ming Y, Ouyang G. Antiproliferarea și apoptoza induse de curcumină în celulele cancerului ovarian uman. Cell Biol Int. 2006;30(3):221–6.ArticolCASGoogle Academic
Lin YG, Kunnumakkara AB, Nair A, Merritt WM, Han LY, Armaiz-Pena GN, Kamat AA, Spannuth WA, Gershenson DM, Lutgendorf SK. Curcumina inhibă creșterea tumorii și angiogeneza în carcinomul ovarian prin țintirea căii factorului nuclear-κB. Clin Cancer Res. 2007;13(11):3423–30.ArticolCASGoogle Academic
Ji C, Cao C, Lu S, Kivlin R, Amaral A, Kouttab N, Yang H, Chu W, Bi Z, Di W. Curcumina atenuează reglarea AQP3 indusă de EGF și migrarea celulelor în celulele cancerului ovarian uman. Cancer Chemother Pharmacol. 2008;62(5):857–65.ArticolCASGoogle Academic
Xiaoling M, Jing Z, Fang X, Liangdan T. Curcumina inhibă invazia și metastaza în celulele canceroase ovariane umane SKOV3 de către axa CXCL12-CXCR4. Afr J Biotechnol. 2010;9(48):8230–4.ArticolGoogle Academic
Wahl H, Tan L, Griffith K, Choi M, Liu JR. Curcumina îmbunătățește apoptoza indusă de Apo2L/TRAIL în celulele canceroase ovariane chimiorezistente. Ginecol Oncol. 2007;105(1):104–12.ArticolCASGoogle Academic
Cornelison R, Llaneza DC, Landen CN. Terapeutice emergente pentru a depăși chimiorezistența în cancerul ovarian epitelial: o mini-revizuire. Int J Mol Sci. 2017;18(10):2171.ArticolCASGoogle Academic
He M, Wang D, Zou D, Wang C, Lopes-Bastos B, Jiang WG, Chester J, Zhou Q, Cai J. Reutilizarea curcuminei ca agent anti-metastatic pentru tratamentul cancerului ovarian epitelial: in vitro model folosind sferoizi de cancer ovarian îmbogățiți cu celule stem canceroase. Oncotarget. 2016;7(52):86374–87.ArticolGoogle Academic
Yen HY, Tsao CW, Lin YW, Kuo CC, Tsao CH, Liu CY. Reglarea carcinogenezei și modulării prin semnalizarea Wnt/beta-cateninei de către curcumină într-o linie celulară de cancer ovarian. Sci Rep. 2019;9(1):17267.ArticolCASGoogle Academic
Tian M, Tian D, Qiao X, Li J, Zhang L. Modularea semnalizării NF-kB -STAT3 indusă de Myb și rezistența la cisplatină rezultată în cancerul ovarian de către factorii dietetici. J Cell Physiol. 2019;234(11):21126–34.ArticolCASGoogle Academic
Kwon Y. Polifenolii derivați din alimente inhibă creșterea celulelor canceroase ovariane, indiferent de capacitatea acestora de a induce răspunsuri antioxidante. Heliyon. 2018;4(8):e00753.ArticolGoogle Academic
Choe SR, Kim YN, Park CG, Cho KH, Cho DY, Lee HY. RCP induce fosforilarea FAK și invazia celulelor canceroase ovariane cu inhibare de către curcumină. Exp Mol Med. 2018;50(4):52.ArticolCASGoogle Academic
Sahin K, Orhan C, Tuzcu M, Sahin N, Tastan H, Ozercan IH, Guler O, Kahraman N, Kucuk O, Ozpolat B. Chemopreventive and Antitumor Efficacy of Curcumin in a Spontaneously Developing Hen Ovarian Cancer Model. Cancer Prev Res (Philadelphia, Pa). 2018;11(1):59–67.ArticolCASGoogle Academic
Zhao J, Pan Y, Li X, Zhang X, Xue Y, Wang T, Zhao S, Hou Y. Dihidroartemisinina și curcumina induc sinergic apoptoza în celulele SKOV3 prin suprareglarea MiR-124 care vizează Midkine. Cell Physiol Biochim. 2017;43(2):589–601.ArticolCASGoogle Academic
Zhao SF, Zhang X, Zhang XJ, Shi XQ, Yu ZJ, Kan QC. Inducerea microARN-9 mediază citotoxicitatea curcuminei împotriva celulelor canceroase ovariane SKOV3. Asian Pac J Cancer Prev. 2014;15(8):3363–8.ArticolGoogle Academic
Lv J, Shao Q, Wang H, Shi H, Wang T, Gao W, Song B, Zheng G, Kong B, Qu X. Efectele și mecanismele curcuminei și polizaharidei de busuioc asupra invaziei celulelor SKOV3 și a celulelor dendritice. Mol Med Rep. 2013;8(5):1580–6.ArticolCASGoogle Academic
Seo JH, Jeong KJ, Oh WJ, Sul HJ, Sohn JS, Kim YK, Kang JK, Park CG, Lee HY. Acidul lizofosfatidic induce fosforilarea STAT3 și motilitatea celulelor canceroase ovariane: inhibarea acestora de către curcumină. Cancer Lett. 2010;288(1):50–6.ArticolCASGoogle Academic
Montopoli M, Ragazzi E, Froldi G, Caparrotta L. Inhibarea ciclului celular și apoptoza induse de curcumină și cisplatină sau oxaliplatină în celulele de carcinom ovarian uman. Celula Prolif. 2009;42(2):195–206.ArticolCASGoogle Academic
Pan W, Yang H, Cao C, Song X, Wallin B, Kivlin R, Lu S, Hu G, Di W, Wan Y. AMPK mediază moartea celulară indusă de curcumină în celulele cancerului ovarian CaOV3. Oncol Rep. 2008;20(6):1553–9.CASGoogle Academic
Yu Z, Wan Y, Liu Y, Yang J, Li L, Zhang W. Curcumina a indus apoptoza prin căile de semnalizare PI3K/Akt în celulele SKOV3. Pharm Biol. 2016;54(10):2026–32.ArticolCASGoogle Academic
Li-duan Z, Qiang-song T, Cui-huan W. Inhibarea creșterii și mecanismele de inducere a apoptozei ale curcuminei pe linia celulară de cancer ovarian uman A2780. Chin J Integr Med. 2006;12(2):126–31.ArticolGoogle Academic
Yin Z, Sun J. Curcumina induce apoptoza celulelor umane SKOV3 prin activarea Rho-kinazei. Eur J Gynaecol Oncol. 2014;35(4):433–7.CASGoogle Academic
Duse L, Agel MR, Pinnapireddy SR, Schafer J, Selo MA, Ehrhardt C, Bakowsky U. Terapia fotodinamică a celulelor de carcinom ovarian cu nanoparticule polimerice biodegradabile încărcate cu curcumină. Farmaceutică. 2019;11(6):282.ArticolCASGoogle Academic
Ramezani T, Nabiuni M, Baharara J, Parivar K, Namvar F. Sensitization of Resistance Ovarian Cancer Cells to Cisplatin by Biogenic Synthesized Silver Nanoparticules through p53 Activation. Iran J Pharm Res. 2019;18(1):222–31.CASGoogle Academic
Fatease AA, Shah V, Nguyen DX, Cote B, LeBlanc N, Rao DA, Alani AWG. Chemosensibilizarea și atenuarea cardiotoxicității induse de adriamicină folosind micelii polimerici combinați pentru co-livrarea de quercetină/resveratrol și resveratrol/curcumină în cancerul ovarian. Nanomedicina. 2019;19:39–48.ArticolCASGoogle Academic
Seyed Hosseini E, Alizadeh Zarei M, Babashah S, Nakhaei Sistani R, Sadeghizadeh M, Haddad Kashani H, Amini Mahabadi J, Izadpanah F, Atlasi MA, Nikzad H. Studies on combination of oxaliplatin and dendrosomal nanocurcumin on proliferation, and apoptosis in proliferation, and apoptosis expresie lungă a ARN-ului necodant în celulele canceroase ovariane. Cell Biol Toxicol. 2019;35(3):247–66.ArticolGoogle Academic
Dwivedi P, Yuan S, Han S, Mangrio FA, Zhu Z, Lei F, Ming Z, Cheng L, Liu Z, Si T și colab. Microîncapsularea curcuminei în microparticule PLGA: programată pentru aplicarea în terapia cancerului ovarian. Artif Cells Nanomed Biotechnol. 2018;46(sup3):S481–s491.ArticolCASGoogle Academic
Liu Z, Zhu YY, Li ZY, Ning SQ. Evaluarea eficacității paclitaxelului cu combinația de curcumină în celulele cancerului ovarian. Oncologie Let. 2016;12(5):3944–8.ArticolCASGoogle Academic
Liu L, Xiong X, Shen M, Ru D, Gao P, Zhang X, Huang C, Sun Y, Li H, Duan Y. Co-livrare de triptolide și curcumină pentru terapia de țintire a cancerului ovarian prin nanoparticule mPEG-DPPE/CaP. J Biomed Nanotechnol. 2018;14(10):1761–72.ArticolCASGoogle Academic
Du Z, Sha X. Demetoxicurcumin a inhibat creșterea celulelor de cancer ovarian epiteliu uman prin reglarea miR-551a. Tumor Biol. 2017;39(3):1010428317694302.ArticolCASGoogle Academic
Baghbani F, Moztarzadeh F. Ocolirea cancerului ovarian multidrog rezistent folosind doxorubicină/curcumină co-livrând nanopicături de alginat sensibile la ultrasunete. Coloizii Surf B, Biointerfețe. 2017;153:132–40.ArticolCASGoogle Academic
Alberti D, Protti N, Franck M, Stefania R, Bortolussi S, Altieri S, Deagostino A, Aime S, Geninatti Crich S. Theranostic Nanoparticles Loaded with Imaging Probes and Rubrocurcumin for Combined Cancer Therapy by Folate Receptor Targeting. ChemMedChem. 2017;12(7):502–9.ArticolCASGoogle Academic
Bondi ML, Emma MR, Botto C, Augello G, Azzolina A, Di Gaudio F, Craparo EF, Cavallaro G, Bachvarov D, Cervello M. Biocompatible Lipid Nanoparticules as Carriers To Improve Curcumin Efficacy in Ovarian Cancer Treatment. J Agric Food Chim. 2017;65(7):1342–52.ArticolCASGoogle Academic
Steuber N, Vo K, Wadhwa R, Birch J, Iacoban P, Chavez P, Elbayoumi TA. Platforma de nanoemulsie cu tocotrienol a curcuminei provoacă apoptoză crescută și creșterea eficacității anticanceroase împotriva carcinomului mamar și ovarian. Int J Mol Sci. 2016;17(11):1792.ArticolCASGoogle Academic
Pei H, Yang Y, Cui L, Yang J, Li X, Yang Y, Duan H. Bisdemetoxicurcumina inhibă cancerul ovarian prin reducerea expresiilor MMP mediate de stresul oxidativ. Sci Rep. 2016;6:28773.ArticolCASGoogle Academic
Arzuman L, Beale P, Yu JQ, Huq F. Sinteza clorurii de tris (chinolină) monocloroplatină (II) și activitatea sa singură și în combinație cu capsaicină și curcumină în liniile celulare de cancer ovarian uman. Anticancer Res. 2016;36(6):2809–18.CASGoogle Academic
Zhou L, Duan X, Zeng S, Men K, Zhang X, Yang L, Li X. Livrarea codificată a SH-aspirinei și curcuminei de către nanoparticulele mPEG-PLGA a îmbunătățit activitatea antitumorală prin inducerea apoptozei mitocondriale. Int J Nanomedicine. 2015;10:5205.ArticolCASGoogle Academic
Scarano W, De Souza P, Stenzel MH. Livrarea cu două medicamente a medicamentelor cu curcumină și platină în micelele polimerice sporește efectele sinergice: un act dublu pentru tratamentul cancerului multirezistent. Biomater Sci. 2015;3(1):163–74.ArticolCASGoogle Academic
Gou Q, Liu L, Wang C, Wu Q, Sun L, Yang X, Xie Y, Li P, Gong C. Nanoansamblurile polimerice care captează curcumina depășesc rezistența la mai multe medicamente în cancerul ovarian. Coloizii Surf B Biointerfețe. 2015;126:26–34.ArticolCASGoogle Academic
Arzuman L, Beale P, Chan C, Yu JQ, Huq F. Sinergism din combinații de clorură de tris (benzimidazol) monocloroplatină (II) cu capsaicină, quercetină, curcumină și cisplatină în liniile celulare de cancer ovarian uman. Anticancer Res. 2014;34(10):5453–64.CASGoogle Academic
Sarisozen C, Abouzeid AH, Torchilin VP. Efectul co-livrării de paclitaxel și curcumină prin micelii mixte pe bază de PEG-PE țintite pe transferină asupra cancerului ovarian rezistent în sferoizi 3-D și tumori in vivo. Eur J Pharm Biopharm. 2014;88(2):539–50.ArticolCASGoogle Academic
Abouzeid AH, Patel NR, Torchilin VP. Polietilen glicol-fosfatidiletanolamină (PEG-PE)/micele de vitamina E pentru administrarea concomitentă de paclitaxel și curcumină pentru a depăși rezistența la mai multe medicamente în cancerul ovarian. Int J Pharm. 2014;464(1-2):178–84.ArticolCASGoogle Academic
Kumar SSD, Surianarayanan M, Vijayaraghavan R, Mandal AB, Macfarlane DR. Nanoparticule de poli (2-hidroxietil metacrilat) încărcate cu curcumină din lichid ionic gelificat – Citotoxicitate in vitro și activitate anticanceroasă în celulele SKOV-3. Eur J Pharm Sci. 2014;51:34–44.ArticolCASGoogle Academic
Qu W, Xiao J, Zhang H, Chen Q, Wang Z, Shi H, Gong L, Chen J, Liu Y, Cao R. B19, un nou analog monocarbonil al curcuminei, induce apoptoza celulelor canceroase ovariane umane prin activarea reticulului endoplasmatic stresul și calea de semnalizare a autofagiei. Int J Biol Sci. 2013;9(8):766.ArticolCASGoogle Academic
Saxena V, MD Hussain. Miceliile polimerice amestecate pentru livrarea curcuminei în cancerul ovarian multidrog rezistent. J Biomed Nanotechnol. 2013;9(7):1146–54.ArticolCASGoogle Academic
Kanai M, Imaizumi A, Otsuka Y, Sasaki H, Hashiguchi M, Tsujiko K, Matsumoto S, Ishiguro H, Chiba T. Studiu de creștere a dozei și farmacocinetică a curcuminei nanoparticule, un potențial agent anticancer cu biodisponibilitate îmbunătățită, la voluntari umani sănătoși. Cancer Chemother Pharmacol. 2012;69(1):65–70.ArticolCASGoogle Academic
Ravindranath V, Chandrasekhara N. Studii in vitro privind absorbția intestinală a curcuminei la șobolani. Toxicologie. 1981;20(2-3):251–7.ArticolCASGoogle Academic
Ireson CR, Jones DJ, Orr S, Coughtrie MW, Boocock DJ, Williams ML, Farmer PB, Steward WP, Gescher AJ. Metabolizarea agentului chimiopreventiv al cancerului curcumină în intestinul uman și șobolan. Biomarkeri de epidemiol de cancer Prev. 2002;11(1):105–11.CASGoogle Academic
Sharma RA, McLelland HR, Hill KA, Ireson CR, Euden SA, Manson MM, Pirmohamed M, Marnett LJ, Gescher AJ, Steward WP. Studiu farmacodinamic și farmacocinetic al extractului oral de curcuma la pacienții cu cancer colorectal. Clin Cancer Res. 2001;7(7):1894–900.CASGoogle Academic
Sharma RA, Euden SA, Platton SL, Cooke DN, Shafayat A, Hewitt HR, Marczylo TH, Morgan B, Hemingway D, Plummer SM. Studiul clinic de fază I cu curcumină orală: biomarkeri ai activității sistemice și complianței. Clin Cancer Res. 2004;10(20):6847–54.ArticolCASGoogle Academic
Garcea G, Jones D, Singh R, Dennison A, Farmer P, Sharma R, Steward W, Gescher A, Berry D. Detectarea curcuminei și a metaboliților săi în țesutul hepatic și sângele portal al pacienților după administrare orală. Br J Cancer. 2004;90(5):1011–5.ArticolCASGoogle Academic
Cheng A, Hsu C, Lin J, Hsu M, Ho Y, Shen T, Ko J, Lin J, Lin B, Wu M. Studiul clinic de fază 1 cu curcumină, un agent chimiopreventiv. Pacienți cu risc ridicat de leziuni pre-maligne. Anticancer Res. 2001;21:2895–001.CASGoogle Academic
Lao CD, Ruffin MT, Normolle D, Heath DD, Murray SI, Bailey JM, Boggs ME, Crowell J, Rock CL, Brenner DE. Creșterea dozei unei formulări de curcuminoid. Complement BMC Altern Med. 2006;6(1):1–4.ArticolCASGoogle Academic
Soni S, Ak B, Rk S, Maitra A. Livrarea derivatului de 5-fluorouracil hidrofobizat la țesutul cerebral prin cale intravenoasă folosind nanogeluri modificate la suprafață. J țintă de droguri. 2006;14(2):87–95.ArticolCASGoogle Academic
Rai M, Pandit R, Gaikwad S, Yadav A, Gade A. Potential applications of curcumin and curcumin nanoparticules: from traditional therapeutics to modern nanomedicine. Nanotechnol Rev. 2015;4(2):161–72.ArticolCASGoogle Academic
Mahal A, Wu P, Jiang ZH, Wei X. Sinteza și activitatea citotoxică a noilor derivați de tetrahidrocurcumină care poartă fragment de pirazol. Nat Prod Bioprospect. 2017;7(6):461–9.ArticolCASGoogle Academic
Momtazi AA, Sahebkar A. Curcumină difluorinată: un analog de curcumină promițător cu activitate antitumorală îmbunătățită și profil farmacocinetic. Design farmaceutic actual. 2016;22(28):4386–97.ArticolCASGoogle Academic
Tripathi A, Misra K. Proiectarea și dezvoltarea de noi analogi/congeneri de curcumină ca inhibitori ai creșterii celulelor stem ale cancerului de sân. Chim Eng Trans. 2016;49:79–84.Google Academic
Borik RM, Fawzy NM, Abu-Bakr SM, Aly MS. Proiectare, sinteză, evaluare anticancer și studii de andocare a noilor derivați heterociclici obținuți prin reacții care implică curcumină. Molecule (Basel, Elveția). 2018;23(6):1398.ArticolCASGoogle Academic
Hackler L Jr, Ozsvari B, Gyuris M, Sipos P, Fabian G, Molnar E, Marton A, Farago N, Mihaly J, Nagy LI, et al. Analogul de curcumină C-150, care influențează căile NF-kappaB, UPR și Akt/Notch, are o activitate anticanceroasă puternică in vitro și in vivo. Plus unu. 2016;11(3):e0149832.ArticolCASGoogle Academic
Grynkiewicz G, Slifirski P. Curcumină și curcuminoizi în căutarea statutului medicinal. Acta biochimica Polonica. 2012;59(2):201–12.ArticolCASGoogle Academic
Bhattacharyya U, Kumar B, Garai A, Bhattacharyya A, Kumar A, Banerjee S, Kondaiah P, Chakravarty AR. „Medicament” de curcumină stabilizat în conjugați de oxidovanadiu (IV)-BODIPY pentru fotocitotoxicitatea țintită mitocondriilor. Inorg Chem. 2017;56(20):12457–68.ArticolCASGoogle Academic
Liu Y, Zhou J, Hu Y, Wang J, Yuan C. Curcumina inhibă creșterea celulelor cancerului de sân uman prin demetilarea promotorului DLC1. Mol Cell Biochim. 2017;425(1-2):47–58.ArticolCASGoogle Academic
Zhang J, Feng Z, Wang C, Zhou H, Liu W, Kanchana K, Dai X, Zou P, Gu J, Cai L și colab. Derivatul de curcumină WZ35 suprimă eficient progresia cancerului de colon prin inducerea producției de ROS și a apoptozei dependente de stres ER. Am J Cancer Res. 2017;7(2):275–88.CASGoogle Academic
Shehzad A, Khan S, Shehzad O, Lee YS. Curcumină promisiuni terapeutice și biodisponibilitate în cancerul colorectal. Droguri astăzi (Barc). 2010;46(7):523–32.ArticolCASGoogle Academic
Somers-Edgar TJ, Taurin S, Larsen L, Chandramouli A, Nelson MA, Rosengren RJ. Mecanisme pentru activitatea derivaților heterociclici de ciclohexanonă curcumină în liniile celulare de cancer de sân uman negativ pentru receptorul de estrogen. Investește noi medicamente. 2011;29(1):87–97.ArticolCASGoogle Academic
Kumar AP, Garcia GE, Ghosh R, Rajnarayanan RV, Alworth WL, Slaga TJ. Esterul metilic al acidului 4-hidroxi-3-metoxibenzoic: un derivat de curcumină vizează calea de semnalizare a supraviețuirii celulelor B Akt/NF kappa: potențialul de gestionare a cancerului de prostată. Neoplazie (New York, NY). 2003;5(3):255–66.ArticolCASGoogle Academic
Pan MH, Huang TM, Lin JK. Biotransformarea curcuminei prin reducere și glucuronidare la șoareci. Drug Metab Dispos. 1999;27(4):486–94.CASGoogle Academic
Heger M, van Golen RF, Broekgaarden M, Michel MC. Baza moleculară pentru farmacocinetica și farmacodinamia curcuminei și a metaboliților săi în relație cu cancerul. Pharmacol Rev. 2014;66(1):222–307.ArticolCASGoogle Academic
Sandur SK, Pandey MK, Sung B, Ahn KS, Murakami A, Sethi G, Limtrakul P, Badmaev V, Aggarwal BB. Curcumina, demetoxicurcumina, bisdemetoxicurcumina, tetrahidrocurcumina și turmeronii reglează diferențial răspunsurile antiinflamatorii și anti-proliferative printr-un mecanism independent de ROS. Carcinogeneza. 2007;28(8):1765–73.ArticolCASGoogle Academic
Somparn P, Phisalaphong C, Nakornchai S, Unchern S, Morales NP. Activități antioxidante comparative ale curcuminei și derivaților săi demetoxi și hidrogenați. Biol Pharm Bull. 2007;30(1):74–8.ArticolCASGoogle Academic
Prasad S, Tyagi AK, Aggarwal BB. Evoluții recente în ceea ce privește livrarea, biodisponibilitatea, absorbția și metabolismul curcuminei: pigmentul auriu din condimentul auriu. Cancer Res Treat. 2014;46(1):2.ArticolCASGoogle Academic
Pal A, Sung B, Prasad BAB, Schuber PT Jr, Prasad S, Aggarwal BB, Bornmann WG. Glucuronide de curcumină: Evaluarea activității proliferative împotriva liniilor celulare umane. Bioorg Med Chim. 2014;22(1):435–9.ArticolCASGoogle Academic
Xu H, Gong Z, Zhou S, Yang S, Wang D, Chen X, Wu J, Liu L, Zhong S, Zhao J. Curcumina lipozomală care vizează cancerul endometrial prin calea NF-κB. Cell Physiol Biochim. 2018;48(2):569–82.ArticolCASGoogle Academic
Hosseini ES, Zarei MA, Babashah S, Sistani RN, Sadeghizadeh M, Kashani HH, Mahabadi JA, Izadpanah F, Atlasi MA, Nikzad H. Studies on combination of oxaliplatin and dendrosomal nanocurcumin on proliferation, apoptosis induction, and long non-coding ARN expresie în celulele canceroase ovariane. Biol celular. 2019;35(3):247–66.Google Academic
Centrul de Cercetare Halal al IRI, FDA, Teheran, IranMohammad Hossein Pourhanifeh
Departamentul de Biotehnologie Medicală, Facultatea de Medicină, Universitatea de Științe Medicale Arak, Arak, IranMaryam Darvish
Membru al Facultății, Filiala Ardestan, Universitatea Islamic Azad, Ardestan, IranJavad Tabatabaeian & Mahboobeh Rabbani Fard
Departamentul de Chirurgie orală și maxilo-facială, Universitatea de Științe Medicale Kashan, Kashan, IranReza Mottaghi
Centrul de Cercetare în Traumă, Universitatea de Științe Medicale Kashan, Kashan, IranMohammad Javad Azadchehr
Centrul de dezvoltare a cercetării clinice (CRDC), Spitalul Sayad Shirazi, Universitatea de Științe Medicale Golestan, Gorgan, IranMoghaddaseh Jahanshahi
Centrul de Cercetare a Inflamației Neurogenice, Universitatea de Științe Medicale Mashhad, Mashhad, IranAmirhossein Sahebkar
Școala de Farmacie, Universitatea de Științe Medicale Mashhad, Mashhad, IranAmirhossein Sahebkar
Centrul de Cercetare pentru Biochimie și Nutriție în Boli Metabolice, Institutul de Științe de bază, Universitatea de Științe Medicale Kashan, Kashan, IranHamed Mirzaei
Contribuții
HM implicat în conceperea, proiectarea, analiza statistică și redactarea manuscrisului. MD, MH-P, RM, MR, JT, MJ, MJA și AS au contribuit la colectarea datelor și la redactarea manuscrisului. Toți autorii au aprobat versiunea finală pentru depunere.
Autorii nu au niciun conflict de interese de dezvăluit.
Informații suplimentare
Nota editorului
Springer Nature rămâne neutră în ceea ce privește revendicările jurisdicționale din hărțile publicate și afilierile instituționale.
Drepturi și permisiuni
Acces deschis Acest articol este licențiat în baza unei licențe internaționale Creative Commons Attribution 4.0, care permite utilizarea, partajarea, adaptarea, distribuirea și reproducerea în orice mediu sau format, atâta timp cât acordați un credit corespunzător autorilor originali și sursei, furnizați un link către licența Creative Commons și indicați dacă s-au făcut modificări. Imaginile sau alte materiale ale terților din acest articol sunt incluse în licența Creative Commons a articolului, cu excepția cazului în care se indică altfel într-o linie de credit a materialului. Dacă materialul nu este inclus în licența Creative Commons a articolului și utilizarea dorită nu este permisă de reglementările legale sau depășește utilizarea permisă, va trebui să obțineți permisiunea direct de la deținătorul drepturilor de autor. Pentru a vedea o copie a acestei licențe, vizitați http://creativecommons.org/licenses/by/4.0/ . Renunțarea la Creative Commons Public Domain Dedication ( http://creativecommons.org/publicdomain/zero/1.0/ ) se aplică datelor puse la dispoziție în acest articol, cu excepția cazului în care se specifică altfel într-o linie de credit a datelor.
Pourhanifeh, MH, Darvish, M., Tabatabaeian, J. şi colab. Rolul terapeutic al curcuminei și al formulărilor sale noi în cancerele ginecologice. J Ovarian Res 13 , 130 (2020). https://doi.org/10.1186/s13048-020-00731-7
Melatonina ( N-acetil-5-metoxitriptamina) este o indoleamina cu proprietati antioxidante, cronobiotice si antiinflamatorii; nivelurile reduse ale acestui hormon sunt asociate cu un risc mai mare de cancer. Mai multe efecte benefice ale melatoninei au fost descrise într-un număr mare de tumori, inclusiv cancere hepatice. În această lucrare am revizuit sistematic publicațiile din ultimii 15 ani care au evaluat mecanismele care stau la baza activităților melatoninei împotriva cancerelor hepatice și rolul său ca coadjuvant în tratamentul acestor tumori. Cercetarea literaturii a fost efectuată utilizând bazele de date PubMed, Scopus și Web of Science (WOS) și, după screening, au fost incluse 51 de articole. Rezultatele din studiile selectate au indicat acțiunile utile ale melatoninei în prevenirea carcinogenezei și ca opțiune de tratament promițătoare pentru tumorile hepatice primare, carcinomul hepatocelular (HCC) și colangiocarcinomul (CCA), fie singur, fie în combinație cu alți compuși. Diferite procese au fost modulate de indol, cum ar fi inhibarea stresului oxidativ, proliferarea, angiogeneza și invazia, promovarea răspunsului sistemului imunitar, oprirea ciclului celular și apoptoza, precum și recuperarea ritmurilor circadiene și modularea autofagiei. Luate împreună, prezenta revizuire sistematică evidențiază dovezile care documentează rolul potențial al melatoninei în îmbunătățirea peisajului tratamentului tumorii hepatice. cum ar fi inhibarea stresului oxidativ, proliferarea, angiogeneza și invazia, promovarea răspunsului sistemului imunitar, oprirea ciclului celular și apoptoza, precum și recuperarea ritmurilor circadiene și modularea autofagiei. Luate împreună, prezenta revizuire sistematică evidențiază dovezile care documentează rolul potențial al melatoninei în îmbunătățirea peisajului tratamentului tumorii hepatice. cum ar fi inhibarea stresului oxidativ, proliferarea, angiogeneza și invazia, promovarea răspunsului sistemului imunitar, oprirea ciclului celular și apoptoza, precum și recuperarea ritmurilor circadiene și modularea autofagiei. Luate împreună, prezenta revizuire sistematică evidențiază dovezile care documentează rolul potențial al melatoninei în îmbunătățirea peisajului tratamentului tumorii hepatice.
Cuvinte cheie: colangiocarcinom, carcinom hepatocelular, cancer hepatic, melatonină
Cancerul hepatic primar constituie al șaselea tip de tumoare cel mai răspândit și este a patra cauză comună de mortalitate legată de cancer la nivel mondial [ 1 ]. Un total de 841.080 de cazuri noi au fost diagnosticate în 2018, cu aproximativ 781.631 de decese și o rată a mortalității standardizată pe vârstă de 8,5/100.000. Cele mai frecvente tipuri de cancer hepatic primar la adulți sunt carcinomul hepatocelular (HCC) și colangiocarcinomul (CCA), constituind HCC 70–85% din cazuri și CCA 30–15%, în funcție de țară [2 ] . Cancerul hepatic apare și la copii și adolescenți, reprezentând hepatoblastomul (HB) și HCC pentru 67–80% și, respectiv, 20–33% din cazuri [ 2 , 3 ].
Din păcate, majoritatea pacienților cu cancer hepatic sunt diagnosticați în stadii avansate când tratamentul chirurgical nu mai este o opțiune disponibilă [ 4 ]. Mai mult, mortalitatea ridicată asociată cu cancerul hepatic este legată de lipsa de sensibilitate a acestuia și de dezvoltarea rezistenței la câteva tratamente care duc la eșecul chimioterapiei. Aceste dezavantaje pot fi explicate, cel puțin parțial, prin diferitele fenotipuri și caracteristici histologice ale celulelor hepatice tumorale și micromediul lor [ 5 ]. Mai mult, refractaritatea ridicată a cancerului hepatic la tratament a fost asociată cu interacțiunea unor mecanisme foarte complexe și diverse de chimiorezistență, care pot acționa sinergic pentru a proteja celulele tumorale de agenții chimioterapici [3 ] .
Melatonina ( N -acetil-5-metoxitriptamina), principalul produs al glandei pineale, este o indoleamină cu proprietăți antioxidante, cronobiotice și antiinflamatorii [ 6 ]. O reducere a nivelurilor de melatonina sau o excreție scăzută a principalului său metabolit, 6-sulfatoximelatonina, au fost legate de un risc crescut de cancer, sugerând un rol anticancerigen al acestei indoleamine [7 ] . În mod colectiv, datele publicate susțin puternic acțiunile oncostatice ale melatoninei asupra diferitelor tipuri de tumori, inclusiv cancerul hepatic [ 8 , 9 , 10 ].
Rolul inhibitor al melatoninei în tumorile hepatice implică o serie de procese moleculare și celulare diferite, inclusiv reducerea proliferării celulare, oprirea ciclului celular, limitarea angiogenezei și metastazei și promovarea morții celulare [ 10 , 11 , 12 , 13 , 14 , 15 , 16 , 17 ]. Mai mult, se pare că melatonina crește sensibilitatea celulelor canceroase hepatice la tratamentele disponibile în prezent [ 4 , 18 ].
În articolul de față, trecem în revistă în mod sistematic literatura științifică publicată în ultimii 15 ani, care se concentrează pe principalele mecanisme moleculare și celulare asociate cu efectele antitumorale ale melatoninei în cancerele hepatice și utilitatea potențială a acesteia în îmbunătățirea tratamentului acestor tipuri de tumori.
Această revizuire sistematică a fost efectuată urmând liniile directoare „Preferred Reporting Items for Systematic Reviews and Meta-Analyses” (PRISMA) [ 19 ] ( Tabelul S1 ). Pentru căutări cuprinzătoare în literatură, am efectuat căutarea articolelor în PubMed, Scopus și Web of Science (WOS) de la început până la 9 octombrie 2020, identificând un total de 369 de articole, 88 de la PubMed, 148 de la Scopus și 133 de la WOS.
Strategia de căutare completă a fost următoarea pentru fiecare bază de date:
PubMed: (“melatonină” [Toate câmpurile]) ȘI (“cancer la ficat” [Toate câmpurile] SAU “tumoare la ficat” [Toate câmpurile] SAU “hepatocarcinom”[Toate câmpurile] SAU “HCC”[Toate câmpurile] SAU “carcinom hepatocelular „[Toate câmpurile] SAU „angiosarcom”[Toate câmpurile] SAU „colangiocarcinom”[Toate câmpurile] SAU „hepatoblastom”[Toate domeniile]).
Scopus: TITOLU-ABS-KEY („melatonină”) ȘI („cancer la ficat” SAU „tumoare la ficat” SAU „hepatocarcinom” SAU „HCC” SAU „carcinom hepatocelular” SAU „angiosarcom” SAU „colangiocarcinom” SAU „hepatoblastom”) ).
WOS: TS = ((„melatonină”) ȘI („cancer hepatic” SAU „tumoare hepatică” SAU „hepatocarcinom” SAU „HCC” SAU „carcinom hepatocelular” SAU „angiosarcom” SAU „colangiocarcinom” SAU „hepatoblastom”).
2.2. Criterii de includere și excludere
Am aplicat următoarele criterii de includere pentru selectarea articolelor: (1) angajarea cancerului hepatic ca model de studiu; (2) utilizarea melatoninei ca tratament fie individual, fie în combinație cu un alt agent; (3) articole care au raportat efecte antioxidante sau antitumorale ale melatoninei. Au fost excluse articolele care îndeplineau următoarele criterii: (1) articole de recenzie sau compilații similare; (2) comunicații pentru conferințe sau congrese; (3) articole cu text integral într-o altă limbă decât engleza; (4) studii publicate înainte de 2005.
2.3. Selectarea studiilor
Doi autori au efectuat în mod independent selecția studiului și orice dezacord a fost rezolvat de un al treilea autor. După efectuarea căutării complete a articolelor originale, duplicatele dintre bazele de date au fost eliminate și articolele au fost supuse screening-ului. Lucrările au fost apoi selectate pe baza criteriilor de includere și au fost evaluate individual, eliminându-le pe cele care îndeplineau criteriile de excludere. În cele din urmă, lucrările originale care îndeplineau toate criteriile de eligibilitate au fost identificate și incluse în analiza calitativă.
2.4. Extragerea datelor
Datele din articolele incluse au fost extrase de doi autori independenți, compiland într-un tabel următoarele informații: numele primului autor și anul publicării, țara, tipul tumorii hepatice, modelul experimental (in vitro sau in vivo) și dimensiunea eșantionului, strategia de administrare a melatoninei , regimul de tratament (doză, frecvență și durată) și modificări ale proceselor studiate.
După căutările în baza de date și identificarea articolelor originale, au fost eliminate 156 de duplicate, iar 213 articole au fost analizate după titlu și rezumat. Dintre acestea, 57 de lucrări nu au îndeplinit criteriile de includere și, astfel, au fost aruncate. Restul de 156 de articole au fost analizate individual și 105 dintre ele care au îndeplinit criteriile de excludere au fost apoi eliminate, lăsând 51 de publicații identificate ca studii relevante care au fost incluse în această revizuire sistematică. Procesul de selecție a studiilor este descris înfigura 1.
Diagrama de flux a căutării și selecției studiilor efectuate în conformitate cu declarația PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses). WOS, Web of Science.
Un număr tot mai mare de articole care raportează proprietățile benefice ale melatoninei în tumorile hepatice au fost publicate în ultimii ani (Figura 2). Principalele date furnizate de studiile incluse sunt compilate întabelul 1. Dintre aceste publicații, cinci studii au fost efectuate cu modele CCA, iar restul de 46 au folosit modele HCC, ceea ce este în concordanță cu faptul menționat anterior că HCC și CCA sunt principalele tumori hepatice primare. De asemenea, se remarcă faptul că 50 de articole au analizat proprietățile melatoninei ca un singur tratament, în timp ce 32 au evaluat potențiale combinații cu agenți chimioterapeutici, inhibitori specifici sau compuși anticancer. În ceea ce privește modelul experimental, 21 de studii au investigat efectele melatoninei folosind modele animale, 17 în HCC și patru în CCA, în timp ce 36 de articole au implicat analize in vitro, 35 în HCC și doar unul în CCA, constatând și un raport de caz în HCC. Deși majoritatea articolelor au folosit modele celulare sau animale similare (linia celulară HepG2 umană și leziuni hepatice induse de DEN pentru HCC; linii de celule KKU-M055 și KKU-M214 șiOpisthorchis viverrini pentru CCA), intervalul de concentrație a melatoninei a fost larg, de la 0,01 nM la 10 mM în cultura celulară și de la 4 la 50 mg/kg în experimentele pe animale.
Distribuția temporală a numărului de articole publicate după 2004 care evaluează efectele melatoninei în tumorile hepatice doar cu modele in vitro sau in vivo, cu ambele tipuri de modele sau în raport de caz.
tabelul 1
Caracteristicile de bază ale studiilor experimentale care evaluează efectele antitumorale ale melatoninei împotriva tumorilor hepatice.
Primul autor, Anul publicării, [Referință]
Țară
Tumora hepatică
Model experimental ( n , dimensiunea eșantionului per grup)
Linii de celule HCC umane in vitro : HCC24/KMUH (rezistente la stresul oxidativ indus de AmB) și HCC38/KMUH: (susceptibil la stresul oxidativ indus de AmB)
Hamsteri aurii sirieni masculi in vivo de 4 până la 6 săptămâni Inoculare orală a 50 de metacercarii de Opisthorchis viverrini și 12,5 ppm DEN ( n = 15)
Hamsteri aurii sirieni masculi in vivo cu vârsta de 6 până la 8 săptămâni Infecție orală cu 50 de metacercarii de Opisthorchis viverrini și administrare orală cu DEN ( n = 7)
2-AAF, 2-acetilaminofluoren; AmB, amfotericină B; BCG, Bacilul Calmette-Guerin; Bmal1, proteina 1 asemănătoare creierului și mușchilor; CCA, colangiocarcinom; CDDP, Cis-diclorodiaminplatină; DEN, dietilnitrozamină; EGCG, (–)-epigalocatechin-3-galat; EMT, tranziție epitelial-mezenchimală; ER, reticul endoplasmatic; Exo-con, exozomi din celulele HepG2; Exo-MT, exozomi din celulele HepG2 tratate cu melatonină; HCC, hepatocarcinom; MF, câmp magnetic; IL-1p, interleukina 1 beta; IL-2, interleukină-2; MSC, celule stem mezenchimale; NF-kB, factor nuclear-kappa B; ARNi, ARN de interferență; siARN, ARN de interferență mic.
În ultimii ani, a existat o creștere constantă a numărului de publicații care utilizează atât metode in vivo, cât și in vitro, efectuate pentru a analiza rolul melatoninei în reducerea hepatocarcinogenezei. Acesta a fost cazul studiilor care au folosit melatonina ca unic tratament sau când a fost utilizată în asociere cu alte molecule.
Această secțiune următoare rezumă principalele constatări obținute în ultimii 15 ani. Deoarece HCC constituie cel mai frecvent cancer hepatic, majoritatea datelor sunt legate de acest fenotip de cancer; constatările privind alte tipuri de cancer sunt rezumate atunci când sunt disponibile.
3.2. Melatonina ca agent antioxidant și cronobiotic în cancerul hepatic
Procesele carcinogenetice implică adesea stres oxidativ, iar administrarea de molecule antioxidante poate reduce deteriorarea hepatocitelor canceroase [ 61 , 62 , 63 ]. Melatonina acționează ca un antioxidant indirect și ca un captator direct de radicali liberi și are un rol important ca agent imunomodulator și cronobiotic în diferite țesuturi, inclusiv în ficat [ 6 , 14 , 64 , 65 ].
Într-un model animal de HCC indus de administrarea de dietilnitrozamină (DEN) la șobolani, creșteri ale transaminazelor hepatice (aspartat și alanin transaminaze, AST și respectiv ALT), α-fetoproteina, greutatea ficatului și incidența tumorii (care sunt indici ai afectarii hepatice tumorale) au fost depistate. Mai mult, hepatocarcinogeneza a indus peroxidarea lipidelor (măsurată ca substanțe reactive la acid tiobarbituric – TBARS) și a scăzut activitatea serică a superoxid dismutază (SOD), catalaza (CAT), glutation peroxidază (GPx), glutation S-transferaza (GST) și glutation redus. (GSH), care este un sistem antioxidant major în toate țesuturile. Administrarea melatoninei (5 mg/kg greutate corporală timp de 20 de săptămâni) a abrogat aceste modificări și a redus stresul oxidativ și hepatocarcinogeneza [ 22].]. În alte experimente folosind același model in vivo de HCC, autorii au administrat suplimentar CCl4 cu DEN în timpul inducerii tumorii, constatând creșterea AST și ALT; dar, în mod curios, în această situație au fost crescute și nivelurile de TBARS și GSH, precum și activitățile GPx și GST, ceea ce se poate datora proliferării mai mari a celulelor tumorale. Administrarea melatoninei (5 mg/kg greutate corporală timp de 20 de săptămâni) a inversat toate aceste modificări la niveluri aproape de control, ceea ce probabil a contribuit la rolul său hepatoprotector [ 20 ].
În ciuda acțiunii antioxidante cunoscute a melatoninei, în anumite circumstanțe această indoleamină poate induce o creștere a stresului oxidativ. Grupul nostru a demonstrat că administrarea concomitentă a melatoninei (1 și 2 mM) cu sorafenib 2,5 mM (un medicament inhibitor al multikinazei care constituie tratamentul standard pentru HCC), în celulele Hep3B derivate din HCC uman, a produs acumularea de specii reactive de oxigen (ROS) în mitocondrii. . În plus, acest efect a indus depolarizarea membranei mitocondriale și, ulterior, activarea morții celulare prin mitofagie, un proces care poate fi util pentru a îmbunătăți sensibilitatea celulelor tumorale la sorafenib [ 18 ].
Din păcate, informațiile legate de efectele antitumorale ale melatoninei care implică capacitatea sa antioxidantă în alte cancere hepatice sunt mai limitate. Unele experimente efectuate folosind linii celulare CCA umane (KKU-M055 și KKU-M214) au demonstrat că administrarea de melatonine (0,5, 1 și 2 mM timp de 48 de ore) crește producția de ROS și citocrom C, ducând la moartea celulei prin apoptoză într-o concentrație – mod dependent [ 39 ]. Mai mult, a fost găsit într-un model in vivo de CCA la hamsteri dezvoltat prin infecția cu trematodul Opisthorchis viverrini.că administrarea melatoninei în doze diferite (5, 10 și 20 mg/kg greutate corporală timp de 30 de zile) reduce în mod dependent de doză activitatea ALT și stresul oxidativ/nitrozativ. În plus, melatonina a scăzut expresia ARNm a genelor generatoare de oxidanți, cum ar fi sintaza de oxid nitric inductibil (iNOS), factorul nuclear-kappa B (NF-κB), ciclooxigenaza-2 (COX-2) și citokinele proinflamatorii factorul de necroză tumorală alfa (TNF). -α) și interleukina 1 beta (IL-1p). Aceste efecte au fost însoțite de o creștere a expresiei genelor care codifică proteine antioxidante, inclusiv factorul 2 asociat eritroidului nuclear 2 (Nrf2) și mangan superoxid dismutază (Mn-SOD), o reducere a leziunilor hepatice (evaluată prin activitatea oxigenazei heme) și o diminuarea expresiei citokeratinei-19 (CK-19) – indicând o proliferare minoră a celulelor ductului biliar.26 ]. Aceste rezultate sugerează că modularea stresului oxidativ și nitrozativ de către melatonină ar putea fi utilă nu numai împotriva HCC, ci și în tratamentul și prevenirea CAA.
Deoarece ceasul circadian joacă un rol cheie în fiziologia ficatului și se știe că cronodisrupția crește hepatocarcinogeneza [ 66 ], au fost efectuate diferite experimente pentru a analiza asocierea dintre efectele antioxidante și cronobiotice ale melatoninei în HCC. Astfel, un studiu preliminar efectuat într-un model HCC indus de DEN la șoareci masculi indieni de câmp ( Mus booduga) au comparat efectele melatoninei (0,5 mg/kg greutate corporală) și oxaliplatinei (2 mg/kg greutate corporală) atunci când au fost administrate timp de 10 săptămâni; obiectivele au inclus dovezi de stres oxidativ și perturbări ale ritmului locomotor circadian. Rezultatele au arătat că ambele medicamente au ameliorat modificările activităților AST și ALT induse de DEN și au normalizat activitățile antioxidante SOD, CAT, GPx și GST fără diferențe semnificative între animalele tratate cu melatonină și oxaliplatină. Cu toate acestea, melatonina a fost mai eficientă decât oxaliplatina în contracararea modificărilor induse de DEN în ritmurile circadiene ale activității locomotorii [ 38 ].
Aceste rezultate au fost confirmate într-un alt studiu, folosind un model similar de HCC indus de DEN la șobolani, în care melatonină (5 mg/kg greutate corporală timp de 20 de săptămâni) și α-cetoglutarat (2 g/kg greutate corporală timp de 20 de săptămâni) au fost comparate efectele antitumorale asociate cu modificări ale ritmurilor circadiene ale sistemului antioxidant. În acest studiu, autorii au observat că tumorile detectabile la ficat au fost complet prevenite de melatonină sau α-cetoglutarat. Mai mult, administrarea DEN a crescut semnificativ mezorul și amplitudinea ritmurilor circadiene ale activităților AST și ALT și al a-fetoproteinei plasmatice (cu întârziere a acrofazei AST și ALT și avansarea fazei a-fetoproteinei); Administrarea de melatonină sau α-cetoglutarat a contracarat parțial aceste efecte. Mai mult, DEN a indus modificări ale valorilor mezorului diferiților parametri antioxidanti (creșterea mezorului TBARS, dar reducerea concentrației GSH mezor și activitățile SOD, CAT, GPx și GST), unde acrofazele au fost întârziate în toate cazurile. Melatonina, dar nu α-cetoglutaratul, mezorul TBARS redus și mezorul SOD crescut; cu toate acestea, modificările acrofazelor au fost prevenite doar parțial prin administrarea de melatonină sau α-cetoglutarat. În mod colectiv, aceste rezultate sugerează că melatonina, similar cu alte molecule antitumorale HCC, cum ar fi α-cetoglutaratul, poate modula ritmurile de 24 de ore de peroxidare a lipidelor și antioxidanți.23 ].
Rezultatele altor studii sunt în concordanță cu constatările anterioare. Astfel, s-a demonstrat că melatonina modulează ritmul circadian (acrofază, amplitudine și mezor) al peroxidării lipidelor, al enzimelor antioxidante și al nivelurilor de GSH. Experimentele in vivo, care induc HCC prin administrarea DEN la șobolani, au descoperit că hepatocarcinogeneza a indus valori mai mari ale peroxidării lipidelor (măsurate ca producție de TBARS), creșteri ale acrofazei SOD, CAT, GPx și GSH, împreună cu reduceri ale valorilor mezor ale aceste enzime antioxidante și GSH. Aceste modificări ale ritmurilor circadiene se pot datora utilizării sporite a acestor antioxidanți în timpul hepatocarcinogenezei și au fost inversate prin administrarea de melatonină (5 mg/kg greutate corporală timp de 20 de săptămâni), sugerând că efectele sale antitumorale ar putea fi asociate, cel puțin parțial,21 ].
În cele din urmă, modularea ritmurilor circadiene de către melatonină a fost implicată nu numai cu reglarea enzimelor antioxidante, ci și cu unele modificări metabolice. Un studiu combinat in vitro și in vivo, folosind celule 7288CTC HCC și xenogrefe tumorale 7288CTC in situ, a arătat că expunerea la lumina LED îmbogățită cu albastru de zi poate reduce progresia tumorii printr-un mecanism asociat cu creșterea nivelurilor endogene de melatonină. Acest lucru conduce la inhibarea metabolismului hepatomului prin suprimarea efectului Warburg (captarea de glucoză și producția de lactat) și împiedicând absorbția acidului linoleic în funcție de AMP ciclic (AMPc) [52 ] .
3.3. Oprirea ciclului celular de către melatonina în cancerul hepatic
Carcinogeneza este un proces în mai multe etape promovat de obicei de o proliferare celulară perturbată și necontrolată, cu dereglarea ciclului celular, asociată cu o creștere a viabilității celulelor tumorale. În HCC, expunerea la melatonină de 1 mM a celulelor HepG2 induce o scădere a procentului de celule în faza G0/G1 și o creștere a celulelor în faza S, confirmând efectul antiproliferativ al metoxiindolului [24] .]. Într-un studiu mai amplu, grupul nostru a demonstrat că administrarea in vitro de melatonină (0,1–10 mM timp de 10 zile) determină o reducere dependentă de doză și timp a viabilității celulelor HCC umane HepG2; aceste rezultate au fost asociate cu modificări în cinetica ciclului celular, cu o creștere a procentului de celule în faza G0/G1 însoțită de o reducere a numărului de celule în faza G2/M; aceste modificări au fost legate de inducerea expresiei p53 și activarea ulterioară a p21 (un inhibitor puternic al kinazelor ciclului celular – CDKs) [ 10 ]. Rezultatele au fost confirmate recent de alți autori, nu numai în HepG2, ci și în celulele Hep3B HCC; au raportat o reducere a expresiei ciclinei D1 și inducerea opririi ciclului celular cu melatonină 1 și 2 mM [ 59 ].
Creșteri ale expresiei proteice a principalilor receptori de melatonină din ficat (MT1, MT3 și receptorul orfan alfa-RORα legat de acid retinoic), care ar putea determina efectele citostatice și antiproliferative ale acestui indol, au fost găsite în celulele HepG2 după 1 an. şi administrare de melatonină 2,5 mM [ 11 ]. Activarea MT1 pare să fie legată de modularea cAMP și de membrii familiei protein kinazelor activate de mitogen (MAPK), cum ar fi kinaza 1 N-terminală c-Jun (JNK), kinaza reglată de semnal extracelular (ERK) și p38, dereglările de care se găsesc adesea în diferite tipuri de cancer, inclusiv HCC [ 12]. Mai mult, în CCA, experimentele in vitro și in vivo și datele din biopsiile umane au demonstrat o reducere a nivelurilor endogene de melatonina, dar creșteri ale receptorilor MT1 și MT2, care ar putea constitui un mecanism compensator al acestei tumori pentru a întârzia progresia creșterii celulare indusă de o secreție endogene de melatonină diminuată [ 28 ].
În mod curios, experimentele in vitro folosind celule HepG2 HCC expuse la câmp magnetic (50 Hz, 10 µT), care acționează ca un presupus inductor de proliferare celulară, au raportat că administrarea de 10 nM de melatonină produce efecte citostatice și antiproliferative. Această modificare a fost însoțită de o reducere a exprimării antigenului nuclear celular în proliferare (PCNA), o proteină nucleară nonhistone necesară pentru sinteza ADN-ului care este de obicei crescută în timpul fazei G1/S [30 ] .
Alte studii, folosind un model in vivo de HCC indus de DEN la șoareci, au observat că administrarea de melatonine (5 și 10 mg/kg greutate corporală, zilnic timp de 10, 20, 30 și 40 de săptămâni) induce atât expresia p21 cât și p53 într-un mod dependent de timp și doză; indolul atenuează, de asemenea, expresia proteinelor proliferative/reglatoare ale ciclului celular (Ki67, PCNA, ciclina D1, ciclina E, CDK4 și CDK6), reducând activitatea factorilor de transcripție precum NF-kB și transductorul de semnal și activatorul transcripției 3 ( STAT-3) și modularea sistemului sfingozin kinază/sphingozin 1-fosfat (SphK/S1P). Toate elementele menționate sunt frecvent asociate cu hepatocarcinogeneza [ 47]. Mai mult, în studiile in vitro folosind celule Hep3B HCC, s-a demonstrat că aceste efecte sunt asociate, cel puțin parțial, cu expresia normalizată a proteinei 1 (Bmal1) asemănătoare creierului și mușchilor și a ciclurilor circadiene de ieșire locomotorie a proteinei kaput ( Clock) proteine - care constituie componente ale mașinii circadiane în hepatocarcinogeneză – când s-a administrat 0,5 și 1 mM melatonină [ 14 ].
Administrarea combinată de melatonină poate îmbunătăți efectele citostatice și antiproliferative ale altor molecule împotriva HCC, cum ar fi sorafenib. Într-adevăr, combinația de 0,2-2 mM melatonină și sorafenib a îmbunătățit oprirea ciclului celular la faza G0/G1 atât în liniile celulare HCC Bel-7402, cât și SMMC-7721, prin reglarea p27 – un inhibitor al mai multor CDK și reglarea în jos a p -AKT și de asemenea, diferite proteine, inclusiv c-myc, ciclina D1 și CDK4/6, care joacă roluri cheie în proliferarea celulară. Aceste rezultate au fost validate folosind un model in vivo de tumoră xenogrefă tratată cu 25 mg/kg greutate corporală (zilnic timp de 18 zile) [ 46]]. Mai recent, un studiu preliminar folosind celule HepG2 HCC, a confirmat, de asemenea, că melatonina 3 mM a crescut efectele antiproliferative ale (–)-epigallocatechin-3-galat (EGCG) printr-un mecanism asociat p21 [56 ] .
3.4. Modularea apoptozei de către melatonina în cancerul hepatic
Rezistența la apoptoză este unul dintre principalele semne distinctive ale cancerului și inducerea sa selectivă în celulele canceroase a apărut ca o posibilitate interesantă pentru noi tratamente antitumorale. În ficat, semnalizarea apoptotică este transdusă în principal prin două căi moleculare: extrinsecă (sau mediată de receptorul morții) și intrinsecă (sau dependentă de mitocondrii). Calea extrinsecă este inițiată de liganzii morții, cum ar fi Fas-ligand (FasL), conducând la scindarea procaspazei-8 la forma sa activă, care ulterior reglează în sus efectorii din aval, cum ar fi caspaza-3. În schimb, calea intrinsecă este declanșată de disfuncția mitocondriilor, ducând la modificări ale proteinelor din familia Bcl-2, cum ar fi translocarea mitocondrială a proteinei proapoptotice Bax, care poate fi mediată de membrii superfamiliei MAPK, cum ar fi JNK și p38,67 , 68 ].
Experimentele in vitro folosind celule HepG2 umane, au demonstrat că administrarea de melatonină (0,1–10 mM timp de 10 zile) este capabilă să inducă atât căi extrinseci, cât și intrinseci de apoptoză, cu inducerea activității caspazei-8 independente de FasL, eliberarea citocromului C citosol. din mitocondrii cu reglarea în sus a lui Bax și inducerea activității caspazei-9 într-o manieră dependentă de doză și timp. Aceste mecanisme sunt responsabile pentru inducerea activității caspazei-3 și a proteolizei poli(ADP-ribozei) polimerazei (PARP), conducând la moartea hepatocitelor tumorale prin apoptoză [ 10 ]. În aceste condiții, activarea Bax este asociată cu supraexpresia Bim prin translocarea nucleară și activarea factorului de transcripție Forkhead box protein O3 (FoxO3a) atunci când este expus la 1 și 2 mM melatonină [29] .]. În plus, studiile in vitro care utilizează celule Hep3B HCC au demonstrat că modularea genelor implicate în mașinile circadiane (cum ar fi Bmal1, Clock sau Sirtuin 1—Sirt1) cu doze de melatonină de 0,5 și 1 mM poate fi asociată cu creșteri ale Bax/Bcl- 2 și ulterior inducerea apoptozei [ 14 ].
Melatonina, de asemenea, sensibilizează celulele HCC la stresul reticulului endoplasmatic (ER), inducând astfel apoptoza. În studiul în cauză, melatonina 1 nM a scăzut nivelurile de COX-2, care sunt de obicei crescute în celulele tumorale hepatice, promovând rata apoptozei prin creșterea nivelurilor factorului de transcripție proapoptotic CHOP (GADD153) și reducând raportul dintre Bcl- 2 proteina antiapoptotică și Bax proapoptotic (Bcl-2/Bax) în celulele HepG2 [ 32 ]. Efecte similare au fost găsite de alți autori, care au indicat în plus că inhibarea COX-2 poate fi asociată cu modificări ale factorului de transcripție activator 6 (ATF-6) de către melatonină 1 nM [41] .]. Aceste rezultate au fost confirmate folosind un model in vivo de HCC indus de DEN la șobolani, unde melatonina (1 mg/kg greutate corporală, zilnic timp de 45 și 90 de zile) a diminuat expresia COX-2, ceea ce duce la creșterea activității CHOP și ATF-6 și a redus raportul Bcl-2/Bax, determinând un răspuns de apoptoză prin reglarea în creștere a activității caspazelor-3, -8 și -9 și proteolizei PARP [17 ] .
Informațiile legate de efectul proapoptotic al melatoninei în monoterapie în alte cancere hepatice sunt mai limitate. Cu toate acestea, așa cum s-a indicat mai sus, experimentele efectuate folosind linii celulare umane CCA (KKU-M055 și KKU-M214) au demonstrat că concentrațiile de melatonină de 0,5, 1 și 2 mM timp de 48 de ore măresc producția de ROS și citocrom C, ducând la creșterea caspazei- 3 și expresia caspazei-6 și apoptoza ulterioară într-un mod dependent de doză [ 39 ]. Aceste rezultate au fost confirmate într-un model in vivo de CCA la hamsteri, indus de DEN și co-infectie cu Opisthorchis viverrini. În acest studiu, administrarea melatoninei (50 mg/kg greutate corporală, zilnic timp de 120 de zile) a activat calea intrinsecă a apoptozei prin reducerea raportului Bcl-2/Bax, creșterea eliberării citocromului C și activarea ulterioară a caspazei-3. Acest lucru a fost cauzat de un mecanism asociat cu atenuarea daunelor oxidative și nitrozative ADN care este legată de activarea Nrf2 și inhibarea căilor NF-κB [ 35 ]. Rezultate contrare au fost găsite de Colombo și colab. 2018 [ 51 ], unde melatonina 1 mM a indus expresia proteinei NF-κB în celulele HepG2 [ 51 ].
Diverse studii au indicat că melatonina sensibilizează celulele hepatice tumorale la apoptoză atunci când este administrată concomitent cu doxorubicină, un medicament anticancer foarte eficient, cu un spectru larg de activitate. În acest studiu, melatonina 10 nM în combinație cu doxorubicină inițiază un efect proapoptotic sinergic în celulele HCC HepG2 și Bel-7402 prin calea intrinsecă a apoptozei prin reglarea în sus a lui Bax, reglarea în jos a Bcl-2 și activarea caspazei-3 [25 ]]. Variază de la 1 nM la 1 mM de melatonină, prin creșterea nivelurilor CHOP și reducerea exprimării diferitelor proteine inhibitoare de apoptoză (supraviețuire, proteine apoptotice inhibitoare celulare și X-linked – cIAP și, respectiv, XIAP), au inversat rezistența indusă de stres ER la doxorubicină și creșterea apoptozei în liniile celulare HepG2 și SMMC-7721 HCC, printr-un mecanism dependent de COX-2/PI3K/AKT care a îmbunătățit răspunsul citotoxic la această moleculă [33 , 34 ] .
Sorafenib, așa cum sa indicat anterior, este un medicament de primă linie pentru tratamentul HCC avansat, dar în ciuda capacității sale de a crește supraviețuirea la pacienții cu HCC, succesul său este destul de scăzut pe termen lung, datorită dezvoltării celulelor rezistente prin mai multe mecanisme. [ 69 ]. Administrarea concomitentă a melatoninei 1-5 mM promovează apoptoza indusă de sorafenib prin activarea sinergică a căii JNK/c-jun și a caspazei-3 într-o manieră dependentă de doză în celulele HCC HuH7 [44 ] . Mai mult, așa cum s-a indicat mai sus, 50 pM melatonină a crescut, de asemenea, apoptoza indusă de regorafenib; această moleculă inhibă multiple kinaze și a fost aprobată pentru pacienții cu HCC rezistent la sorafenib [ 70]. S-a observat că melatonina poate spori reglarea Sirt3 asociată cu regorafenib, promovând disfuncția mitocondrială cu depolarizarea mitocondrială și inducerea apoptozei în celulele HCC SMMC-7721 [ 60 ].
Melatonina crește, de asemenea, efectul proapoptotic al cisplatinei, o moleculă antitumorală frecvent utilizată în tratamentul cancerului. Experimentele in vitro folosind celule Bel-7402, SNU-449, HepG2 și Hep3B HCC au demonstrat că tratamentul combinat al cisplatinei cu melatonină 1 mM crește calea intrinsecă a apoptozei și inhibă căile NF-κB/COX-2, îmbunătățind eficiența terapeutică. de cisplatină în HCC [ 43 ]. Mai recent, s-a demonstrat, de asemenea, că, în celulele HCC HepG2 și Hep3B, melatonina 1 și 2 mM mărește apoptoza indusă de cisplatină prin reglarea în jos a diferitelor proteine antiapoptotice (inclusiv Bcl-2, Bcl-xL sau Mcl-1) și reglarea clivată. caspaza-3 și PARP, efecte aparent legate de activarea căii de semnalizare Hippo, care promovează apoptoza și suprimă proliferarea celulară.59 ].
Analiza transcriptomică a genelor asociate cu rezistența la 5-fluorouracil (5-FU), care este un agent chimioterapeutic de rutină, a arătat că o doză de melatonină de 2,5 mM induce calea de semnalizare FoxO în celulele HepG2 și HuH7 și în probele de la pacienții cu HCC, îmbunătățind 5- Sensibilitatea FU [ 58 ].
În ultimii ani, a existat un interes din ce în ce mai mare pentru utilizarea flavonoidelor ca agenți anticancer [ 62 ]. Experimentele in vitro au demonstrat, de asemenea, că administrarea concomitentă de melatonină 2 și 4 mM potențează efectele proapoptotice ale EGCG în celulele HepG2 prin inhibarea Bcl-2 și COX-2, rezultând o citotoxicitate sporită împotriva celulelor HCC [56 ] .
În cele din urmă, într-un model in vivo de HCC după administrarea DEN la șobolani, melatonina (20 mg/kg greutate corporală, de două ori pe săptămână timp de 5 săptămâni) a îmbunătățit terapiile bazate pe celule stem mezenchimale (MSC) împotriva HCC prin diferite căi, inclusiv apoptoza prin reglarea în sus a Bax și a caspazei-3 și reglarea în jos a Bcl-2 și a supraviețuirii [ 54 ]. Aceste rezultate au fost raportate și de alți autori într-un model similar care folosește aceleași doze de indol, arătând că combinația de MSC și melatonină induce o activare mai mare a caspazelor-3 și -9 și apoptoza ulterioară [55 ] .
3.5. Modularea autofagiei de către melatonina în cancerul hepatic
Autofagia constituie un sistem de degradare în vrac care joacă un rol cheie în homeostazia celulară pentru a promova adaptarea și supraviețuirea celulară; totuși, atunci când stimularea excesivă duce la moartea celulară programată în loc de supraviețuire într-un număr de situații patofiziologice diferite, inclusiv tumorigeneză [ 71 ]. Prin urmare, în celulele canceroase, autofagia poate acționa ca o sabie cu două tăișuri îndepărtând celulele maligne noi și mitocondriile deteriorate în stadiile incipiente, dar inducerea supraviețuirii în condiții de hipoxie și ischemie în fazele ulterioare, poate fi, de asemenea, asociată cu rezistența la chimioterapie și progresia tumorii. 18 ].
Unele experimente in vivo au demonstrat că melatonina (10 și 20 mg/kg greutate corporală, zilnic timp de 14 zile) declanșează un proces de autofagie la șoarecii purtători de tumori H22, cu inducerea expresiei Beclin-1 și creșterea conversiei microtubulilor asociate. proteina 1 lanțul ușor 3 (LC3)-I la LC3-II (proteina asociată cu membrana autofagozomului) prin reducerea țintei de mamifere a rapamicinei (mTOR) și semnalizarea AKT. În mod interesant, blocarea acestui proces autofagic prin administrarea de Beclin-1 ARNi sau 3-metiladenină (3-MA) a sporit apoptoza asociată melatoninei în celulele HCC de șoarece H22 in vitro [31 ] .
Ceramidele au apărut ca efectori cheie în modularea autofagiei, mediand diafonia cu apoptoza [ 72] .]. Grupul nostru a descoperit că administrarea de melatonină 2 mM ar putea induce autofagie tranzitorie în celulele HepG2 asociate cu fosforilarea JNK, creșterea expresiei Beclin-1, degradarea p62 și co-localizarea LC3-II cu proteina membranară asociată lizozomal 2 (LAMP-2). Mai mult, în aceste condiții, tăcere siRNA a proteinei legate de autofagie 5 (ATG5) a suprimat modificările observate anterior în LC3-II și p62 induse de melatonină, dar a sensibilizat celulele HepG2 la apoptoza asociată melatoninei (cu creșterea proteolizei PARP și a caspazei). 3 activare). În plus, indolul a crescut nivelul de ceramide, dar perturbarea sintezei ceramidelor prin inhibarea serin palmitoil transferazei (SPT) sau a enzimelor sfingomielinazei acide (ASMaza) a dus la mecanisme diferite,16 ].
Modularea autofagiei pare să fie implicată în sensibilitatea celulelor tumorale și rezistența la tratamentele actuale împotriva HCC, cum ar fi sorafenib [ 73 ]. Alte experimente HCC in vitro folosind celule HepG2 și Bel-7402 au raportat în mod interesant că administrarea concomitentă a unei doze mai mici de melatonină (10 µM) cu sorafenib a redus autofagia indusă de sorafenib, arătând o reducere a raportului LC3-II/LC3-I și o creștere. a expresiei p62, însoțită de o creștere a apoptozei cu creșteri ale expresiei Bax și o scădere a Bcl-2 [ 45]. Aceste rezultate au fost confirmate recent de alți autori care au efectuat experimente folosind atât celule HCC (HepG2, 7721 și HuH7) cât și specimene HCC de la pacienți. Ei au raportat că administrarea concomitentă de doze mici de melatonină (10 µM) cu sorafenib crește moartea celulelor tumorale, blochează autofagia de supraviețuire indusă de sorafenib printr-o reducere a Beclin-1, dar depășind rezistența la apoptoză. Aceste rezultate par să fi fost asociate cu reținerea melatoninei a stresului ER printr-o reglare în jos atât a kinazei de stres ER asemănătoare PKR (PERK), cât și a factorului de transcripție de activare-4 (ATF-4), care, la rândul său, a redus expresia Beclin-1, conducând la conversie minoră a LC3-I în LC3-II. Acest blocaj al autofagiei de supraviețuire poate îmbunătăți efectele antitumorale ale sorafenibului în HCC [ 57 ].
Mitofagia, o formă distinctă de autofagie, promovează turnover-ul mitocondriilor deteriorate prinse în autofagolizozomi. După cum sa indicat mai sus, administrarea concomitentă a melatoninei (1 și 2 mM) cu sorafenib 2,5 mM în celulele Hep3B derivate din HCC uman, în condiții normoxice, a produs acumularea de ROS în mitocondrii și depolarizarea membranei mitocondriale. După aceste modificări, a fost detectată o co-localizare precoce a mitocondriilor cu lizozomi, corelând cu expresia kinazei putative 1 induse de PTEN (PINK1) și Parkin, care sunt markeri de mitofagie. Aceste efecte au potențat apoptoza celulelor tumorale (cu creșteri ale expresiei Bax și hidrolizei PARP) și au fost, de asemenea, asociate cu reducerea proliferării celulelor tumorale [ 18] .]. În schimb, în condiții de hipoxie, care este un fenotip cunoscut asociat cu eșecul chimioterapiei [ 74 ], mitofagia a acționat ca un mecanism citoprotector în celulele tumorale HCC. Astfel, în celulele Hep3B aflate în hipoxie, s-a demonstrat că administrarea concomitentă a melatoninei 2 mM blochează mitofagia citoprotectoare indusă de micromediul hipoxic după tratamentul cu sorafenib 5 mM; acest blocaj pare să fie asociat cu reglarea în jos a factorului inductibil de hipoxie-1α (HIF-1α) prin inhibarea complexului mTOR 1 (mTORC1)/proteina ribozomală S6 kinaza beta-1 (p70S6K)/proteina ribozomală S6 (RP-). S6) calea [ 4 ].
Administrarea concomitentă de melatonină 1 mM cu cisplatină, în celulele HepG2 HCC, a indus progresia autofagiei prin creșterea Beclin-1 și LC3-II, care a fost asociată cu suprimarea supraexprimării mTOR, precum și cu proteina complementară 1 încrucișată de reparare prin excizie ADN. (ERCC1) induse anterior de tratamentul cu cisplatină. Aceste rezultate confirmă că administrarea concomitentă a melatoninei beneficiază de efectele antitumorale ale cisplatinei în HCC [ 37 ].
3.6. Modularea angiogenezei și invazia de către melatonina în cancerul hepatic
Angiogeneza – procesul de creștere a noilor vase de sânge din sistemul vascular existent – joacă un rol cheie în facilitarea creșterii tumorii și a procesului metastatic și rezultă dintr-un echilibru dinamic între factorii proangiogenici și antiangiogenici. Angiogeneza a fost larg legată de progresia, invazia și metastaza tumorilor hepatice, iar inhibarea acesteia reprezintă o abordare testabilă pentru tratamentul acestui tip de cancer [ 75 , 76 , 77 ].
Condițiile normoxice din micromediul tumoral conduc la ubiquitinarea HIF-1α și la degradarea proteazomală ulterioară, în timp ce în hipoxie acest factor se stabilizează și se translocă în nucleu, inducând expresia mai multor gene, inclusiv factorul de creștere a endotelial vascular proangiogenic (VEGF) [69 ] . Mai mult, în hipoxie, fosforilarea STAT-3 pare să îmbunătățească stabilizarea HIF-1α, formând un complex transcripțional împreună cu co-activatorul CBP/p300, crescând astfel expresia VEGF și angiogeneza ulterioară [78] .]. Experimentele in vitro, efectuate în celule HepG2 tratate cu melatonină 1 mM, au demonstrat că această moleculă reduce expresia VEGF și HIF-1α, afectând migrarea și invazia celulelor și reducând proliferarea celulară atât în condiții de normoxie, cât și de hipoxie [40 ] . Experimente suplimentare in vitro, efectuate de asemenea în celule HepG2 tratate cu melatonină 1 mM sub hipoxie indusă de CoCl2, au arătat că melatonina nu numai că reduce proliferarea celulară, ci și interferează cu activarea transcripțională a VEGF ; aceasta blochează angiogeneza și invazivitatea celulelor tumorale, printr-o reducere a interacțiunii fizice dintre fosfo-STAT-3, HIF-1α și CBP/p300 în cadrul promotorului VEGF [13 ] .
Cercetările efectuate folosind celule HepG2 și HuH7 tratate cu melatonină 1 mM au demonstrat efectul antiproliferativ al acestei indoleamine, însoțit de inhibarea capacităților de migrare și invazie a celulelor într-o manieră dependentă de doză; aceste constatări au fost, de asemenea, confirmate într-un model in vivo de transplant de xenogrefă HCC în care animalele au fost tratate cu 40 mg/kg de melatonină în greutate corporală. Melatonina a crescut expresia factorului de transcripție forkhead box A2 (FOXA2), a indus suprareglarea transcriptului intrronic 1 ARN carbamoil-fosfat sintazei 1 lung necodificator (lncRNA-CPS1-IT1), care, la rândul său, a blocat HIF-1α care a suprimat tranziția epitelial-mezenchimală. progresia (EMT), migrarea celulelor și invazia [ 48]. Scăderi ale progresiei și invazivității în HCC au fost, de asemenea, raportate prin suprimarea reparării ADN-ului prin inducerea lncRNA RAD51 ARN antisens 1 (lncRNA RAD51-AS1) atunci când melatonina a fost administrată la concentrații de 1 mM în celulele HepG2 [50 ] . În plus, nu numai lncRNA-urile, ci și unele micro-ARN-uri au fost implicate în activitățile anti-invazive și antimetastatice ale melatoninei în HCC. În același mod, de asemenea, în liniile celulare HepG2 și HuH7, administrarea de melatonină 1 și 2 mM a indus activarea miRNA let7i-p, care, la rândul său, a reglat în jos proto-oncogenei factorului 1 activat ras (RAF-1), a afectat migrarea celulelor HCC. [ 53 ].
Metaloproteinazele matriceale (MMP) sunt o familie de endopeptidaze dependente de zinc care degradează matricea extracelulară, iar supraexpresia lor contribuie la creșterea și metastazarea HCC prin inducerea angiogenezei [ 79 ]. Printre aceste proteine, MMP-9 a fost asociată cu mai multe etape de creștere a tumorii, invazie vasculară și metastaze [ 80 ]. S-a raportat că melatonina 1 mM a redus activitatea MMP-9 indusă de IL-1p, afectând invazia și motilitatea celulară în celulele HepG2. Acest efect a fost legat de suprimarea translocării NF-κB induse de IL-1β și a activității transcripționale, ceea ce a condus la reglarea în jos a expresiei genei MMP-9, care a coincis cu reglarea în sus a inhibitorului de țesut inhibitor specific MMP-9 al metaloproteinazelor ( TIMP)-1 de melatonină [ 15].
Unii autori au indicat că modularea HCC de către melatonină depinde atât de caracteristicile celulelor tumorale, cât și de concentrația moleculei administrate. Într-adevăr, un studiu in vitro utilizând două doze diferite de melatonină (1 µM și 100 µM) administrate la două linii de celule umane HCC diferite, HCC24/KMUH care nu sunt susceptibile la stresul indus de amfotericina B (AmB) și HCC38/KMUH care sunt susceptibile. , a produs rezultate contradictorii în ceea ce privește exprimarea chemokinelor proangiogenice și angiostatice sub stresul indus de AmB. Prin urmare, expresia agenților proangiogenici, cum ar fi ligandul 2 al chemokinei (motivul CC) (CCL-2) și interleukina (IL-8) a fost indusă atât prin administrarea de melatonină 1 µM, cât și 100 µM în celulele HCC24/KMUH, dar, în mod curios, expresia Ligandul 10 al chemokinei cu motiv CXC angiostatic (CXCL-10) a fost de asemenea crescut. In orice caz, expresia CCL-2 și IL-8, împreună cu o altă chemokină proangiogenă numită ligandul 6 al chemokinei cu motiv CXC (CXCL-6) și enzima antioxidantă Mn-SOD, au fost reduse de ambele concentrații de melatonină în HCC38/KMUH. Acest lucru a arătat că în condiții de stres AmB, melatonina inhibă semnalul proangiogenic și acționează ca un antioxidant substituind funcția Mn-SOD. Descoperirile indică faptul că aplicarea clinică a melatoninei la pacienții cu HCC ar trebui să ia în considerare caracteristicile tumorii hepatice pentru optimizarea concentrațiilor de indol. Acest lucru a arătat că în condiții de stres AmB, melatonina inhibă semnalul proangiogenic și acționează ca un antioxidant substituind funcția Mn-SOD. [.27 ].
3.7. Imunomodularea melatoninei în cancerul hepatic
După cum s-a indicat anterior, melatonina funcționează atât ca antioxidant și cronobiotic, cât și ca agent imunomodulator [ 6 , 14 , 64 , 65 ]. Unele rapoarte de caz la oameni au arătat că administrarea concomitentă de 20 mg/zi de melatonină împreună cu imunoterapie constând din interleukină-2 (IL-2) și Bacil Calmette-Guerin, după îndepărtarea chirurgicală inițială a nodulilor, are ca rezultat o reducere a progresiei HCC. și dezvoltarea de noi tumori hepatice [ 36 ]. Cu toate acestea, sunt necesare studii suplimentare care să analizeze mecanismele moleculare și celulare implicate în rolul imunomodulator al melatoninei asupra cancerului hepatic.
Unele informații relevante au fost obținute dintr-un studiu interesant in vitro și in vivo folosind exozomi derivați de HCC netratați (Exo-con) sau tratați cu 0,1 mM tratați cu melatonină (Exo-MT), care au fost co-cultivați cu THP- uman. 1 și linii celulare de monocite/macrofage RAW264.7 de șoarece sau injectate în șoareci nuzi BALB/C. Rezultatele acestor experimente au sugerat că Exo-con reglează expresia ligandului morții programate 1 (PD-L1) și secreția de citokine inflamatorii (IL-6, IL-10, IL-1β și TNF-α) în macrofage, deoarece aceste modificări au fost blocate în inactivarea STAT-3 a specimenelor Exo-Mt, prin urmare modulând răspunsul imun [ 42 ].
În mod similar, într-un model in vivo de CCA indus de DEN și co-infectat cu Opisthorchis viverrine , s-a observat proliferarea celulelor căilor biliare cu inducerea expresiei factorului de creștere transformator β (TGF-β). Aceste modificări au fost însoțite de infiltrarea celulelor T helper (Th17) producătoare de IL-17 și de supraexpresie forkhead box P3 (foxp3) – care este un regulator principal în dezvoltarea și funcția celulelor T reglatoare, împreună cu creșteri ale celulelor inflamatorii, în special eozinofilele. și inducerea expresiei NF-kB, COX-2 și IL-1. Administrarea melatoninei (50 mg/kg greutate corporală, zilnic timp de 30 de zile) a suprimat toate aceste modificări, sugerând că indolul ar putea fi eficient în tratamentul și prevenirea CCA printr-un mecanism dublu care implică atât imunomodularea, cât și capacitatea sa antiinflamatoare.49 ].
3.8. Limitări
Din câte știm, aceasta este prima revizuire sistematică care rezumă efectele antitumorale ale melatoninei în cazul cancerului hepatic și a fost scrisă cu intenția de a oferi o mai bună înțelegere a mecanismelor implicate. Cu toate acestea, există unele limitări, în principal din cauza cantității mari și eterogenității articolelor incluse în cele din urmă. Deși rolul antioxidant al melatoninei este bine stabilit, există și alte investigații în care acest indol a promovat generarea de ROS atât în HCC, cât și în CCA, ceea ce s-ar putea datora concentrațiilor mari de melatonină uneori folosite. Aceasta poate fi una dintre principalele limitări; astfel, atât in vitro, cât și in vivo, doza/concentrația de melatonină a variat mult între experimentele rezumate aici. Acest lucru a împiedicat concluziile ferme referitoare la eficacitatea absolută a melatoninei ca tratament pentru cancerul de ficat. Din fericire, melatonina nu are toxicitate semnificativă atunci când este administrată animalelor, inclusiv oamenilor și poate fi administrată în cantități într-un interval foarte mare. Mai mult, de ani de zile se știe că concentrațiile necesare pentru a documenta efectele oncostatice ale melatoninei in vitro sunt mult mai mari decât se anticipa în comparație cu rezultatele obținute in vivo. Acest lucru a fost interpretat ca însemnând că în condiții in vitro toate acțiunile melatoninei nu pot fi manifestate, deoarece metabolismul celulelor în cultură probabil diferă de cel in vivo. de ani de zile se știe că concentrațiile necesare pentru a documenta efectele oncostatice ale melatoninei in vitro sunt mult mai mari decât se anticipa în raport cu constatările obținute in vivo. Acest lucru a fost interpretat ca însemnând că în condiții in vitro toate acțiunile melatoninei nu pot fi manifestate, deoarece metabolismul celulelor în cultură probabil diferă de cel in vivo. de ani de zile se știe că concentrațiile necesare pentru a documenta efectele oncostatice ale melatoninei in vitro sunt mult mai mari decât se anticipa în raport cu constatările obținute in vivo. Acest lucru a fost interpretat ca însemnând că în condiții in vitro toate acțiunile melatoninei nu pot fi manifestate, deoarece metabolismul celulelor în cultură probabil diferă de cel in vivo.81 ].
În ceea ce privește ciclul celular, există o lipsă de investigație privind efectele în CCA, în timp ce oprirea ciclului celular in vivo indusă de melatonină este raportată în toate studiile HCC. În mod similar, evaluarea autofagiei în CCA este, de asemenea, limitată, în timp ce a fost evaluată pe deplin în HCC. Rezultatele studiilor privind autofagia sunt oarecum divergente, deoarece autofagia acționează ca un proces cu două tăișuri în cancer, iar melatonina exercită, de asemenea, acțiuni duale asupra modulării mașinării autofagiei în funcție de micromediul celular; Specificitatea contextului în raport cu acțiunile melatoninei în cancer au fost, de asemenea, raportate anterior [ 82]. În schimb, efectele melatoninei asupra angiogenezei și abilităților invazive și migratorii în HCC par să fie consistente între investigații, cu unele rezultate diferite cauzate de caracteristicile tumorale diferite și concentrațiile de melatonină utilizate.
Una dintre cele mai puțin studiate proprietăți ale melatoninei în ceea ce privește cancerul hepatic este imunomodularea. Deși o activitate imunomodulatoare clară asociată cu acțiunea antiinflamatoare a melatoninei a fost demonstrată atât în HCC, cât și în CCA, sunt necesare studii suplimentare pentru a stabili mecanismele care stau la baza. Deși numărul de studii în care au fost evaluate strategiile de combinare cu melatonina este limitat, articolele care analizează coadministrarea cu medicamente antitumorale, flavonoide sau molecule inhibitoare demonstrează un rol potențial al melatoninei în îmbunătățirea sensibilității cancerelor hepatice la mai multe medicamente. Pe scurt, există încă deficiențe în ceea ce privește utilitatea potențială a melatoninei ca agent anti-cancer, nu numai ca unic, ci și ca tratament combinat, în diferitele procese descrise aici.
Aceasta este prima revizuire sistematică care reunește efectele antitumorale exercitate de melatonina în tumorile hepatice, un cancer din păcate obișnuit și mortal pentru care există opțiuni limitate de tratament. Rezultatele studiilor analizate aici sugerează că melatonina are în general proprietăți benefice în prevenirea și tratamentul principalelor tumori hepatice primare, HCC și CCA, prin modularea unui număr mare de procese celulare, protejând chiar celulele de hepatotoxicitatea altor medicamente. Printre efectele melatoninei, așa cum se arată înFigura 3, îmbunătățește răspunsul imun, îmbunătățește apoptoza și influențează pozitiv ciclul celular și ritmurile circadiene; de asemenea, afectează angiogeneza tumorii, invazia și proliferarea celulară. Având în vedere diferitele sale roluri asupra autofagiei, melatonina poate modula acest proces în funcție de contextul celular, având întotdeauna ca scop împiedicarea progresiei tumorii. În cele din urmă, studiile combinate cu diferite tipuri de molecule, medicamente antitumorale, flavonoide și chimioterapice, oferă dovezi ale efectelor potențiale ale melatoninei ca agent coadjuvant pentru îmbunătățirea tratamentelor actuale. În general, această revizuire sistematică oferă informații utile și susține rolul melatoninei ca medicament promițător în armamentul de tratament al tumorilor hepatice.
CIBERehd este finanțat de Instituto de Salud Carlos III, Spania. PF-P. și FF sunt susținute de Ministerul Educației din Spania (FPU17/01995 și, respectiv, FPU16/05277) și CM-B. este susținut de Asociația Española Contra el Cancer (AECC)-Junta Provincial de León.
Conceptualizare, JLM, JG-G., MJT și RJR; metodologie, PF-P. și JLM; analiza formală, toți autorii; investigație, toți autorii; redactare—pregătirea proiectului original, JLM și PF-P.; scriere – recenzie și editare, toți autorii; vizualizare, toți autorii; supraveghere, JLM și JG-G. Toți autorii au citit și au fost de acord cu versiunea publicată a manuscrisului.
Bray F., Ferlay J., Soerjomataram I., Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimations of incident and mortality worldwide for 36 cancers in 185 states. CA Cancer J. Clin. 2018; 68 :394–424. doi: 10.3322/caac.21492. [ PubMed ] [ CrossRef ] [ Google Scholar ]2.
Sharma R. Epidemiologia descriptivă a incidenței și mortalității cancerului primar de ficat în 185 de țări: Evidence from GLOBOCAN 2018. Jpn. J. Clin. Oncol. 2020; 50 :1370–1379. doi: 10.1093/jjco/hyaa130. [ PubMed ] [ CrossRef ] [ Google Scholar ]3.
Marin JJG, Briz O., Herraez E., Lozano E., Asensio M., Di Giacomo S., Romero MR, Osorio-Padilla LM, Santos-Llamas AI, Serrano MA, et al. Bazele moleculare ale răspunsului slab al cancerului hepatic la chimioterapie. Clin. Res. Hepatol. Gastroenterol. 2018; 42 :182–192. doi: 10.1016/j.clinre.2017.12.006. [ PubMed ] [ CrossRef ] [ Google Scholar ]4.
Prieto-Domínguez N., Méndez-Blanco C., Carbajo-Pescador S., Fondevila F., García-Palomo A., González-Gallego J., Mauriz JL Melatonina îmbunătățește acțiunile sorafenibului în celulele hepatocarcinomului uman prin inhibarea mTORC1/ p70S6K/HIF-1α și mitofagie mediată de hipoxie. Oncotarget. 2017; 8 :91402–91414. doi: 10.18632/oncotarget.20592. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]5.
Wang G., Wang Q., Liang N., Xue H., Yang T., Chen X., Qiu Z., Zeng C., Sun T., Yuan W. și colab. Genele driver oncogene și micromediul tumoral determină tipul de cancer la ficat. Moartea celulară Dis. 2020; 11 :313. doi: 10.1038/s41419-020-2509-x. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]6.
Mauriz JL, Collado PS, Veneroso C., Reiter RJ, González-Gallego J. O revizuire a aspectelor moleculare ale acțiunilor antiinflamatorii melatoninei: perspective recente și perspective noi. J. Pineal Res. 2013; 54 :1–14. doi: 10.1111/j.1600-079X.2012.01014.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]7.
Sánchez-Hidalgo M., Guerrero JM, Villegas I., Packham G., de la Lastra CA Melatonina, un inductor natural de moarte celulară programată în cancer. Curr. Med. Chim. 2012; 19 :3805–3821. doi: 10.2174/092986712801661013. [ PubMed ] [ CrossRef ] [ Google Scholar ]8.
Reiter RJ, Rosales-Corral SA, Tan DX, Acuña-Castroviejo D., Qin L., Yang SF, Xu K. Melatonina, un agent anti-cancer cu servicii complete: inhibarea inițierii, progresiei și metastazelor. Int. J. Mol. Sci. 2017; 18 :843. doi: 10.3390/ijms18040843. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]9.
Li Y., Li S., Zhou Y., Meng X., Zhang J.-J., Xu D.-P., Li H.-B. Melatonina pentru prevenirea și tratamentul cancerului. Oncotarget. 2017; 8 :39896–39921. doi: 10.18632/oncotarget.16379. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]10.
Martín-Renedo J., Mauriz JL, Jorquera F., Ruiz-Andrés O., González P., González-Gallego J. Melatonina induce stoparea ciclului celular și apoptoza în linia celulară HepG2 de hepatocarcinom. J. Pineal Res. 2008; 45 :532–540. doi: 10.1111/j.1600-079X.2008.00641.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]11.
Carbajo-Pescador S., Martín-Renedo J., García-Palomo A., Tuñón MJ, Mauriz JL, González-Gallego J. Modificări în expresia receptorilor melatoninei induse de tratamentul cu melatonina în celulele HepG2 de hepatocarcinom. J. Pineal Res. 2009; 47 :330–338. doi: 10.1111/j.1600-079X.2009.00719.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]12.
Carbajo-Pescador S., García-Palomo A., Martín-Renedo J., Piva M., González-Gallego J., Mauriz JL Modularea melatoninei a căilor de semnalizare intracelulară în linia celulară HepG2 de hepatocarcinom: Rolul receptorului MT1. J. Pineal Res. 2011; 51 :463–471. doi: 10.1111/j.1600-079X.2011.00910.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]13.
Carbajo-Pescador S., Ordoñez R., Benet M., Jover R., García-Palomo A., Mauriz JL, González-Gallego J. Inhibarea expresiei VEGF prin blocarea semnalizării Hif1α și STAT3 mediază anti-angiogenic efectul melatoninei în celulele canceroase hepatice HepG2. Br. J. Cancer. 2013; 109 :83–91. doi: 10.1038/bjc.2013.285. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]14.
Sánchez DI, González-Fernández B., Crespo I., San-Miguel B., Álvarez M., González-Gallego J., Tuñón MJ Melatonina modulează ceasurile circadiene dereglate la șoareci cu carcinom hepatocelular indus de dietilnitrozamină. J. Pineal Res. 2018; 65 :e12506. doi: 10.1111/jpi.12506. [ PubMed ] [ CrossRef ] [ Google Scholar ]15.
Ordoñez R., Carbajo-Pescador S., Prieto-Dominguez N., García-Palomo A., González-Gallego J., Mauriz JL Inhibarea metaloproteinazei-9 a matricei și a factorului nuclear kappa B contribuie la prevenirea motilității și a melatoninei invazivitatea celulelor canceroase hepatice HepG2. J. Pineal Res. 2014; 56 :20–30. doi: 10.1111/jpi.12092. [ PubMed ] [ CrossRef ] [ Google Scholar ]16.
Ordoñez R., Fernández A., Prieto-Domínguez N., Martínez L., García-Ruiz C., Fernández-Checa JC, Mauriz JL, González-Gallego J. Metabolismul ceramidei reglează autofagia și moartea celulară apoptotică indusă de melatonină în celulele canceroase hepatice. J. Pineal Res. 2015; 59 :178–189. doi: 10.1111/jpi.12249. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]17.
Moreira AJ, Ordoñez R., Cerski CT, Picada JN, García-Palomo A., Marroni NP, Mauriz JL, González-Gallego J. Melatonina activează stresul reticulului endoplasmatic și apoptoza la șobolani cu hepatocarcinogeneză indusă de dietilnitrozamină. Plus unu. 2015; 10 :e0144517. doi: 10.1371/journal.pone.0144517. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]18.
Prieto-Domínguez N., Ordóñez R., Fernández A., Méndez-Blanco C., Baulies A., Garcia-Ruiz C., Fernández-Checa JC, Mauriz JL, González-Gallego J. Melatonin-induced increase in sensibilitatea celulelor de carcinom hepatocelular uman la sorafenib este asociată cu producția de specii reactive de oxigen și mitofagie. J. Pineal Res. 2016; 61 :396–407. doi: 10.1111/jpi.12358. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]19.
Moher D., Liberati A., Tetzlaff J., Altman DG Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA statement. PLoS Med. 2009; 6 :e1000097. doi: 10.1371/journal.pmed.1000097. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]20.
Dakshayani KB, Subramanian P., Manivasagam T., Mohamed Essa M., Manoharan S. Melatonina modulează dezechilibrul oxidant-antioxidant în timpul hepatocarcinogenei induse de N-nitrosodietilamină la șobolani. J. Pharm. Farmacia. Sci. 2005; 8 :316–321. [ PubMed ] [ Google Scholar ]21.
Dakshayani KB, Subramanian P., Essa MM Efectul melatoninei asupra hepatocarcinogenezei induse de N-nitrosodietilamină la șobolani cu referire la ritmurile circadiene biochimice. Toxicol. Mech. Metode. 2007; 17 :67–75. doi: 10.1080/15376520500195798. [ PubMed ] [ CrossRef ] [ Google Scholar ]22.
Subramanian P., Mirunalini S., Dakshayani KB, Pandi-Perumal SR, Trakht I., Cardinali DP Prevenirea prin melatonina a hepatocarcinogenezei la șobolani injectați cu N-nitrosodietilamină. J. Pineal Res. 2007; 43 :305–312. doi: 10.1111/j.1600-079X.2007.00478.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]23.
Subramanian P., Dakshayani KB, Pandi-Perumal SR, Trakht I., Cardinali DP Ritmuri de 24 de ore în stresul oxidativ în timpul hepatocarcinogenezei la șobolani: Efectul melatoninei sau al α-cetoglutaratului. Redox Rep. 2008; 13 :78–86. doi: 10.1179/135100008X259178. [ PubMed ] [ CrossRef ] [ Google Scholar ]24.
Ozdemir F., Deniz O., Kaynar K., Arslan M., Kavgaci H., Yildiz B., Aydin F. Efectele melatoninei asupra liniei celulare de hepatom uman (Hep G2). Bratislava Med. J. 2009; 110 :276–279. [ PubMed ] [ Google Scholar ]25.
Fan L.-L., Sun G.-P., Wei W., Wang Z.-G., Ge L., Fu W.-Z., Wang H. Melatonina și doxorubicina induc sinergic apoptoza celulară la om linii celulare de hepatom. Lumea J. Gastroenterol. 2010; 16 :1473–1481. doi: 10.3748/wjg.v16.i12.1473. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]26.
Laothong U., Pinlaor P., Hiraku Y., Boonsiri P., Prakobwong S., Khoontawad J., Pinlaor S. Protective effect of melatonin against Opisthorchis viverrini -induced oxidative and nitrosative ADN and nitrosative injuries the hamsters. J. Pineal Res. 2010; 49 :271–282. doi: 10.1111/j.1600-079X.2010.00792.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]27.
Lin ZY, Chuang WL Concentrațiile farmacologice de melatonină au influență diversă asupra expresiilor diferențiale ale genelor chemokine angiogenice în diferite linii celulare de carcinom hepatocelular. Biomed. Pharmacother. 2010; 64 :659–662. doi: 10.1016/j.biopha.2010.09.006. [ PubMed ] [ CrossRef ] [ Google Scholar ]28.
Han Y., DeMorrow S., Invernizzi P., Jing Q., Glaser S., Renzi A., Meng F., Venter J., Bernuzzi F., White M., et al. Melatonina exercită printr-o buclă autocrină efecte antiproliferative în colangiocarcinom; sinteza acestuia este redusă favorizând creşterea colangiocarcinomului. A.m. J. Physiol. Test gastro-intestinal. Ficat Physiol. 2011; 301 :623–633. doi: 10.1152/ajpgi.00118.2011. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]29.
Carbajo-Pescador S., Steinmetz C., Kashyap A., Lorenz S., Mauriz JL, Heise M., Galle PR, González-Gallego J., Strand S. Melatonina induce reglarea transcripțională a Bim de către FoxO3a în celulele HepG2 . Br. J. Cancer. 2013; 108 :442–449. doi: 10.1038/bjc.2012.563. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]30.
Cid MA, Úbeda A., Hernández-Bule ML, Martínez MA, Trillo M.Á. Efectele antagoniste ale unui câmp magnetic de 50 Hz și ale melatoninei în proliferarea și diferențierea celulelor hepatocarcinomului. Celulă. Physiol. Biochim. 2012; 30 :1502–1516. doi: 10.1159/000343338. [ PubMed ] [ CrossRef ] [ Google Scholar ]31.
Liu C., Jia Z., Zhang X., Hou J., Wang L., Hao S., Ruan X., Yu Z., Zheng Y. Implicarea melatoninei în supraviețuirea celulelor H22 de hepatom de șoarece mediată de autofagie. Int. Imunofarmacol. 2012; 12 :394–401. doi: 10.1016/j.intimp.2011.12.012. [ PubMed ] [ CrossRef ] [ Google Scholar ]32.
Zha L., Fan L., Sun G., Wang H., Ma T., Zhong F., Wei W. Melatonina sensibilizează celulele umane de hepatom la apoptoza indusă de stresul reticulului endoplasmatic. J. Pineal Res. 2012; 52 :322–331. doi: 10.1111/j.1600-079X.2011.00946.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]33.
Fan L., Sun G., Ma T., Zhong F., Lei Y., Li X., Wei W. Melatonina inversează stresul reticulului endoplasmatic indus de tunicamicină în celulele carcinomului hepatocelular uman și îmbunătățește răspunsul citotoxic la doxorubicină prin creșterea CHOP și scăderea Survivin. J. Pineal Res. 2013; 55 :184–194. doi: 10.1111/jpi.12061. [ PubMed ] [ CrossRef ] [ Google Scholar ]34.
Fan L., Sun G., Ma T., Zhong F., Wei W. Melatonina învinge rezistența la apoptoză în carcinomul hepatocelular uman prin țintirea Survivin și XIAP. J. Pineal Res. 2013; 55 :174–183. doi: 10.1111/jpi.12060. [ PubMed ] [ CrossRef ] [ Google Scholar ]35.
Laothong U., Pinlaor P., Boonsiri P., Pairojkul C., Priprem A., Johns NP, Charoensuk L., Intuyod K., Pinlaor S. Melatonina inhibă colangiocarcinomul și reduce leziunile hepatice la Opisthorchis viverrini infectate și N. -hamsteri tratati cu nitrosodimetilamina. J. Pineal Res. 2013; 55 :257–266. doi: 10.1111/jpi.12068. [ PubMed ] [ CrossRef ] [ Google Scholar ]36.
Tomov B., Popov D., Tomova R., Vladov N., Den Otter W., Krastev Z. Răspunsul terapeutic al carcinomului hepatocelular netratabil după aplicarea modulatorilor imunitari IL-2, BCG și melatonina. Anticancer Res. 2013; 33 :4531–4536. [ PubMed ] [ Google Scholar ]37.
Bennukul K., Numkliang S., Leardkamolkarn V. Melatonina atenuează moartea celulelor HepG2 indusă de cisplatină prin reglarea expresiilor mTOR și ERCC1. Lumea J. Hepatol. 2014; 6 :230–242. doi: 10.4254/wjh.v6.i4.230. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]38.
Verma D., Hashim O., Jayapalan J., Subramanian P. Efectul melatoninei asupra statusului antioxidant și a ritmului circadian al activității în timpul hepatocarcinogenezei la șoareci. J. Cancer Res. Acolo. 2014; 10 :1040–1044. doi: 10.4103/0973-1482.138227. [ PubMed ] [ CrossRef ] [ Google Scholar ]39.
Laothong U., Hiraku Y., Oikawa S., Intuyod K., Murata M., Pinlaor S. Melatonina induce apoptoza în liniile celulare de colangiocarcinom prin activarea căii mitocondriale mediate de specii reactive de oxigen. Oncol. Rep. 2015; 33 :1443–1449. doi: 10.3892/or.2015.3738. [ PubMed ] [ CrossRef ] [ Google Scholar ]40.
Colombo J., Wolf Maciel JM, Ferreira LC, Da Silva RF, Pires De Campos Zuccari DA Efectele melatoninei asupra expresiei HIF-1α și VEGF și asupra proprietăților invazive ale celulelor hepatocarcinomului. Oncol. Lett. 2016; 12 :231–237. doi: 10.3892/ol.2016.4605. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]41.
Bu L.-J., Yu H.-Q., Fan L.-L., Li X.-Q., Wang F., Liu J.-T., Zhong F., Zhang C.-J ., Wei W., Wang H., şi colab. Melatonina, un nou inhibitor selectiv de ATF-6, induce apoptoza celulelor hepatomului uman prin reglarea COX-2. Lumea J. Gastroenterol. 2017; 23 :986–998. doi: 10.3748/wjg.v23.i6.986. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]42.
Cheng L., Liu J., Liu Q., Liu Y., Fan L., Wang F., Yu H., Li Y., Bu L., Li X. și colab. Exozomii din celulele de carcinom hepatocelular tratate cu melatonină modifică starea de imunosupresie prin calea STAT3 în macrofage. Int. J. Biol. Sci. 2017; 13 :723–734. doi: 10.7150/ijbs.19642. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]43.
Hao J., Li Z., Zhang C., Yu W., Tang Z., Li Y., Feng X., Gao Y., Liu Q., Huang W., și colab. Direcționarea semnalizării NF-κB/AP-2β pentru a îmbunătăți activitatea antitumorală a cisplatinei de către melatonina în celulele carcinomului hepatocelular. A.m. J. Cancer Res. 2017; 7 :13–27. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]44.
Lin S., Hoffmann K., Gao C., Petrulionis M., Herr I., Schemmer P. Melatonina promovează apoptoza indusă de sorafenib prin activarea sinergică a căii JNK/c-jun în carcinomul hepatocelular uman. J. Pineal Res. 2017; 62 :e12398. doi: 10.1111/jpi.12398. [ PubMed ] [ CrossRef ] [ Google Scholar ]45.
Liu Y., Liu J., Cheng L., Fan L., Wang F., Yu H., Liu Q., Li Y., Bu L., Li X. și colab. Melatonina crește efectele antitumorale ale sorafenibului asupra liniilor celulare de hepatom uman prin autofagie de reglare în jos. Int. J. Clin. Exp. Med. 2017; 10 :14109–14120. [ Google Scholar ]46.
Long F., Dong C., Jiang K., Xu Y., Chi X., Sun D., Liang R., Gao Z., Shao S., Wang L. Melatonina îmbunătățește efectul antitumoral al sorafenibului prin oprirea ciclului celular mediată de AKT/p27 în liniile celulare de hepatocarcinom. RSC Adv. 2017; 7 :21342–21351. doi: 10.1039/C7RA02113E. [ CrossRef ] [ Google Scholar ]47.
Sánchez DI, González-Fernández B., San-Miguel B., de Urbina JO, Crespo I., González-Gallego J., Tuñón MJ Melatonina previne dereglarea căii de semnalizare sfingozin kinazei / sfingozină 1-fosfat la șoarece model de carcinom hepatocelular indus de dietilnitrozamină. J. Pineal Res. 2017; 62 :e12369. doi: 10.1111/jpi.12369. [ PubMed ] [ CrossRef ] [ Google Scholar ]48.
Wongsena W., Charoensuk L., Dangtakot R., Pinlaor P., Intuyod K., Pinlaor S. Melatonina suprimă eozinofilele și celulele Th17 la hamsterii tratați cu o combinație de infecție a ficatului uman și cancerigen chimic. Pharmacol. Rep. 2018; 70 :98–105. doi: 10.1016/j.pharep.2017.07.017. [ PubMed ] [ CrossRef ] [ Google Scholar ]50.
Chen C.-C., Chen C.-Y., Wang S.-H., Yeh C.-T., Su SC, Ueng S.-H., Chuang W.-Y., Hsueh C. , Wang T.-H. Melatonina sensibilizează celulele de carcinom hepatocelular la chimioterapie prin suprimarea reparării ADN-ului mediată de ARN-ul RAD51-AS1, necodificator. Cancers (Basel) 2018; 10 :320. doi: 10.3390/cancers10090320. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]51.
Dauchy RT, Wren-Dail MA, Dupepe LM, Hill SM, Xiang S., Anbalagan M., Belancio VP, Dauchy EM, Blask DE Efectul luminii LED îmbogățite cu albastru de zi asupra inhibării melatoninei circadiane de noapte a hepatomului 7288CTC Warburg efect și progresie. Comp. Med. 2018; 68 :269–279. doi: 10.30802/AALAS-CM-17-000107. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]53.
Wang TH, Hsueh C., Chen CC, Li WS, Yeh CT, Lian JH, Chang JL, Chen CY Melatonina inhibă progresia carcinomului hepatocelular prin reducerea RAF1 mediată de microARN let7i-3p. Int. J. Mol. Sci. 2018; 19 :2687. doi: 10.3390/ijms19092687. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]54.
El-Magd MA, Mohamed Y., El-Shetry ES, Elsayed SA, Abo Gazia M., Abdel-Aleem GA, Shafik NM, Abdo WS, El-Desouki NI, Basyony MA Melatonina maximizează potențialul terapeutic al non- MSC-uri precondiționate într-un model de șobolan indus de DEN de HCC. Biomed. Pharmacother. 2019; 114 :108732. doi: 10.1016/j.biopha.2019.108732. [ PubMed ] [ CrossRef ] [ Google Scholar ]55.
Mohamed Y., Basyony MA, El-Desouki NI, Abdo WS, El-Magd MA Efectul terapeutic potențial pentru melatonina și celulele stem mezenchimale asupra carcinomului hepatocelular. Biomedicina. 2019; 9 :23–29. doi: 10.1051/bmdcn/2019090424. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]56.
Zhang L., He Y., Wu X., Zhao G., Zhang K., Yang CS, Reiter RJ, Zhang J. Melatonina și (-)-epigallocatechin-3-Gallate: Parteneri în lupta împotriva cancerului. Celulele. 2019; 8 :745. doi: 10.3390/cells8070745. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]57.
Zhou B., Lu Q., Liu J., Fan L., Wang Y., Wei W., Wang H., Sun G. Melatonina crește sensibilitatea carcinomului hepatocelular la sorafenib prin calea PERK-ATf4-Beclin1 . Int. J. Biol. Sci. 2019; 15 :1905–1920. doi: 10.7150/ijbs.32550. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]58.
Ao L., Li L., Sun H., Chen H., Li Y., Huang H., Wang X., Guo Z., Zhou R. Analiza transcriptomică asupra efectelor melatoninei în carcinoamele gastrointestinale. BMC Gastroenterol. 2020; 20 :233. doi: 10.1186/s12876-020-01383-z. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]59.
Mi L., Kuang H. Melatonina reglează rezistența la cisplatină și metabolismul glucozei prin semnalizarea hipopotamilor în celulele carcinomului hepatocelular. Managementul cancerului. Res. 2020; 12 :1863–1874. doi: 10.2147/CMAR.S230466. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]60.
Wang R., Liu Y., Mi X., Chen Q., Jiang P., Hou J., Lin Y., Li S., Ji B., Fang Y. Sirt3 promovează sensibilitatea celulelor de carcinom hepatocelular la regorafenib prin accelerarea disfuncției mitocondriale. Arc. Biochim. Biophys. 2020; 689 :108415. doi: 10.1016/j.abb.2020.108415. [ PubMed ] [ CrossRef ] [ Google Scholar ]61.
Moreira AJ, Rodrigues G., Bona S., Cerski CT, Marroni CA, Mauriz JL, González-Gallego J., Marroni NP Stresul oxidativ și deteriorarea celulară într-un model de leziuni precanceroase și carcinom hepatocelular avansat la șobolani. Toxicol. Rep. 2015; 2 :333–340. doi: 10.1016/j.toxrep.2014.11.015. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]62.
Fernández-Palanca P., Fondevila F., Méndez-Blanco C., Tuñón MJ, González-Gallego J., Mauriz JL Efectele antitumorale ale quercetinei în modelele de hepatocarcinom in vitro și in vivo: o revizuire sistematică. Nutrienți. 2019; 11 :2875. doi: 10.3390/nu11122875. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]63.
Pourhanifeh MH, Mehrzadi S., Kamali M., Hosseinzadeh A. Melatonina și cancerele gastrointestinale: dovezi actuale bazate pe căile de semnalizare subiacente. EURO. J. Pharmacol. 2020; 886 :173471. doi: 10.1016/j.ejphar.2020.173471. [ PubMed ] [ CrossRef ] [ Google Scholar ]64.
González-Fernández B., Sánchez DI, Crespo I., San-Miguel B., De Urbina JO, González-Gallego J., Tuñón MJ Melatonina atenuează dereglarea căii de ceas circadian la șoarecii cu fibroză indusă de CCl4 și uman celule stelate hepatice. Față. Pharmacol. 2018; 9 :556. doi: 10.3389/fphar.2018.00556. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ] Retras65.
Crespo I., Fernández-Palanca P., San-Miguel B., Álvarez M., González-Gallego J., Tuñón MJ Melatonina modulează mitofagia, imunitatea înnăscută și ceasurile circadiene într-un model de insuficiență hepatică fulminantă indusă de virus. J. Cell. Mol. Med. 2020; 24 :7625–7636. doi: 10.1111/jcmm.15398. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]66.
Filipski E., Subramanian P., Carrière J., Guettier C., Barbason H., Lévi F. Perturbarea circadiană accelerează carcinogeneza ficatului la șoareci. Mutat. Res. Genet. Toxicol. Mediul. Mutagen. 2009; 680 :95–105. doi: 10.1016/j.mrgentox.2009.10.002. [ PubMed ] [ CrossRef ] [ Google Scholar ]67.
Pfeffer CM, Singh ATK Apoptosis: O țintă pentru terapia anticancer. Int. J. Mol. Sci. 2018; 19 :448. doi: 10.3390/ijms19020448. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]68.
Kim B.-J., Ryu S.-W., Song B.-J. Fosforilarea mediată de JNK și p38 kinază a Bax duce la activarea și translocarea mitocondrială și la apoptoza celulelor HepG2 de hepatom uman. J. Biol. Chim. 2006; 281 :21256–21265. doi: 10.1074/jbc.M510644200. [ PubMed ] [ CrossRef ] [ Google Scholar ]69.
Méndez-Blanco C., Fondevila F., García-Palomo A., González-Gallego J., Mauriz JL Rezistența la sorafenib în hepatocarcinom: Rolul factorilor inducibili de hipoxia. Exp. Mol. Med. 2018; 50 :134. doi: 10.1038/s12276-018-0159-1. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]70.
Fondevila F., Méndez-Blanco C., Fernández-Palanca P., González-Gallego J., Mauriz JL Activitatea antitumorală a tratamentelor cu regorafenib unic și combinat în modelele preclinice ale cancerelor hepatice și gastrointestinale. Exp. Mol. Med. 2019; 51 :109. doi: 10.1038/s12276-019-0308-1. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]71.
Yang S., Yang L., Li X., Li B., Li Y., Zhang X., Ma Y., Peng X., Jin H., Li H. Noi perspective asupra autofagiei în carcinomul hepatocelular: mecanisme si strategii terapeutice. A.m. J. Cancer Res. 2019; 9 :1329–1353. [ Articol gratuit PMC ] [ PubMed ] [ Google Scholar ]72.
Morales A., Lee H., Goñi FM, Kolesnick R., Fernandez-Checa JC Sphingolipides and cell death. Apoptoza. 2007; 12 :923–939. doi: 10.1007/s10495-007-0721-0. [ PubMed ] [ CrossRef ] [ Google Scholar ]73.
Prieto-Domínguez N., Ordóñez R., Fernández A., García-Palomo A., Muntané J., González-Gallego J., Mauriz JL Modulation of autophagy by sorafenib: Effects on treatment response. Față. Pharmacol. 2016; 7 :151. doi: 10.3389/fphar.2016.00151. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]74.
Méndez-Blanco C., Fondevila F., Fernández-Palanca P., García-Palomo A., van Pelt J., Verslype C., González-Gallego J., Mauriz JL Stabilization of hypoxia-inductible factors and BNIP3 promoter metilarea contribuie la rezistența dobândită la sorafenib în celulele hepatocarcinomului uman. cancere. 2019; 11 :1984. doi: 10.3390/cancers11121984. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]75.
Mauriz JL, González-Gallego J. Medicamente antiangiogenice: cunoștințe actuale și noi abordări ale terapiei cancerului. J. Pharm. Sci. 2008; 97 :4129–4154. doi: 10.1002/jps.21286. [ PubMed ] [ CrossRef ] [ Google Scholar ]76.
Morse MA, Sun W., Kim R., He AR, Abada PB, Mynderse M., Finn RS Rolul angiogenezei în carcinomul hepatocelular. Clin. Cancer Res. 2019; 25 :912–920. doi: 10.1158/1078-0432.CCR-18-1254. [ PubMed ] [ CrossRef ] [ Google Scholar ]77.
Mancinelli R., Mammola CL, Sferra R., Pompili S., Vetuschi A., Pannarale L. Role of the angiogenic factors in cholangiocarcinoma. Appl. Sci. 2019; 9 :1393. doi: 10.3390/app9071393. [ CrossRef ] [ Google Scholar ]78.
Jung JE, Lee H., Cho I., Chung DH, Yoon S., Yang YM, Lee JW, Choi S., Park J., Ye S. și colab. STAT3 este un potențial modulator al expresiei VEGF mediată de HIF-1 în celulele de carcinom renal uman. FASEB J. 2005; 19 :1296–1298. doi: 10.1096/fj.04-3099fje. [ PubMed ] [ CrossRef ] [ Google Scholar ]79.
Yu H., Pan C., Zhao S., Wang Z., Zhang H., Wu W. Resveratrolul inhibă expresia metaloproteinazei-9 a matricei mediată de factorul de necroză tumorală și invazia celulelor de carcinom hepatocelular uman. Biomed. Pharmacother. 2008; 62 :366–372. doi: 10.1016/j.biopha.2007.09.006. [ PubMed ] [ CrossRef ] [ Google Scholar ]80.
Scheau C., Badarau IA, Costache R., Caruntu C., Mihai GL, Didilescu AC, Constantin C., Neagu M. The role of matrix metaloproteinases in the epithelial-mesenchymal transition of hepatocellular carcinoma. Anal. Celulă. Pathol. 2019; 2019 :9423907. doi: 10.1155/2019/9423907. [ Articol gratuit PMC ] [ PubMed ] [ CrossRef ] [ Google Scholar ]81.
Reiter RJ, Sharma R., Ma Q. Comutarea celulelor bolnave de la glicoliza aerobă citosolică la fosforilarea oxidativă mitocondrială: un ritm metabolic reglat de melatonină? J. Pineal Res. 2021; 70 :e12677. doi: 10.1111/jpi.12677. [ PubMed ] [ CrossRef ] [ Google Scholar ]82.
Bizarri M., Proietti S., Cucina A., Reiter RJ Mecanismele moleculare ale acțiunilor pro-apoptotice ale melatoninei în cancer: O revizuire. Opinia expertului. Acolo. Ținte. 2013; 17 :1483–1496. doi: 10.1517/14728222.2013.834890. [ PubMed ] [ CrossRef ] [ Google Scholar ]
Articolele de la
Antioxidanți sunt furnizate aici prin amabilitatea
Institutului Multidisciplinar de Publicare Digitală (MDPI)
Scopul acestei revizuiri a fost de a revizui și de a discuta diferite fitochimice care prezintă efecte benefice asupra canalelor membranare mutante și, prin urmare, de a îmbunătăți conductanța transmembranară. Aceste fitochimice terapeutice pot avea potențialul de a reduce mortalitatea și morbiditatea pacienților cu CF. Patru baze de date au fost căutate folosind cuvinte cheie. Au fost identificate studii relevante, iar articolele conexe au fost separate. Google Scholar, precum și literatura gri (adică, informații care nu sunt produse de editori comerciali), au fost, de asemenea, verificate pentru articole similare pentru a localiza/identifica studii suplimentare. Bazele de date relevante au fost căutate a doua oară pentru a se asigura că au fost incluse studii recente. În concluzie, în timp ce curcumina, genisteina și resveratrolul și-au demonstrat eficacitatea în acest sens, trebuie subliniat faptul că cumarinele, quercetina, și alte medicamente pe bază de plante au, de asemenea, efecte benefice asupra funcției transportorului, conductivității transmembranare și activității generale ale canalului. Studii suplimentare in vitro și in vivo ar trebui efectuate pe CFTR mutant pentru a defini fără echivoc mecanismul prin care fitochimicele modifică funcția/activitatea canalului transmembranar, deoarece rezultatele studiilor evaluate în această revizuire au un grad ridicat de eterogenitate și discrepanță. În cele din urmă, cercetările continue să fie întreprinse pentru a defini în mod clar mecanismul(ele) de acțiune și efectele terapeutice pe care fitochimicele terapeutice le au asupra simptomelor observate la pacienții cu FC într-un efort de a reduce mortalitatea și morbiditatea.
1Departamento de Docencia e Investigación, Facultad de Ciencias Médicas, Pontificia Universidad Católica Argentina, Buenos Aires 1007, Argentina
2Centro de Neuropsiquiatría y Neurología de la Conducta, Hospital de Clínicas „José de San Martín”, Facultatea de Medicina, Universidad de Buenos Aires, Buenos Aires 1121, Argentina
Boala Alzheimer (AD) este o problemă majoră de sănătate și există o recunoaștere din ce în ce mai mare că eforturile de prevenire a acesteia trebuie întreprinse atât de organizații guvernamentale, cât și de organizații neguvernamentale. În acest context, produsul pineal, melatonina, are o semnificație promițătoare datorită proprietăților sale cronobiotice/citoprotectoare potențial utile pentru o serie de aspecte ale AD. Una dintre caracteristicile înaintării în vârstă este scăderea treptată a nivelului circulant de melatonina. Un număr limitat de studii terapeutice au indicat că melatonina are o valoare terapeutică ca medicament neuroprotector în tratamentul AD și afectarea cognitivă minimă (care poate evolua către AD). Atât in vitro, cât și in vivo, melatonina a prevenit neurodegenerarea observată în modelele experimentale de AD. Pentru ca aceste efecte să apară, sunt necesare doze de melatonină cu aproximativ două ordine de mărime mai mari decât cele necesare pentru a afecta somnul și ritmicitatea circadiană. Mai recent, atenția s-a concentrat asupra dezvoltării de analogi potenți ai melatoninei cu efecte prelungite, care au fost folosiți în studiile clinice la pacienții cu tulburări de somn sau depresive în doze considerabil mai mari decât cele utilizate pentru melatonina. Având în vedere că potențele relative ale analogilor sunt mai mari decât cele ale compusului natural, studiile clinice care utilizează melatonina în intervalul 50-100 mg/zi sunt urgent necesare pentru a evalua validitatea sa terapeutică în tulburările neurodegenerative precum AD
Abstract grafic
1. Introducere
Tulburările neurodegenerative sunt un grup de boli cronice și progresive caracterizate prin pierderi selective și simetrice ale neuronilor în sistemele cognitive, motorii sau senzoriale. Boala Alzheimer (AD) este reprezentantul cel mai relevant din punct de vedere clinic al acestui tip de tulburări. Deși originea neurodegenerării specifice în AD rămâne nedefinită, trei procese majore și frecvent interconectate, și anume deteriorarea mediată de radicali liberi, disfuncția mitocondrială și excitotoxicitatea, stau la baza mecanismelor fiziopatologice care duc la moartea neuronală [1
] .AD are un impact foarte mare asupra sănătății. Într-o publicație recentă [
2] organizația Alzheimer’s Disease International (ADI) afirmă că 44 de milioane de oameni trăiesc cu demență în prezent și că această cifră va crește la 135 de milioane de oameni până în 2050. Costul economic actual al demenței este de 604 miliarde de dolari anual. Trebuie menționat că noile estimări reprezintă o creștere de 17% față de cifrele publicate în 2009, îmbătrânirea populației fiind principalul motor al creșterii proiectate. Raportul ADI prezice, de asemenea, o schimbare în distribuția poverii globale a demenței până în 2050, 71% dintre toate persoanele afectate trăind în țări cu venituri mici sau medii. Se estimează că 10% din cazurile de demență pot fi evitate prin campanii care vizează obezitatea, hipertensiunea, diabetul, subactivitatea și fumatul, precum și prin educație și îmbunătățirea cognitivă. Prin urmare,Aportul regulat de antioxidanți de către vârstnici a fost una dintre strategiile recomandate pentru prevenirea bolilor neurodegenerative asociate vârstei, mediate de radicali liberi, dar eficacitatea acestui tratament este dezbătută [3
] . În acest context, utilizarea melatoninei ca agent citoprotector devine de mare interes. Melatonina este un metoxiindol bine conservat care se găsește în majoritatea filelor și are acțiuni citoprotectoare remarcabile pe lângă proprietățile cronobiotice. Sursa melatoninei circulante este glanda pineală, iar o cantitate substanțială de date susține că scăderea melatoninei plasmatice este una dintre caracteristicile înaintării în vârstă [
4 ].]. Vom trece în revistă pe scurt câteva aspecte relevante ale biologiei melatoninei în ceea ce privește procesul neurodegenerativ. Apoi, vor fi discutate teoria și modul de acțiune al melatoninei în AD. În cele din urmă, studiile clinice care susțin utilizarea terapeutică a melatoninei în AD vor fi evaluate critic.
2. Biologia de bază a melatoninei relevante pentru neurodegenerare
Melatonina circulantă la mamifere derivă aproape exclusiv din glanda pineală [
5 ]. În plus, melatonina este sintetizată local în multe celule, țesuturi și organe, inclusiv limfocite, măduvă osoasă, timus, tractul gastrointestinal, piele și ochi (vezi [6,
) . Faptul că apariția melatoninei nu este limitată la vertebrate, ci este prezentă aproape omniprezent în numeroși taxoni, inclusiv, de exemplu, bacterii, eucariote unicelulare și plante [6, 8], subliniază că această moleculă a câștigat
suplimentare în cursul evolutiei.Atât la alte animale, cât și la oameni, melatonina participă la diverse funcții fiziologice semnalând nu numai durata nopții, ci și facilitând, printre altele, eliminarea radicalilor liberi și răspunsul imun, prezentând astfel proprietăți citoprotectoare relevante [6
] . Într-adevăr, citoprotecția poate explica bine prezența melatoninei în majoritatea organismelor vii. Funcțiile sistemice și intracelulare ale melatoninei diferă în cantitățile necesare de agonist. În general, efectele melatoninei asupra ritmurilor biologice sunt exercitate la concentrații nanomolare și prin mecanisme mediate de receptor. Efectele citoprotectoare ale melatoninei necesită cu 100–1000 de concentrații mai mari, compatibile cu cantitățile atinse intracelular de melatonina în mai multe țesuturi [
9 ] și este metabolizată în principal în ficat, în principal prin hidroxilare la poziția C6. Acesta este catalizat selectiv de citocromul microzomal hepatic P450 1A2 (CYP1A2), cu contribuții minore din CYP2C19 hepatic și CYP1A1 și CYP1B11A în mare parte extrahepatic [
12 ]. 6-hidroximelatonina formată este apoi conjugată cu sulfat sau glucuronid pentru a fi excretată în urină. Un alt metabolit important este
N -acetilserotonina care se formează prin
O -demetilare și poate reprezenta până la 20% din doza administrată de melatonină [
13 ].Melatonina este, de asemenea, metabolizată în țesuturi prin scindarea inelului pirolic oxidativ în derivați de chinuramină. Produsul primar de scindare este
N1 – acetil-
N2 -formil-5-metoxichinuramina (AFMK), care este deformilată, fie de arilaminformamidază, fie de hemoperoxidază, la
N1
– acetil -5-metoxicinuramină (AMK) [
14 ]. S-a propus că AFMK este metabolitul activ primar al melatoninei pentru a media citoprotecția [
15 ]. Melatonina este, de asemenea, transformată în 3-hidroximelatonină ciclică într-un proces care curăță direct doi radicali hidroxil [
15]. Datorită metabolismului său rapid, melatonina are un timp de înjumătățire scurt de aproximativ o oră, deși există o variație inter-individuală marcată a nivelurilor plasmatice ale melatoninei după administrarea orală [16
2 se găsesc în membrana celulară ca dimeri și heterodimeri. GPR50, un ortolog al receptorului melatoninei cuplat cu proteina G care nu leagă melatonina în sine, dimerizează cu receptorii MT
Receptorul MT2 uman prezintă o afinitate mai mică decât receptorul MT1 uman
şi devine desensibilizat după expunerea la melatonină, probabil prin internalizare.Ca reprezentanți ai familiei de receptori cuplati cu proteina G, receptorii MT
1 și MT
2 acționează printr-un număr de mecanisme de transducție a semnalului [
1 este cuplat la proteinele G care mediază inhibarea adenililciclazei și activarea fosfolipazei C. Receptorul MT2 este, de asemenea, cuplat la inhibarea adenilil ciclazei și, în plus, inhibă calea guanilil ciclază solubilă
.Prin utilizarea autoradiografiei receptorilor cu ligand neselectiv 2-[125I]iodomelatonină și reacție cantitativă în lanț a polimerazei-transcripție inversă în timp real pentru a marca ARNm a receptorului melatoninei, s-au identificat receptorii MT
1 și MT
2 regiuni ale creierului. La nivelul hipocampusului, receptorii MT
2 au fost detectați în neuronii piramidali CA3 și CA4, care primesc inputuri excitatoare glutamatergice din cortexul entorinal, în timp ce receptorii MT
1 au fost exprimați predominant în CA1 [
6 ].Melatonina se leagă, de asemenea, la factorii de transcripție aparținând superfamiliei receptorilor acidului retinoic, în special, variantele de îmbinare ale RORα (RORα1, RORα2 și RORα izoforma d) și RZRβ [21
] . Subformele receptorului acidului retinoic sunt exprimate în mod omniprezent în țesuturile mamiferelor [
22 ].Melatonina este un antioxidant puternic care captează radicalii ∙OH, precum și alte specii de oxigen radical (ROS) și specii de azot radical (RNS) și care dă naștere cascadei de metaboliți menționați mai sus care împărtășesc proprietăți antioxidante [23
] . Melatonina acționează, de asemenea, indirect pentru a promova expresia genică a enzimelor antioxidante și pentru a inhiba expresia genică a enzimelor prooxidante [
27 ]. În special, acest lucru este valabil pentru glutation peroxidază (GPx) și pentru glutation reductază (GRd), probabil ca răspuns la creșterile dependente de GPx ale GSSG, forma oxidată a glutationului (GSH). Melatonina contribuie la menținerea nivelurilor normale de GSH a creierului [
28] prin stimularea biosintezei GSH prin γ-glutamilcisteina sintetază și glucozo-6-fosfat dehidrogenază [
29 ]. Melatonina are o superioritate demonstrată față de vitamina C și E în protecția împotriva daunelor oxidative și în eliminarea radicalilor liberi [
23 ].Melatonina are proprietăți antiinflamatorii semnificative, probabil prin inhibarea legării factorului nuclear κB (NFκB) de ADN, scăzând astfel sinteza citokinelor proinflamatorii prin inhibarea ciclooxigenazei (Cox) [30], în principal
33 ]. Deși acțiunea directă a melatoninei ca agent antioxidant este în mare parte independentă de interacțiunea receptorului [
34 ], reglarea în sus a enzimelor antioxidante de către metoxiindol implică transcripția nucleară și, în unele cazuri, receptorii RZR/RORα [
35] .].Eficacitatea melatoninei în inhibarea daunelor oxidative a fost testată într-o varietate de modele de boli neurologice. În plus față de modelele animale de AD discutate mai jos, s-a demonstrat că melatonina reduce daunele neuronale din cauza toxicității cadmiului [
51].Proprietățile neuroprotectoare ale melatoninei, precum și efectele sale reglatoare asupra tulburărilor circadiene, validează beneficiile acesteia ca substanță terapeutică în tratamentul preventiv al AD. Mai mult, melatonina exercită efecte anti-excitatoare și, la doze suficiente, efecte sedative [
53 ], astfel că poate exista un al doilea mod de acțiune neuroprotector care implică sistemul ergic de acid y-aminobutiric (GABA) ca mediator. Acest punct de vedere este susținut de studii care indică faptul că melatonina protejează neuronii de toxicitatea peptidei amiloid-β (Aβ) (o neurotoxină principală implicată în AD) prin activarea receptorilor GABA [54
] .Studiile timpurii privind acțiunile anti-excitotoxice ale melatoninei au folosit kainatul, un agonist al receptorilor ionotropi de glutamat și au susținut ipoteza că melatonina previne moartea neuronală indusă de aminoacizii excitatori [55
] . De asemenea, s-a raportat că administrarea de melatonină reduce leziunile neuronilor CA1 hipocampali cauzate de ischemia tranzitorie a creierului anterior [
59 ].Diferitele tipuri de toxicități enumerate mai sus pot duce la moarte neuronală prin necroză sau apoptoză. Moartea neuronală apoptotică necesită sinteza de ARN și proteine și epuizarea factorilor trofici. Apoptoza implică, de asemenea, rupturi monocatenar ale ADN-ului și s-a descoperit că factorii neurotrofici salvează neuronii de acest tip de moarte [
60 ]. Ele pot acționa prin intermediul componentelor celulare anti-apoptotice, cum ar fi proteina proto-oncogenă a limfomului B (Bcl-2).
Studiile in vitro indică faptul că melatonina îmbunătățește expresia Bcl-2 și previne apoptoza [
66].Procesul de îmbătrânire al creierului este cunoscut în prezent suficient de detaliat [
67 ]. O serie de procese de senescență se bazează pe deteriorarea mitocondrială, inclusiv apoptoza prin peroxidarea cardiolipinei, eliberarea citocromului C și defalcarea mtPTP și reducerea masei mitocondriale. Blocarea lanțului de transport de electroni duce la o aprovizionare insuficientă cu energie și la afectarea funcției și viabilității celulelor. Generarea crescută de ROS și RNS provoacă leziuni concomitente la endotelii, proteinele ADN și lipide. În plus, o supraexcitare neuronală cu supraîncărcare cu calciu și activarea microgliei apar odată cu vârsta. Ca o consecință a deteriorării ADN-ului, are loc o uzură crescută a telomerilor împreună cu o reducere a celulelor cu capacitate proliferativă ridicată, cum ar fi celulele progenitoare imune, ducând astfel la imunosenescență [67]
.]. Această situație este însoțită de creșterea răspunsurilor autoimune și de un proces inflamator care duce la creșterea citokinelor proinflamatorii.Dovezile derivate din studiile pe animale indică faptul că melatonina poate reduce majoritatea aspectelor îmbătrânirii creierului [
67].]. În primul rând, prin acțiunea asupra oscilatorilor circadieni centrali și periferici, melatonina crește amplitudinile ritmului deprimat și coordonarea slabă a ritmurilor observate la persoanele în vârstă. Melatonina afectează expresia proteinei de ceas și poate modula perturbarea sensibilității metabolice în senescență. Prin susținerea fluxului de electroni mitocondriali și reducerea debordării electronilor, melatonina îmbunătățește eficiența respiratorie și aprovizionarea cu energie și previne apoptoza. Reducerea ROS și RNS exercitată prin eliminarea radicalilor, reglarea în sus a enzimelor antioxidante, inhibarea enzimelor prooxidante, creșterea GSH și formarea de radicali mai mici duce la o deteriorare redusă a proteinelor, lipidelor și ADN-ului, împreună cu inhibarea telomerilor induși de oxidanți. uzură și supraexcitare neuronală. Acțiunile antiinflamatorii ale melatoninei sunt cruciale în reducerea proceselor legate de îmbătrânire, precum și în îmbunătățirea sensibilității la insulină și a contracarării sindromului metabolic atribuit metoxiindolului. În cele din urmă, melatonina modulează imunitatea naturală și adaptivă și îmbunătățește imunosenescența, parțial printr-un număr mai mare de celule cu capacitate proliferativă ridicată, inclusiv leucocite, celule stem și celule progenitoare.
3. Prezentare generală a terapiei cu melatonina pentru boala Alzheimer – Teorie și mod de acțiune
Plăcile senile extracelulare, formate în principal din depozite de Aβ, și încurcăturile neurofibrilare intracelulare, rezultate în principal din proteina asociată microtubulilor (MAP) anormal hiperfosforilată, sunt caracteristica patologică majoră a AD. Ap joacă un rol important în promovarea degenerării neuronale în AD transformând neuronii vulnerabili la creșterile legate de vârstă ale nivelurilor de stres oxidativ și un metabolism energetic celular alterat.Aβ este compus din 39–43 de resturi de aminoacizi derivate din precursorul său, proteina precursor amiloid (APP) [
68 ]. APP este procesat proteolitic de către α- sau β-secretaze în diferite căi. Calea α-non-amiloidogenă implică scindarea APP de către α-secretază pentru a elibera un fragment
N -terminal al APP, care după scindarea de către γ-secretază împiedică formarea Ap [
68 ]. Calea β-amiloidogenă include β-secretaza, care are ca rezultat formarea peptidei Aβ intacte și este mediată de scindarea secvenţială a β-secretazei și γ-secretazei la N- și C-terminal al
secvenţei Aβ
[ 68
] .Melatonina a inhibat nivelurile normale de secreție de APP solubilă în diferite linii celulare interferând cu maturarea APP [
69 ]. În plus, administrarea melatoninei reduce eficient generarea și depunerea Ap
74 ]. În general, rezultatele la șoarecii transgenici susțin opinia conform căreia melatonina ar trebui administrată într-o fază timpurie pentru a regla metabolismul APP și Aβ în principal prin prevenirea formării lor, cu un efect anti-amiloid redus ulterior. Astfel, terapia cu melatonină la șoarecii bătrâni Tg2576 începând cu vârsta de 14 luni nu a putut preveni depunerea suplimentară de Aβ.
75 ] în timp ce un tratament similar începând cu vârsta de a patra lună a fost eficient pentru a o reduce [
70 ]. Deoarece patologia plăcii de amiloid este observată în mod obișnuit la șoarecii Tg2576 cu vârsta de 10 până la 12 luni [
76 ], datele indică eficacitatea melatoninei în prevenirea formării timpurii a plăcii de amiloid, mai degrabă decât ulterior.Modul în care melatonina își exercită efectul inhibitor asupra generării de Ap rămâne nedefinit. Scindarea proteolitică a APP de către calea α-secretazei este reglată de mulți stimuli fiziologici și patologici, în special prin activarea proteinei kinazei (PK) C și clivajul mediat de secretază a APP. Inhibarea glicogen sintazei kinazei-3 (GSK-3) și reglarea în sus a kinazei
N -terminale c-Jun au ca rezultat o activitate ridicată a metaloproteinazelor matriceale cu degradarea crescândă a Ap [
77 ]. Activitatea enzimei de degradare a insulinei (IDE) care reglează nivelurile de insulină, Aβ și APP, a scăzut după creșterea Apei [
78] .]. GSK-3 interacționează cu presenilin-1, un cofactor al y-secretazei, fosforilarea GSK-3 de către PKC ducând la inactivarea y-secretazei. Într-adevăr, GSK-3 poate fi una dintre căile de semnalizare comune, crescând generarea de Ap și hiperfosforilarea tau, iar melatonina ar putea regla procesarea APP prin căile PKC și GSK-3 (
80 ]. Această interacțiune între melatonină și Aβ pare să depindă mai degrabă de caracteristicile structurale ale melatoninei decât de proprietățile sale antioxidante, deoarece nu ar putea fi imitată de alți captatori de radicali liberi [
79 ]. Prin blocarea formării foilor secundare, melatonina nu numai că reduce neurotoxicitatea, ci și facilitează clearance-ul peptidei, crescând degradarea proteolitică, de exemplu, prin IDE.
Figura 1. Efectul melatoninei asupra semnalizării afectate a insulinei cerebrale în AD. Figura schematizează procesele care leagă deficiența insulină/receptorul de insulină cu plăcile senile și încurcăturile neurofibrilare. Între paranteze, acțiunea demonstrată a melatoninei, așa cum este discutată în text.Stresul oxidativ joacă un rol central în neurotoxicitatea indusă de Ap și moartea celulelor. Acumularea datelor susține că melatonina protejează eficient celulele împotriva daunelor oxidative induse de Aβ și a morții celulare
85.]. În celulele și animalele tratate cu Ap, melatonina și-ar putea exercita activitatea protectoare printr-un efect antioxidant, în timp ce în celulele transfectate cu APP și modelele animale transgenice, mecanismul de bază poate implica în primul rând inhibarea generării de foi de β și/sau fibrile de amiloid. Ap agregat generează ROS care produc moartea neuronală prin deteriorarea lipidelor membranei neuronale, proteinelor și acizilor nucleici. S-a observat protecție împotriva toxicității Aβ de către melatonină, în special la nivel mitocondrial [
89 ]. Amploarea patologiei neurofibrilare se corelează cu severitatea demenței la pacienții cu AD. Nivelul de tau hiperfosforilat este de trei până la patru ori mai mare în creierul pacienților cu AD decât în creierul adulților normali [
91 ].Melatonina atenuează eficient hiperfosforilarea tau prin afectarea protein kinazelor și fosfatazelor într-un număr de modele experimentale, inclusiv expunerea celulelor neuroblastomului N2a și SH-SY5Y la wortmannin [92], calyculin
101 ]. Inhibarea biosintezei melatoninei la șobolani nu numai că a dus la afectarea memoriei spațiale, dar a indus și o creștere a fosforilării tau, un efect prevenit de suplimentarea cu melatonina [102]
.].Se știe că stresul oxidativ influențează starea de fosforilare a tau [
104 ]. Acumularea de proteine greșite și agregate în neuronii creierului ai AD este considerată o consecință a stresului oxidativ, în plus față de modificările structurale moleculare datorate vârstei [
105 ]. Deoarece melatonina previne, ca antioxidant și captator de radicali liberi, supraproducția de radicali liberi, pare fezabil ca prevenirea fosforilării tau de către melatonina să aibă loc parțial datorită activității sale antioxidante. În plus, mai multe studii au indicat că melatonina poate acționa ca un modulator al enzimelor într-un mod care nu are legătură cu proprietățile sale antioxidante. Acestea includ reglarea de către melatonina a PKA [
120 ].Un alt eveniment major în patogeneza AD este deficitul funcției colinergice [
121 ]. Neuronii din nucleul bazal al lui Meynert, principala sursă de inervație colinergică a cortexului cerebral și a hipocampului, suferă o degenerare profundă și selectivă în creierul AD [
122 ]. Nivelurile de acetilcolină (ACh) sunt reduse în stadiul incipient al AD, în timp ce activitățile enzimelor de sinteză și degradare a colin acetiltransferaza (ChAT) și acetilcolinesteraza (AChE) nu se modifică până la o fază târzie a AD [123
] . Deoarece scăderea activității ChAT în neocortexul pacienților cu AD sa corelat cu severitatea demenței, inhibitorii AChE au devenit un tratament standard al AD ușoară până la moderată [125]
.].Melatonina are un efect protector asupra sistemului colinergic. Previne inhibarea indusă de peroxinitrit a transportului de colină și a activității ChAT în sinaptozomi și vezicule sinaptice [
126 ]. Tratamentul cu melatonină al șoarecilor transgenici APP695 în vârstă de opt luni a îmbunătățit semnificativ reducerea profundă a activității ChAT în cortexul frontal și hipocampus [
81 ]. Melatonina antagonizează, de asemenea, deficitul de memorie spațială și scăderea activității ChAT găsite la șobolanii adulți ovariectomizați [
127 ]. Cu toate acestea, la șobolanii perfuzați intracerebroventricular cu Aβ timp de 14 zile, melatonina nu a putut restabili activitatea ChAT [
128 ]. Melatonina a inhibat creșterea indusă de lipopolizaharide și streptozotocină a activității AChE [
129]. Recent, hibrizi ai inhibitorului AChE tacrină și melatonină au fost sintetizați ca noi candidați la medicamente pentru tratarea AD [
131 ]. Acești hibrizi au prezentat o activitate antioxidantă și de conservare colinergică mai bună decât tacrina sau melatonina în monoterapie. Administrarea directă intracerebrală a unuia dintre acești hibrizi a scăzut moartea celulară indusă și încărcarea Aβ în parenchimul creierului de șoarece APP/PS1, însoțită de o recuperare a funcției cognitive [
131 ].Există un interes din ce în ce mai mare pentru rolul semnalizării insulinei cerebrale afectate în patologia AD. S-a sugerat că perturbarea semnalizării creierului insulinei/factorului de creștere asemănător insulinei 1 (IGF-1) este un eveniment cauzal cheie care stă la baza AD, fiind legată atât de prezența plăcilor senile, cât și a încurcurilor neurofibrilare [132
] . Acest punct de vedere, însă, nu este universal susținut [
135 ]. O semnalizare afectată a insulinei/receptorului de insulină (IR) duce la scăderea activării mediate de insulină a activității de semnalizare fosfoinozitid 3-kinaze (PI-3K)/Akt, ducând la supraactivarea GSK-3 care promovează direct hiperfosforilarea tau și acumularea de Aβ și plăcile senile. formarea (
Figura 1). Activitatea IDE, care reglează nivelurile de insulină, Aβ și APP, a scăzut după depunerea de Aβ [
78 ]. Administrarea melatoninei reduce semnele sindromului metabolic, cum ar fi hiperglicemia, dislipidemia, hiperinsulinemia, rezistența la insulină, creșterea în greutate și hipertensiunea [
137 ]. Melatonina restabilește mecanismele receptorilor de insulină/insulină și crește activitatea de semnalizare a fosfoinozitidei 3-kinaze/Akt cu o inhibare rezultată a GSK-3 și mai puțină acumulare de Aβ și hiperfosforilare tau (vezi [ 6 )
.]). În plus, întreruperea semnalizării insulinei duce la o scădere a expresiei transportorului de glucoză-1 (GLUT-1) și -3 (GLUT-3), culminând cu absorbția/metabolismul glucozei cerebrale afectate, un alt eveniment contracarat de melatonină. Metabolismul neuronal restabilit al glucozei mărește acilarea tau
N -acetilglucozamină, reducând astfel hiperfosforilarea tau (
Figura 2 rezumă principalele ținte posibile pentru medicație în AD. Food and Drug Administration (FDA) a aprobat doar inhibitori de AChE și blocanți ai receptorilor
N -metil-
D -aspartat (NMDA) pentru uz clinic. După cum sa discutat mai sus, melatonina are proprietatea unică de a afecta toate mecanismele fiziopatologice descrise în
Figura 2 . Prin urmare, potențialul său ca neuroprotector în AD merită să fie explorat.
Figura 2. Ținte posibile pentru medicație în AD. După cum se discută în text, melatonina are potențialul de a afecta toate mecanismele descrise în figură. (*) Aprobat de FDA.
4. Aspecte clinice ale aplicației melatoninei în AD
La pacienții cu dementa, severitatea deficienței mintale s-a corelat semnificativ cu producerea de melatonină nocturnă afectată [ 138 ]. Acesta poate fi un semn precoce al bolii, deoarece nivelurile de melatonina din lichidul cefalorahidian (LCR) sunt reduse la pacienții cu AD autopsiați în stadiul preclinic, stadiul Braak-1. Prin urmare, reducerea melatoninei LCR poate fi un marker pentru detectarea precoce a AD [ 139 ].În plus, nivelurile plasmatice ale melatoninei sunt mai scăzute la pacienții cu AD [ 140 , 141 , 142 , 143 ] și s-au postulat că conexiunile retiniene-SCN-pineale afectate funcțional sunt cauza acestei scăderi [ 144]. Expresia și activitatea genei monoaminoxidazei au crescut, iar nivelurile de ARNm a adrenoceptorului β1 au scăzut, în glanda pineală a pacienților cu AD [ 145 ]. În plus, au apărut modificări în imunoreactivitatea receptorului de melatonină în hipocampul AD [ 146 , 147 ]. În mod colectiv, rezultatele indică faptul că înlocuirea nivelurilor de melatonină poate fi o strategie terapeutică pentru oprirea progresiei AD.„Apusul soarelui”, un fenomen cronobiologic care include gândirea dezorganizată, capacitatea redusă de a menține atenția la stimulii externi, rătăcirea, agitația și tulburările perceptuale și emoționale, este o situație observată frecvent la pacienții cu AD. Ca fenomen legat de timpul zilei, care apare după-amiaza târziu sau seara devreme [
149 ], posibilul efect terapeutic al melatoninei a fost întreținut.
Tabelul 1 rezumă datele care indică faptul că melatonina, ca agent cronobiotic, poate fi eficientă în tratarea apusului soarelui și a ciclurilor neregulate de somn-veghe la pacienții cu AD.O revizuire a rezultatelor publicate privind utilizarea melatoninei în AD [ 150 ] a dat opt rapoarte (cinci studii deschise, două rapoarte de caz) ( N = 89 de pacienți) care susțin o posibilă eficacitate a melatoninei: calitatea somnului s-a îmbunătățit și la pacienții cu apus de soare. a fost redusă și decăderea cognitivă a încetinit progresia. În șase studii dublu-orb, randomizate controlate cu placebo, au fost examinați un număr total de 210 de pacienți cu AD. Calitatea somnului a crescut și apusul soarelui a scăzut semnificativ, iar performanța cognitivă s-a îmbunătățit în patru studii ( N = 143), în timp ce a existat absența efectelor în două studii ( N = 67) [ 150 ].O altă căutare sistematică a studiilor publicate între 1985 și aprilie 2009 privind melatonina și apusul soarelui la pacienții cu AD a fost publicată [ 151 ]. Toate lucrările despre tratamentul cu melatonina în demență au fost preluate și efectele melatoninei asupra tulburărilor de ritm circadian au fost notate prin evaluarea comportamentului de apus/agitat, calitatea somnului și funcționarea în timpul zilei. Au fost revizuite un total de 9 lucrări, inclusiv 4 studii randomizate controlate ( N = 243) și 5 serii de cazuri ( N = 87) ( Tabelul 1 ). Două dintre studiile controlate randomizate au constatat o îmbunătățire semnificativă a comportamentului de apus/agitat. Toate cele cinci serii de cazuri au găsit o îmbunătățire [ 151]. Astfel, dacă melatonina are vreo valoare în tratarea AD rămâne incertă. Probabil, eterogenitatea grupului examinat în AD complet dezvoltat face dificilă dezvăluirea oricărui efect terapeutic în studiile care includ un număr mic de pacienți [
152 ]. Mai mult, expresia hipocampală redusă a receptorilor de melatonină MT 2 la pacienții cu AD [ 146 ] și a receptorilor MT 1 din aparatul circadian poate explica de ce tratamentul cu melatonina este neregulat în acest stadiu avansat al bolii [ 153 , 154 ].
Tabel 1. Studii care includ tratamentul pacienților cu boala Alzheimer (AD) cu melatonină.
Deficiența cognitivă ușoară (MCI) este denumirea dată semnelor și simptomelor diagnosticate la cei care au un deficit obiectiv și măsurabil în funcțiile cognitive, dar cu o relativă conservare a activităților zilnice. Ratele anuale de conversie la AD sunt de până la 10%–15% [
169 ] și MCI reprezintă o etapă importantă clinic pentru identificarea și tratarea persoanelor cu risc. Într-adevăr, procesul degenerativ din creierul AD începe cu 20-30 de ani înainte de debutul clinic al AD [
173 .]. După cum sa menționat mai sus, nivelurile de melatonina LCR scad în stadiile preclinice ale AD, ceea ce sugerează că reducerea melatoninei LCR poate fi un declanșator și un marker precoce pentru AD [ 139 , 145 ]. Deși nu se știe dacă deficiența relativă de melatonină este fie o consecință, fie o cauză a neurodegenerării, pierderea de melatonină probabil agravează boala.Am raportat anterior o analiză retrospectivă în care 3-9 mg zilnic dintr-un preparat de melatonină cu eliberare rapidă per os (po) la culcare timp de până la trei ani au îmbunătățit semnificativ performanța cognitivă și emoțională și ciclul zilnic de somn/veghe la 25 de pacienți cu MCI [174] . Recent am raportat date de la o altă serie de 96 de pacienți cu MCI, dintre care 61 au primit zilnic 3-24 mg de un preparat de melatonină cu eliberare rapidă po, la culcare, timp de 15 până la 60 de luni, în comparație cu un grup similar de 35 de pacienți cu MCI care nu au primit.[ 175 ]. În plus, toți pacienții au primit medicația standard individuală considerată adecvată de către medicul psihiatru curant.Pacienții tratați cu melatonină au prezentat performanțe semnificativ mai bune la mini-examenul stării mentale și la subscala cognitivă a scalei de evaluare a AD. După aplicarea unei baterii neuropsihologice care cuprinde un test Mattis, testul Digit-simbol, sarcinile Trail A și B și testul verbal Rey, s-au găsit performanțe mai bune la pacienții tratați cu melatonină pentru fiecare parametru testat [175 ] . Scorurile anormal de mari ale Beck Depression Inventory au scăzut la pacienții tratați cu melatonină, concomitent cu îmbunătățirea calității somnului și a stării de veghe [ 175 ]. Aceste rezultate susțin în continuare că melatonina este un medicament suplimentar util pentru tratarea MCI într-un mediu clinic.Astfel, o inițiere precoce a tratamentului cu melatonină poate fi decisivă pentru succesul terapeutic [ 75 ]. În Tabelul 2 , sunt rezumate datele publicate privind tratamentul cu melatonină în MCI. Șase studii dublu-orb, randomizate controlate cu placebo și două studii retrospective deschise ( N = 782) au arătat în mod constant că administrarea zilnică de melatonină seara îmbunătățește calitatea somnului și performanța cognitivă la pacienții cu MCI. Prin urmare, tratamentul cu melatonină ar putea fi eficient în stadiile incipiente ale bolii neurodegenerative.Există două motive pentru care este benefică utilizarea melatoninei la pacienții cu MCI. În cursul procesului neurodegenerativ, deteriorarea legată de vârstă în organizarea circadiană devine semnificativ exacerbată și este responsabilă de probleme de comportament precum apusul soarelui [
176 ]. Declinul cognitiv legat de vârstă la adulții în vârstă sănătoși poate fi prezis prin fragmentarea ritmului circadian în comportamentul locomotor. Prin urmare, înlocuirea nivelurilor scăzute de melatonină care apar în creier [ 139 , 145 ] poate fi foarte benefică la pacienții cu MCI.Pe de altă parte, cea mai mare parte a informațiilor despre proprietățile neuroprotectoare ale melatoninei derivate din studii experimentale (vezi pentru [ 177 , 178 ]) face ca este foarte de dorit să se utilizeze doze farmacologice la pacienții cu MCI cu scopul de a opri sau încetini progresia bolii. Deși glanda pineală secretă melatonina care circulă în sânge și LCR [ 179 ], date recente susțin ipoteza că, la ovine și probabil la oameni, numai melatonina LCR, și nu melatonina din sânge, poate furniza cea mai mare parte a melatoninei cerebrale. țesut în concentrații mari [ 180]. Au fost demonstrati gradienti de concentratie semnificativi orientati de la ventricul (aproape de LCR) catre tesutul cerebral, cu concentratii variind cu un factor de la 1 la 125. Prin urmare, pe lângă susținerea rolului LCR în disponibilitatea fiziologică a melatoninei [ 179 ], aceste rezultate implică faptul că trebuie administrate cantități mari, farmacologice de melatonină pentru a avea acces la creier.Mecanismele care explică efectul terapeutic al melatoninei la pacienții cu MCI rămân de definit. Tratamentul cu melatonina promovează în principal somnul cu unde lente la vârstnici [
181 ] și poate fi benefic în MCI prin creșterea fazelor de restaurare ale somnului, inclusiv secreția crescută de GH și neurotrofine.După cum sa subliniat mai sus, melatonina acționează la diferite niveluri relevante pentru dezvoltarea și manifestarea AD. Efectele antioxidante, mitocondriale și antiamiloidogenice pot interfera eventual cu debutul bolii. Prin urmare, momentul în care începe tratamentul cu melatonină poate fi decisiv pentru răspunsul final [ 75 ].Un aspect important care trebuie luat în considerare este doza de melatonină folosită, care poate fi inutil de scăzută atunci când se iau în considerare afinitățile de legare, timpul de înjumătățire și potențele relative ale diferiților agoniști de melatonină de pe piață. Pe lângă faptul că sunt în general mai puternici decât molecula nativă, analogii melatoninei sunt folosiți în cantități considerabil mai mari [ 182 ]. Dozele autorizate de ramelteon agonist al receptorului melatoninei variază de la 8 la 32 mg/zi, în timp ce agomelatina a fost autorizată pentru tratamentul tulburării depresive majore în doze de 25-50 mg/zi. În studiile clinice care implică subiecți umani sănătoși, tasimelteon (Vanda Pharmaceuticals, Washington, DC, SUA), un alt agonist al receptorului de melatonină aprobat recent de FDA este administrat în doze de 20 până la 100 mg/zi [ 183 .] în timp ce farmacocinetica, farmacodinamia și siguranța agonistului receptorului de melatonină TIK-301 (Tikvah Pharmaceuticals, Atlanta, GA, SUA) au fost examinate într-un studiu controlat cu placebo utilizând 20 până la 100 mg/zi [184 ] . Prin urmare, studiile MCI cu doze de melatonină în intervalul 100-300 mg/zi sunt justificate în continuare.O terapie combinată a melatoninei cu un agonist al receptorului melatoninei ar putea fi teoretic benefică pentru pacienții cu AD, mai ales dacă agonistul receptorului melatoninei are un profil farmacocinetic mai bun decât melatonina. Cu toate acestea, agoniştii receptorilor melatoninei au demonstrat beneficii limitate în modelele murine de AD [ 185 ].Deoarece melatonina este eliminată foarte rapid din fluxul sanguin, strategiile cum ar fi inhibarea degradării melatoninei și eliminarea din organism ar putea fi un supliment util la tratamentul cu melatonina exogenă. Într-adevăr, nivelurile plasmatice farmacologice ale melatoninei ar putea fi afectate ca urmare a expunerii concomitente la substanțe chimice care modulează expresia CYP1A2. De exemplu, fluvoxamina, un inhibitor puternic al CYP1A2 și, într-o măsură mai mică, al CYP2C19, a crescut nivelurile plasmatice de melatonina [
187 ]. De asemenea, consumul concomitent de cofeină al cărei metabolism este catalizat în principal de CYP1A2, a dublat nivelurile plasmatice și a crescut biodisponibilitatea melatoninei [
188] .]. Un alt candidat este 5-metoxipsoralen, un medicament utilizat pentru tratamentul psoriazisului, care crește nivelurile plasmatice ale melatoninei endogene și exogene [
191 ].Trebuie subliniat faptul că melatonina are un profil ridicat de siguranță, este de obicei remarcabil de bine tolerată și, în unele studii, a fost administrată la pacienți în doze foarte mari [16 , 192 , 193, 194 195 ] . Melatonina (300 mg/zi) timp de până la trei ani a scăzut stresul oxidativ la pacienții cu scleroză laterală amiotrofică [ 192 ].La copiii cu distrofie musculară, 70 mg/zi de melatonină au redus citokinele și peroxidarea lipidelor [ 193 ]. Doze de 80 mg melatonină pe oră timp de 4 ore au fost administrate bărbaților sănătoși, fără alte reacții adverse în afară de somnolență [ 16 ]. La femeile sănătoase, cărora li sa administrat 300 mg melatonină/zi timp de patru luni, nu au existat efecte secundare [ 194 ]. Un studiu clinic randomizat, controlat, dublu-orb, pe 50 de pacienți trimiși pentru o intervenție chirurgicală hepatică, a indicat că o singură doză enterală preoperatorie de 50 mg/kg melatonină (adică, echivalentul a 3 g pentru un adult de 60 kg) a fost sigură și bine tolerată.
195 ].Un alt rezultat al studiului raportat recent [ 175 ] a fost că atunci când se folosește melatonina, sunt necesare mult mai puține benzodiazepine pentru a trata tulburările de somn în MCI. Deoarece, după cum s-a menționat mai sus, melatonina și benzodiazepinele au împărtășit unele proprietăți neurochimice (
adică , interacțiunea cu mecanismele mediate de GABA în creier [ 196 ]) și comportamentale (de exemplu, o activitate anxiolitică similară dependentă de zi [ 46 ]), terapia cu melatonină a fost postulată să să fie un instrument eficient pentru a scădea doza de benzodiazepine necesară la pacienți [
199]. O analiză retrospectivă recentă a unei baze de date germane de prescripții a identificat 512 pacienți care au inițiat tratament cu melatonină cu eliberare prelungită (2 mg) pe o perioadă de 10 luni [200
] . Din 112 pacienți din acest grup care au utilizat anterior benzodiazepine, 31% au întrerupt tratamentul cu benzodiazepine la trei luni după începerea tratamentului cu melatonină cu eliberare prelungită. Rata de întrerupere a fost mai mare la pacienții care au primit două sau trei rețete de melatonină [
200 ]. Utilizarea prelungită a benzodiazepinelor și agoniștilor receptorilor de benzodiazepină (
Z-droguri) este legată de simptome severe de sevraj și de dependență potențială, care a devenit o problemă de sănătate publică, ducând la mai multe campanii de scădere a consumului acestor medicamente. Un studiu farmacoepidemiologic recent a concluzionat că aceste campanii au eșuat în general atunci când nu au fost asociate cu disponibilitatea și piața melatoninei [
În concluzie, întrebarea dacă melatonina are o valoare terapeutică în prevenirea sau tratarea MCI, care afectează inițierea bolii sau progresia neuropatologiei și mecanismele implicate, merită să fie analizată în continuare. Sunt necesare studii multicentrice dublu-orb pentru a explora și investiga potențialul și utilitatea melatoninei ca medicament antidemență în stadiul incipient al bolii.Deoarece melatonina prezintă atât proprietăți hipnotice, cât și cronobiotice, a fost utilizată terapeutic pentru tratamentul insomniei legate de vârstă, precum și al altor insomnii primare și secundare [
203 ]. Un consens al Asociației Britanice pentru Psihofarmacologie privind tratamentul bazat pe dovezi al insomniei, parasomniei și tulburărilor de somn de ritm circadian a concluzionat că melatonina în doză de 2 mg este tratamentul de primă alegere atunci când un hipnotic este indicat la pacienții cu vârsta peste 55 de ani [
204 ] .După cum arată afinitățile de legare, timpul de înjumătățire și potențele relative ale diferiților agoniști ai melatoninei de pe piață, este clar că studiile care utilizează aceste doze mici de melatonină nu sunt adecvate pentru a oferi o comparație adecvată cu efectul compușilor menționați mai sus, care, în plus pentru a fi în general mai puternice decât molecula nativă sunt folosite în cantități considerabil mai mari [
205 ]. Prin urmare, sunt necesare studii suplimentare care folosesc doze mai mari de melatonină pentru a clarifica potențialele sale implicații neuroprotectoare terapeutice la om. Din studiile pe animale, este clar că o serie de efecte preventive ale melatoninei, cum ar fi cele în tulburările neurodegenerative, au nevoie de doze mari de melatonină pentru a deveni evidente [
177] .]. Dacă ne așteptăm ca melatonina să fie un neuroprotector eficient, este probabil ca dozele mici de melatonină folosite până acum să nu fie foarte benefice.
Tabel 2. Studii care includ tratamentul pacienților cu deficiență cognitivă ușoară (MCI) cu melatonină.
Abrevieri
Ach
acetilcolina
Durere
acetilcolinesteraza
ANUNȚ
Boala Alzheimer
ADI
Alzheimer Disease International
AFMK
N1 – acetil- N2 – formil-5-metoxicinuramină
Akt
protein kinaza identificată în virusul AKT
AMK
N1 – acetil-5-metoxicinuramină
APP
proteine precursoare de amiloid
Ap
β-amiloid agregat
Bcl-2
Proteina proto-oncogenă a limfomului cu celule B
Conversație
colin acetiltransferaza
Cox
ciclooxigenaza
LCR
fluid cerebrospinal
CYP1A1
citocromul P450 1A1
CYP1A2
citocromul P450 1A2
CYP2C19
citocromul P450 2C19
CYP1B1
citocromul P450 1B1
GABA
acid y-aminobutiric
GPR50
G-protein receptor 50 ortholog
GLUT-1
transportor de glucoză-1
GLUT-3
transportor de glucoză-3
GPx
glutation peroxidază
GRd
glutation reductază
GSH
glutation redus
GSK-3
glicogen sintază kinaza-3
IDE
enzimă degradatoare de insulină
iNOS
sintaza de oxid nitric inductibil
IGF-1
Factorul de creștere asemănător insulinei 1
HARTĂ
proteine asociate microtubulilor
MCI
deficit cognitiv minor
mPTP
porul de tranziție al permeabilității mitocondriale
ARNm
acid ribonucleic mesager
MT 1
receptor de melatonina 1
MT 2
receptor de melatonina 2
NFkB
factorul nuclear κB
NMDA
N -metil- d -aspartat
nNOS
oxid nitric sintază neuronală
NU
oxid de azot
NSAD
medicamente antiinflamatoare nesteroidiene
PI3-K
fosfoinozitid 3-kinaza
PK
protein kinaza
REM
miscare rapida a ochilor
RNS
specii reactive de azot
ROR
receptorul orfan legat de receptorul acidului retinoic
ROS
specii reactive de oxigen
RZR
receptorul retinoid Z
SCN
nuclei suprachiasmatici
GAZON
superoxid dismutaza
Mulțumiri
Studiile în laboratoarele autorilor au fost susținute de granturi de la Agencia Nacional de Promoción Científica y Tecnológica, Argentina și Universitatea din Buenos Aires. DPC este un laureat pentru cariera de cercetare de la Consiliul de Cercetare din Argentina (CONICET) și profesor emerit, Universitatea din Buenos Aires. DEV și LIB sunt premiați pentru cariera de cercetare de la CONICET.
Contribuții ale autorului
DPC a scris schița generală a recenziei. Toți ceilalți autori au citit și au fost de acord cu versiunea.
Conflicte de interes
Autorii nu declară niciun conflict de interese.
Referințe
Reiter, RJ; Garcia, JJ; Pie, J. Toxicitate oxidativă în modele de neurodegenerare: răspunsuri la melatonina. Restaurare. Neurol. Neurosci. 1998 , 12 , 135–142. [ Google Scholar ] [ PubMed ]
Johnson, EJ; Vishwanathan, R.; Johnson, MA; Hausman, DB; Davey, A.; Scott, TM; Verde, RC; Miller, LS; Gearing, M.; Woodard, J.; et al. Relația dintre carotenoizii serici și cerebrali, concentrațiile de α-tocoferol și retinol și performanța cognitivă la cel mai vechi vechi din studiul Georgia Centenarian. J. Aging Res. 2013 , 2013 . [ Google Scholar ] [ CrossRef ]
Bubenik, GA; Konturek, SJ Melatonina și îmbătrânirea: perspective pentru tratamentul uman. J. Physiol Pharmacol. 2011 , 62 , 13–19. [ Google Scholar ] [ PubMed ]
Claustrat, B.; Brun, J.; Chazot, G. Fiziologia de bază și patofiziologia melatoninei. Sleep Med. Rev. 2005 , 9 , 11–24. [ Google Scholar ] [ CrossRef ]
Venegas, C.; Garcia, JA; Escames, G.; Ortiz, F.; Lopez, A.; Doerrier, C.; Garcia-Corzo, L.; Lopez, LC; Reiter, RJ; Acuña-Castroviejo, D. Melatonina extrapineală: Analiza distribuției sale subcelulare și a fluctuațiilor zilnice. J. Pineal Res. 2012 , 52 , 217–227. [ Google Scholar ] [ CrossRef ]
Paredes, SD; Korkmaz, A.; Manchester, LC; Tan, DX; Reiter, RJ Phytomelatonin: O revizuire. J. Exp. Bot. 2009 , 60 , 57–69. [ Google Scholar ] [ PubMed ]
Cardinali, DP; Lynch, HJ; Wurtman, RJ Legarea melatoninei de proteinele plasmatice umane și de șobolan. Endocrinologie 1972 , 91 , 1213–1218. [ Google Scholar ] [ CrossRef ]
Ma, X.; Idle, JR; Krausz, KW; Gonzalez, FJ Metabolismul melatoninei de către citocromii umani p450. Drug Metab. Dispos. 2005 , 33 , 489–494. [ Google Scholar ] [ CrossRef ]
Facciola, G.; Hidestrand, M.; von Bahr, C.; Tybring, G. Izoformele citocromului P450 implicate în metabolismul melatoninei în microzomii ficatului uman. EURO. J. Clin. Pharmacol. 2001 , 56 , 881–888. [ Google Scholar ] [ CrossRef ]
Skene, DJ; Papagiannidou, E.; Hashemi, E.; Snelling, J.; Lewis, DF; Fernandez, M.; Ioannides, C. Contribuția CYP1A2 în metabolismul hepatic al melatoninei: Studii cu preparate microzomale izolate și felii de ficat. J. Pineal Res. 2001 , 31 , 333–342. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Young, IM; Leone, RM; Francis, P.; Stovell, P.; Silman, RE Melatonina este metabolizată în N -acetil serotonină și 6-hidroximelatonină la om. J. Clin. Endocrinol. Metab. 1985 , 60 , 114–119. [ Google Scholar ] [ CrossRef ]
Hardeland, R.; Tan, DX; Reiter, RJ Kynuramines, metaboliți ai melatoninei și alți indoli: învierea unei clase aproape uitate de amine biogene. J. Pineal Res. 2009 , 47 , 109–126. [ Google Scholar ] [ CrossRef ]
Tan, DX; Manchester, LC; Terron, deputat; Flores, LJ; Reiter, RJ O moleculă, mulți derivați: o interacțiune fără sfârșit a melatoninei cu speciile reactive de oxigen și azot? J. Pineal Res. 2007 , 42 , 28–42. [ Google Scholar ] [ CrossRef ]
Waldhauser, F.; Waldhauser, M.; Lieberman, HR; Deng, MH; Lynch, HJ; Wurtman, RJ Biodisponibilitatea melatoninei orale la om. Neuroendocrinology 1984 , 39 , 307–313. [ Google Scholar ] [ CrossRef ]
Aldhous, M.; Franey, C.; Wright, J.; Arendt, J. Concentrațiile plasmatice de melatonina la om în urma absorbției orale a diferitelor preparate. Br. J. Clin. Pharmacol. 1985 , 19 , 517–521. [ Google Scholar ] [ CrossRef ]
Fourtillan, JB; Brisson, AM; Gobin, P.; Ingrand, I.; Decourt, JP; Girault, J. Biodisponibilitatea melatoninei la om după administrarea de zi a melatoninei D7 . Biopharm. Eliminarea medicamentelor. 2000 , 21 , 15–22. [ Google Scholar ] [ CrossRef ]
Dubocovich, ML; Delgrange, P.; Krause, DN; Sugden, D.; Cardinali, DP; Olcese, J. Uniunea Internațională de Farmacologie de bază și clinică. LXXV. Nomenclatura, clasificarea și farmacologia receptorilor de melatonină cuplați cu proteina G. Pharmacol. Rev. 2010 , 62 , 343–380. [ Google Scholar ] [ CrossRef ]
Levoye, A.; Dam, J.; Ayoub, MA; Guillaume, JL; Couturier, C.; Delgrange, P.; Jockers, R. Receptorul orfan GPR 50 inhibă în mod specific funcția receptorului de melatonină MT 1 prin heterodimerizare. EMBO J. 2006 , 25 , 3012–3023. [ Google Scholar ] [ CrossRef ]
Wiesenberg, I.; Missbach, M.; Kahlen, JP; Schrader, M.; Carlberg, C. Activarea transcripțională a receptorului nuclear RZR alfa de către melatonina, hormonul glandei pineale și identificarea CGP 52608 ca ligand sintetic. Acizi nucleici Res. 1995 , 23 , 327–333. [ Google Scholar ] [ CrossRef ]
Lardone, PJ; Guerrero, JM; Fernandez-Santos, JM; Rubio, A.; Martin-Lacave, I.; Carrillo-Vico, A. Melatonina sintetizată de limfocitele T ca ligand al receptorului orfan legat de acidul retinoic. J. Pineal Res. 2011 , 51 , 454–462. [ Google Scholar ] [ CrossRef ]
Galano, A.; Tan, DX; Reiter, RJ Melatonina ca un aliat natural împotriva stresului oxidativ: o examinare fizico-chimică. J. Pineal Res. 2011 , 51 , 1–16. [ Google Scholar ] [ CrossRef ]
Antolin, I.; Rodriguez, C.; Sainz, RM; Mayo, JC; Uria, H.; Kotler, ML; Rodriguez-Colunga, MJ; Tolivia, D.; Menendez-Pelaez, A. Neurohormone melatonina previne deteriorarea celulelor: efect asupra expresiei genelor pentru enzimele antioxidante. FASEB J. 1996 , 10 , 882–890. [ Google Scholar ] [ PubMed ]
Pablos, MI; Reiter, RJ; Ortiz, GG; Guerrero, JM; Agapito, MT; Chuang, JI; Sewerynek, E. Ritmurile glutation peroxidazei și glutation reductazei în creierul puiului și inhibarea lor de către lumină. Neurochema. Int. 1998 , 32 , 69–75. [ Google Scholar ] [ CrossRef ]
Rodriguez, C.; Mayo, JC; Sainz, RM; Antolin, I.; Herrera, F.; Martin, V.; Reiter, RJ Reglarea enzimelor antioxidante: un rol semnificativ pentru melatonina. J. Pineal Res. 2004 , 36 , 1–9. [ Google Scholar ] [ CrossRef ]
Jimenez-Ortega, V.; Cano, P.; Cardinali, DP; Esquifino, AI Variația de 24 de ore în expresia genelor a enzimelor căii redox în hipotalamusul de șobolan: Efectul tratamentului cu melatonina. Redox Rep. 2009 , 14 , 132–138. [ Google Scholar ] [ CrossRef ]
Subramanian, P.; Mirunalini, S.; Pandi-Perumal, SR; Trakht, I.; Cardinali, DP Tratamentul cu melatonina imbunatateste statutul antioxidant si scade continutul de lipide din creier si ficat la sobolani. EURO. J. Pharmacol. 2007 , 571 , 116–119. [ Google Scholar ] [ CrossRef ]
Kilanczyk, E.; Bryszewska, M. Efectul melatoninei asupra enzimelor antioxidante în fibroblastele pielii diabetice umane. Celulă. Mol. Biol. Lett. 2003 , 8 , 333–336. [ Google Scholar ] [ PubMed ]
Cardinali, DP; Ritta, MN; Fuentes, AM; Gimeno, MF; Gimeno, AL Eliberarea de prostaglandină E de către hipotalamusul bazal medial de șobolan in vitro . Inhibarea de către melatonină la concentrații submicromolare. EURO. J. Pharmacol. 1980 , 67 , 151–153. [ Google Scholar ] [ CrossRef ]
Deng, WG; Tang, ST; Tseng, HP; Wu, KK Melatonina suprimă macrofagele ciclooxigenazei-2 și expresia inductibilă a oxidului nitric sintazei prin inhibarea acetilării și legarii p52. Blood 2006 , 108 , 518–524. [ Google Scholar ] [ CrossRef ]
Costantino, G.; Cuzzocrea, S.; Mazzon, E.; Caputi, AP Efectele protectoare ale melatoninei în inflamația labei indusă de plasmă activată de zimosan. EURO. J. Pharmacol. 1998 , 363 , 57–63. [ Google Scholar ] [ CrossRef ]
Tan, D.; Reiter, RJ; Chen, LD; Poeggeler, B.; Manchester, LC; Barlow-Walden, LR Atât nivelurile fiziologice, cât și cele farmacologice ale melatoninei reduc formarea aductului ADN indusă de safrolul cancerigen. Carcinogenesis 1994 , 15 , 215–218. [ Google Scholar ] [ CrossRef ]
Leon-Blanco, MM; Guerrero, JM; Reiter, RJ; Pozo, D. Expresia ARN a subunităților de telomerază umană TR și TERT este afectată diferențial de agoniştii receptorului de melatonină din linia celulară tumorală MCF-7. Cancer Lett. 2004 , 216 , 73–80. [ Google Scholar ] [ CrossRef ]
Urata, Y.; Honma, S.; Mergi la S.; Todoroki, S.; Iida, T.; Cho, S.; Honma, K.; Kondo, T. Melatonina induce sintetaza y-glutamilcisteină mediată de proteina activatoare-1 în celulele endoteliale vasculare umane. Radic liber. Biol. Med. 1999 , 27 , 838–847. [ Google Scholar ] [ CrossRef ]
Poliandri, AH; Esquifino, AI; Cano, P.; Jimenez, V.; Lafuente, A.; Cardinali, DP; Duvilanski, BH Efectul protector in vivo al melatoninei asupra modificărilor induse de cadmiu în echilibrul redox și expresia genelor în hipotalamusul de șobolan și pituitara anterioară. J. Pineal Res. 2006 , 41 , 238–246. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Jimenez-Ortega, V.; Cano, P.; Scacchi, PA; Cardinali, DP; Esquifino, AI Perturbarea indusă de cadmiu în expresia de 24 de ore a genelor de ceas și enzime redox în hipotalamusul bazal medial de șobolan. Prevenirea prin melatonină. Față. Neurol. 2011 , 2 . [ Google Scholar ] [ CrossRef ]
Shaikh, AY; Xu, J.; Wu, Y.; El, L.; Hsu, CY Melatonina protejează celulele endoteliale cerebrale bovine de deteriorarea ADN-ului indusă de hiperoxie și de moarte. Neurosci. Lett. 1997 , 229 , 193–197. [ Google Scholar ] [ CrossRef ]
Pablos, MI; Reiter, RJ; Chuang, JI; Ortiz, GG; Guerrero, JM; Sewerynek, E.; Agapito, MT; Melchiorri, D.; Lawrence, R.; Deneke, SM Melatonina administrată acut reduce daunele oxidative în plămâni și creier induse de oxigenul hiperbaric. J. Apl. Physiol. 1997 , 83 , 354–358. [ Google Scholar ] [ PubMed ]
Princ, FG; Juknat, AA; Maxit, AG; Cardalda, C.; Batlle, A. Protecția antioxidantă a melatoninei împotriva daunelor oxidative induse de acidul δ-aminolevulinic în cerebelul șobolanului. J. Pineal Res. 1997 , 23 , 40–46. [ Google Scholar ] [ CrossRef ]
Carneiro, RC; Reiter, RJ Peroxidarea lipidelor indusă de acidul δ-aminolevulinic în rinichi și ficat de șobolan este atenuată de melatonină: un studiu in vitro și in vivo . J. Pineal Res. 1998 , 24 , 131–136. [ Google Scholar ] [ CrossRef ]
Onuki, J.; Almeida, EA; Medeiros, MH; Di, MP Inhibarea deteriorării ADN-ului induse de acidul 5-aminolevulinic de către melatonină, N1 – acetil- N2 – formil-5-metoxicinuramină, quercetină sau resveratrol. J. Pineal Res. 2005 , 38 , 107–115. [ Google Scholar ] [ CrossRef ]
Erol, FS; Topsakal, C.; Ozveren, MF; Kaplan, M.; Ilhan, N.; Ozercan, IH; Yildiz, OG Efectele protectoare ale melatoninei și vitaminei E în leziunile cerebrale datorate radiațiilor gamma: un studiu experimental. Neurochirurgie. Rev. 2004 , 27 , 65–69. [ Google Scholar ] [ CrossRef ]
Shirazi, A.; Haddadi, GH; Asadi-Amoli, F.; Sakhaee, S.; Ghazi-Khansari, M.; Avand, A. Efectul radioprotector al melatoninei în reducerea stresului oxidativ în lentilele de șobolan. Cell J. 2011 , 13 , 79–82. [ Google Scholar ] [ PubMed ]
Taysi, S.; Memisogullari, R.; Koc, M.; Yazici, AT; Aslankurt, M.; Gumustekin, K.; Al, B.; Ozabacigil, F.; Yilmaz, A.; Tahsin, OH Melatonina reduce stresul oxidativ din lentila de șobolan din cauza leziunii oxidative induse de radiații. Int. J. Radiat. Biol. 2008 , 84 , 803–808. [ Google Scholar ] [ CrossRef ]
Lee, EJ; Wu, TS; Lee, MEA; Chen, TY; Tsai, YY; Chuang, JI; Chang, GL Tratamentul întârziat cu melatonină îmbunătățește recuperarea electrofiziologică după ischemia cerebrală focală tranzitorie la șobolani. J. Pineal Res. 2004 , 36 , 33–42. [ Google Scholar ] [ CrossRef ]
Tai, SH; Hung, YC; Lee, EJ; Lee, AC; Chen, TY; Shen, CC; Chen, HY; Lee, MEA; Huang, SY; Wu, TS Melatonina protejează împotriva ischemiei cerebrale focale tranzitorii atât la femelele active din punct de vedere reproductiv, cât și la femelele cu deficit de estrogen: impactul estrogenului circulant asupra răspunsului său hormetic la doză. J. Pineal Res. 2011 , 50 , 292–303. [ Google Scholar ] [ CrossRef ]
Beni, SM; Kohen, R.; Reiter, RJ; Tan, DX; Shohami, E. Neuroprotecția indusă de melatonină după leziuni ale capului închis este asociată cu creșterea antioxidanților din creier și activarea atenuată în fază târzie a NF-kB și AP-1. FASEB J. 2004 , 18 , 149–151. [ Google Scholar ] [ PubMed ]
Tsai, MC; Chen, WJ; Tsai, MS; Ching, CH; Chuang, JI Melatonina atenuează insulta oxidativă indusă de contuzia cerebrală, inactivarea traductorilor de semnal și a activatorilor transcripției 1 și reglarea în sus a supresoarelor de semnalizare a citokinei-3 la șobolani. J. Pineal Res. 2011 , 51 , 233–245. [ Google Scholar ] [ CrossRef ]
Kabadi, SV; Maher, TJ Posttratamentul cu uridină și melatonină în urma unei leziuni cerebrale traumatice reduce edemul în diferite regiuni ale creierului la șobolani. Ann. NY Acad. Sci. 2010 , 1199 , 105–113. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Reiter, RJ; Manchester, LC; Tan, DX Neurotoxine: Mecanisme de radicali liberi si protectie cu melatonina. Curr. Neurofarmacol. 2010 , 8 , 194–210. [ Google Scholar ] [ CrossRef ]
Golombek, DA; Pevet, P.; Cardinali, DP Efectele melatoninei asupra comportamentului: posibilă mediere de către sistemul central GABAergic. Neurosci. Comportament biologic. Rev. 1996 , 20 , 403–412. [ Google Scholar ] [ CrossRef ]
Caumo, W.; Levandovski, R.; Hidalgo, MP Efectul anxiolitic preoperator al melatoninei și clonidinei asupra durerii postoperatorii și a consumului de morfină la pacienții supuși histerectomiei abdominale: un studiu dublu-orb, randomizat, controlat cu placebo. J. Pain 2009 , 10 , 100–108. [ Google Scholar ] [ CrossRef ]
Louzada, PR; Paula Lima, AC; Mendonca-Silva, DL; Noel, F.; de Mello, FG; Ferreira, ST Taurina previne neurotoxicitatea agoniștilor receptorilor beta-amiloid și glutamat: activarea receptorilor GABA și posibile implicații pentru boala Alzheimer și alte tulburări neurologice. FASEB J. 2004 , 18 , 511–518. [ Google Scholar ] [ CrossRef ]
Giusti, P.; Lipartiti, M.; Franceschini, D.; Schiavo, N.; Floreani, M.; Manev, H. Neuroprotecția prin melatonina de la excitotoxicitatea indusă de kainat la șobolani. FASEB J. 1996 , 10 , 891–896. [ Google Scholar ] [ PubMed ]
Manev, H.; Uz, T.; Kharlamov, A.; Cagnoli, CM; Franceschini, D.; Giusti, P. Protecția in vivo împotriva apoptozei induse de kainat de către hormonul pineal melatonina: Efectul melatoninei exogene și al ritmului circadian. Restaurare. Neurol. Neurosci. 1996 , 9 , 251–256. [ Google Scholar ] [ PubMed ]
Cho, S.; Joh, TH; Baik, HH; Dibinis, C.; Volpe, BT Administrarea melatoninei protejează neuronii hipocampali CA1 după ischemia tranzitorie a creierului anterior la șobolani. Brain Res. 1997 , 755 , 335–338. [ Google Scholar ] [ CrossRef ]
Kilic, E.; Ozdemir, YG; Bolay, H.; Kelestimur, H.; Dalkara, T. Pinealectomia agravează iar administrarea melatoninei atenuează afectarea creierului în ischemia focală. J. Cereb. Fluxul de sânge Metab. 1999 , 19 , 511–516. [ Google Scholar ] [ PubMed ]
Furio, AM; Fontao, R.; Falco, N.; Ruiz, JI; Caccuri, RL; Cardinali, DP Efectul neuroprotector al melatoninei asupra toxicității glucocorticoizilor în hipocampul de șobolan. Deschide Physiol. J. 2008 , 1 , 23–27. [ Google Scholar ] [ CrossRef ]
Dodd, S.; Maes, M.; Anderson, G.; Dean, OM; Moylan, S.; Berk, M. Putative neuroprotective agents in neuropsihiatric disorders. Prog. Neuropsihofarmacol. Biol. Psihiatrie 2013 , 42 , 135–145. [ Google Scholar ] [ CrossRef ]
Jiao, S.; Wu, MM; Hu, CL; Zhang, ZH; Mei, YA 2-iodomelatonina agonistul receptorului de melatonina previne apoptoza neuronilor granulelor cerebeloase prin inhibarea curentului K + . J. Pineal Res. 2004 , 36 , 109–116. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Koh, PO Melatonina previne reglarea în jos a fosfoproteinei astrocitare PEA-15 în leziunile cerebrale ischemice. J. Pineal Res. 2011 , 51 , 381–386. [ Google Scholar ] [ CrossRef ]
Radogna, F.; Diederich, M.; Ghibelli, L. Melatonin: O moleculă pleiotropă care reglează inflamația. Biochim. Pharmacol. 2010 , 80 , 1844–1852. [ Google Scholar ] [ CrossRef ]
Peng, TI; Hsiao, CW; Reiter, RJ; Tanaka, M.; Lai, YK; Jou, MJ mtDNA T8993G Inhibarea complexului mitocondrial V indusă de mutația MJ mărește modificările dependente de cardiolipină ale dinamicii mitocondriale în timpul insultelor oxidative, Ca 2+ și lipidice la cibrizii NARP: o țintă potențială terapeutică pentru melatonină. J. Pineal Res. 2012 , 52 , 93–106. [ Google Scholar ] [ CrossRef ]
Jou, MJ Melatonina păstrează tranziția tranzitorie a permeabilității mitocondriale pentru protecție în timpul stresului mitocondrial de Ca 2+ în astrocite. J. Pineal Res. 2011 , 50 , 427–435. [ Google Scholar ] [ CrossRef ]
Andrabi, SA; Sayeed, eu; Siemen, D.; Wolf, G.; Horn, TF Inhibarea directă a porului de tranziție a permeabilității mitocondriale: un posibil mecanism responsabil pentru efectele anti-apoptotice ale melatoninei. FASEB J. 2004 , 18 , 869–871. [ Google Scholar ] [ PubMed ]
Hardeland, R. Melatonina și teoriile îmbătrânirii: o evaluare critică a rolului melatoninei în mecanismele antiîmbătrânire. J. Pineal Res. 2013 , 55 , 325–356. [ Google Scholar ] [ PubMed ]
Selkoe, DJ Biologia celulară a plierii greșite a proteinelor: exemplele bolilor Alzheimer și Parkinson. Nat. Cell Biol. 2004 , 6 , 1054–1061. [ Google Scholar ] [ CrossRef ]
Lahiri, DK; Ghosh, C. Interacțiuni între melatonină, specii reactive de oxigen și oxid nitric. Ann. NY Acad. Sci. 1999 , 893 , 325–330. [ Google Scholar ] [ CrossRef ]
Matsubara, E.; Bryant-Thomas, T.; Pacheco, QJ; Henry, TL; Poeggeler, B.; Herbert, D.; Cruz-Sanchez, F.; Chyan, YJ; Smith, MA; Perry, G.; et al. Melatonina crește supraviețuirea și inhibă patologia oxidativă și amiloidă într-un model transgenic al bolii Alzheimer. J. Neurochem. 2003 , 85 , 1101–1108. [ Google Scholar ] [ CrossRef ]
Lahiri, DK; Chen, D.; Ge, YW; Bondy, SC; Sharman, EH Suplimentarea alimentară cu melatonină reduce nivelul de β-peptide amiloide în cortexul cerebral murin. J. Pineal Res. 2004 , 36 , 224–231. [ Google Scholar ] [ CrossRef ]
Song, W.; Lahiri, DK Melatonina modifică metabolismul proteinei precursoare beta-amiloid din linia celulară neuroendocrină PC12. J. Mol. Neurosci. 1997 , 9 , 75–92. [ Google Scholar ] [ CrossRef ]
Olivieri, G.; Hess, C.; Savaskan, E.; Ly, C.; Meier, F.; Baysang, G.; Brockhaus, M.; Muller-Spahn, F. Melatonina protejează celulele neuroblastomului SHSY5Y de stresul oxidativ indus de cobalt, neurotoxicitatea și secreția crescută de β-amiloid. J. Pineal Res. 2001 , 31 , 320–325. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Quinn, J.; Kulhanek, D.; Nowlin, J.; Jones, R.; Pratico, D.; Rokach, J.; Stackman, R. Terapia cu melatonina cronica nu reuseste sa modifice sarcina de amiloid sau deteriorarea oxidativa la soarecii vechi Tg2576: Implicatii pentru studiile clinice. Brain Res. 2005 , 1037 , 209–213. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Hsiao, K.; Chapman, P.; Nilsen, S.; Eckman, C.; Harigaya, Y.; Younkin, S.; Yang, F.; Cole, G. Deficiențe de memorie corelative, creșterea Apei și plăci de amiloid la șoarecii transgenici. Science 1996 , 274 , 99–102. [ Google Scholar ] [ CrossRef ]
Donnelly, PS; Caragounis, A.; Du, T.; Laughton, KM; Volitakis, I.; Cherny, RA; Sharples, RA; Hill, AF; Li, QX; Masters, CL; et al. Eliberarea intracelulară selectivă a ionilor de cupru și zinc din complecșii bis (thiosemicarbazonato) reduce nivelurile peptidei amiloid-β ale bolii Alzheimer. J. Biol. Chim. 2008 , 283 , 4568–4577. [ Google Scholar ] [ CrossRef ]
Farris, W.; Mansourian, S.; Chang, Y.; Lindsley, L.; Eckman, EA; Frosch, deputat; Eckman, CB; Tanzi, RE; Selkoe, DJ; Guenette, S. Enzima de degradare a insulinei reglează nivelurile de insulină, β-proteinei amiloid și domeniul intracelular al proteinei precursoare de β-amiloid in vivo . Proc. Natl. Acad. Sci. SUA 2003 , 100 , 4162–4167. [ Google Scholar ] [ CrossRef ]
Poeggeler, B.; Miravalle, L.; Zagorski, MG; Wisniewski, T.; Chyan, YJ; Zhang, Y.; Shao, H.; Bryant-Thomas, T.; Vidal, R.; Frangione, B.; et al. Melatonina inversează activitatea profibrilogenă a apolipoproteinei E4 asupra peptidei amiloid Aβ Alzheimer. Biochimie 2001 , 40 , 14995–15001. [ Google Scholar ] [ CrossRef ]
Pappolla, M.; Bozner, P.; Soto, C.; Shao, H.; Robakis, NK; Zagorski, M.; Frangione, B.; Ghiso, J. Inhibarea β-fibrilogenezei Alzheimer de către melatonina. J. Biol. Chim. 1998 , 273 , 7185–7188. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Feng, Z.; Chang, Y.; Cheng, Y.; Zhang, BL; Qu, ZW; Qin, C.; Zhang, JT Melatonina atenuează deficitele comportamentale asociate cu apoptoza și disfuncția sistemului colinergic în modelul de șoarece transgenic APP 695 al bolii Alzheimer. J. Pineal Res. 2004 , 37 , 129–136. [ Google Scholar ] [ CrossRef ]
Zatta, P.; Tognon, G.; Carampin, P. Melatonina previne formarea radicalilor liberi datorită interacțiunii dintre peptidele β-amiloide și ionii metalici [Al III , Zn II , Cu II , Mn II , Fe II ]. J. Pineal Res. 2003 , 35 , 98–103. [ Google Scholar ] [ CrossRef ]
Furio, AM; Cutrera, RA; Castillo Thea, V.; Perez, LS; Riccio, P.; Caccuri, RL; Brusco, LL; Cardinali, DP Efectul melatoninei asupra modificărilor ritmului activității locomotorii la hamsterii sirieni injectați cu peptida β-amiloid 25-35 în nucleele suprachiasmatice. Celulă. Mol. Neurobiol. 2002 , 22 , 699–709. [ Google Scholar ] [ CrossRef ]
Shen, YX; Xu, SY; Wei, W.; Soare, XX; Yang, J.; Liu, LH; Dong, C. Melatonina reduce modificările de memorie și daunele oxidative neuronale la șoarecii tratați cu d -galactoză. J. Pineal Res. 2002 , 32 , 173–178. [ Google Scholar ] [ CrossRef ]
Rosales-Corral, S.; Tan, DX; Reiter, RJ; Valdivia-Velazquez, M.; Martinez-Barboza, G.; Acosta-Martinez, JP; Ortiz, GG Melatonina administrată oral reduce stresul oxidativ și citokinele proinflamatorii induse de peptida amiloid-β în creierul de șobolan: un studiu comparativ, in vivo versus vitamina C și E. J. Pineal Res. 2003 , 35 , 80–84. [ Google Scholar ] [ CrossRef ]
Olcese, JM; Cao, C.; Mori, T.; Mamcarz, MB; Maxwell, A.; Runfeldt, MJ; Wang, L.; Zhang, C.; Lin, X.; Zhang, G.; et al. Protecție împotriva deficitelor cognitive și a markerilor neurodegenerării prin administrarea orală pe termen lung a melatoninei într-un model transgenic al bolii Alzheimer. J. Pineal Res. 2009 , 47 , 82–96. [ Google Scholar ] [ CrossRef ]
Dragicevic, N.; Copes, N.; O’Neal-Moffitt, G.; Jin, J.; Buzzeo, R.; Mamcarz, M.; Tan, J.; Cao, C.; Olcese, JM; Arendash, GW; et al. Tratamentul cu melatonina restabilește funcția mitocondrială la șoarecii Alzheimer: un rol protector mitocondrial al semnalizării receptorilor membranei melatoninei. J. Pineal Res. 2011 , 51 , 75–86. [ Google Scholar ] [ CrossRef ]
Brion, JP; Anderton, BH; Authelet, M.; Dayanandan, R.; Leroy, K.; Lovestone, S.; Octave, JN; Pradier, L.; Touchet, N.; Tremp, G. Încurcăturile neurofibrilare și fosforilarea tau. Biochim. Soc. Symp. 2001 , 81–88. [ Google Scholar ]
Billingsley, ML; Kincaid, RL Fosforilarea și defosforilarea reglementate a proteinei tau: Efecte asupra interacțiunii microtubulilor, a traficului intracelular și a neurodegenerării. Biochim. J. 1997 , 323 , 577–591. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Khatoon, S.; Grundke-Iqbal, I.; Iqbal, K. Nivelurile creierului de proteină tau asociată cu microtubuli sunt crescute în boala Alzheimer: un test radioimuno-slot-blot pentru nanogramele proteinei. J. Neurochem. 1992 , 59 , 750–753. [ Google Scholar ] [ CrossRef ]
Iqbal, K.; Alonso, AC; Chen, S.; Chohan, MO; El-Akkad, E.; Gong, CX; Khatoon, S.; Li, B.; Liu, F.; Rahman, A.; et al. Patologia Tau în boala Alzheimer și alte tauopatii. Biochim. Biophys. Acta 2005 , 1739 , 198–210. [ Google Scholar ] [ CrossRef ]
Deng, YQ; Xu, GG; Duan, P.; Zhang, Q.; Wang, JZ Efectele melatoninei asupra hiperfosforilării tau induse de wortmannin. Acta Pharmacol. Păcat. 2005 , 26 , 519–526. [ Google Scholar ] [ CrossRef ]
Li, SP; Deng, YQ; Wang, XC; Wang, YP; Wang, JZ Melatonina protejează celulele neuroblastomului SH-SY5Y de afectarea neurofilamentului indusă de caliculina A și neurotoxicitatea. J. Pineal Res. 2004 , 36 , 186–191. [ Google Scholar ] [ CrossRef ]
Li, XC; Wang, ZF; Zhang, JX; Wang, Q.; Wang, JZ Efectul melatoninei asupra hiperfosforilării tau induse de calyculin A. EURO. J. Pharmacol. 2005 , 510 , 25–30. [ Google Scholar ] [ CrossRef ]
Xiong, YF; Chen, Q.; Chen, J.; Zhou, J.; Wang, HX Melatonina reduce afectarea transportului axonal și axonopatia indusă de calyculin A. J. Pineal Res. 2011 , 50 , 319–327. [ Google Scholar ] [ CrossRef ]
Benitez-King, G.; Tunez, I.; Bellon, A.; Ortiz, GG; Anton-Tay, F. Melatonina previne modificările citoscheletice și stresul oxidativ indus de acidul okadaic în celulele N1E-115. Exp. Neurol. 2003 , 182 , 151–159. [ Google Scholar ] [ CrossRef ]
Montilla-Lopez, P.; Munoz-Agueda, MC; Feijoo, LM; Munoz-Castaneda, JR; Bujalance-Arenas, I.; Tunez-Finana, I. Comparația melatoninei versus vitamina C asupra stresului oxidativ și activității enzimelor antioxidante în boala Alzheimer indusă de acid okadaic în celulele neuroblastomului. EURO. J. Pharmacol. 2002 , 451 , 237–243. [ Google Scholar ] [ CrossRef ]
Montilla, P.; Feijoo, M.; Munoz, MC; Munoz-Castaneda, JR; Bujalance, I.; Tunez, I. Efectul melatoninei asupra stresului oxidativ în celulele N1E-115 nu este mediat de receptorii MT1. J. Physiol. Biochim. 2003 , 59 , 263–268. [ Google Scholar ] [ CrossRef ]
Liu, SJ; Wang, JZ Fosforilarea tau asemănătoare Alzheimer indusă de wortmannin in vivo și atenuarea acesteia de către melatonină. Acta Pharmacol. Păcat. 2002 , 23 , 183–187. [ Google Scholar ] [ PubMed ]
Wang, XC; Zhang, J.; Yu, X.; Han, L.; Zhou, ZT; Zhang, Y.; Wang, JZ Prevenirea hiperfosforilării tau indusă de izoproterenol de către melatonină la șobolan. Sheng Li Xue Bao 2005 , 57 , 7–12. [ Google Scholar ] [ PubMed ]
Zhu, LQ; Wang, SH; Ling, ZQ; Wang, DL; Wang, JZ Efectul inhibării biosintezei melatoninei asupra reținerii memoriei spațiale și fosforilării tau la șobolan. J. Pineal Res. 2004 , 37 , 71–77. [ Google Scholar ] [ CrossRef ]
Gomez-Ramos, A.; Diaz-Nido, J.; Smith, MA; Perry, G.; Avila, J. Efectul produsului de peroxidare a lipidelor acroleină asupra fosforilării tau în celulele neuronale. J. Neurosci. Res. 2003 , 71 , 863–870. [ Google Scholar ] [ CrossRef ]
Lovell, MA; Xiong, S.; Xie, C.; Davies, P.; Markesbery, WR Inducerea tau hiperfosforilată în culturile de neuroni corticali primari de șobolan mediată de stresul oxidativ și glicogen sintaza kinaza-3. J. Alzheimer Dis. 2004 , 6 , 659–671. [ Google Scholar ] [ PubMed ]
Kenyon, CJ Genetica îmbătrânirii. Natura 2010 , 464 , 504–512. [ Google Scholar ] [ CrossRef ]
Schuster, C.; Williams, LM; Morris, A.; Morgan, PJ; Barrett, P. Receptorul uman de melatonină MT 1 stimulează producția de cAMP în celulele SH-SY5Y ale liniei celulare de neuroblastom uman printr-o cale de transducție a semnalului calciu-calmodulină. J. Neuroendocrinol. 2005 , 17 , 170–178. [ Google Scholar ] [ CrossRef ]
Peschke, E.; Muhlbauer, E.; Musshoff, U.; Csernus, VJ; Chankiewitz, E.; Peschke, D. Influența mediată de receptorul (MT1 ) a melatoninei asupra concentrației de cAMP și secreției de insulină a celulelor de insulinom de șobolan INS-1. J. Pineal Res. 2002 , 33 , 63–71. [ Google Scholar ] [ CrossRef ]
Witt-Enderby, PA; MacKenzie, RS; McKeon, RM; Carroll, EA; Bordt, SL; Melan, MA Inducerea melatoninei a structurilor filamentoase în celulele non-neuronale care este dependentă de expresia receptorului uman de melatonină MT 1 . Cell Motil. Citoschelet. 2000 , 46 , 28–42. [ Google Scholar ] [ CrossRef ]
Rivera-Bermudez, MA; Gerdin, MJ; Earnest, DJ; Dubocovich, ML Reglarea ritmicității bazale în activitatea protein kinazei C de către melatonină în celulele nucleului suprachiasmatic de șobolan imortalizate. Neurosci. Lett. 2003 , 346 , 37–40. [ Google Scholar ] [ CrossRef ]
Benitez-King, G.; Rios, A.; Martinez, A.; Anton-Tay, F. Inhibarea in vitro a activității kinazei II dependente de Ca2 + /calmodulină de către melatonină. Biochim. Biophys. Acta 1996 , 1290 , 191–196. [ Google Scholar ] [ CrossRef ]
Chan, AS; Lai, FP; Lo, RK; Voyno-Yasenetskaya, TA; Stanbridge, EJ; Receptorii Wong, YH Melatonina MT 1 și MT 2 stimulează kinaza N -terminală c-Jun prin proteinele G sensibile și insensibile la toxina pertussis. Semnal celular. 2002 , 14 , 249–257. [ Google Scholar ] [ CrossRef ]
Arends, YM; Duyckaerts, C.; Rozemuller, JM; Eikelenboom, P.; Hauw, JJ Microglia, amiloid și demență în boala alzheimer. Un studiu corelativ. Neurobiol. Îmbătrânire 2000 , 21 , 39–47. [ Google Scholar ] [ PubMed ]
Combadiere, C.; Feumi, C.; Raoul, W.; Keller, N.; Rodero, M.; Pezard, A.; Lavalette, S.; Houssier, M.; Jonet, L.; Picard, E.; et al. Acumularea de celule microglia subretiniene dependentă de CX3CR1 este asociată cu caracteristicile cardinale ale degenerescenței maculare legate de vârstă. J. Clin. Investig. 2007 , 117 , 2920–2928. [ Google Scholar ] [ CrossRef ][ Versiunea verde ]
Streit, WJ; Mrak, RE; Griffin, WS Microglia și neuroinflamație: o perspectivă patologică. J. Neuroinflamm. 2004 , 1 . [ Google Scholar ] [ CrossRef ][ Versiunea verde ]
Shen, Y.; Zhang, G.; Liu, L.; Xu, S. Efectele supresive ale melatoninei asupra activării gliale induse de amiloid-β în hipocampul șobolanului. Arc. Med. Res. 2007 , 38 , 284–290. [ Google Scholar ] [ CrossRef ]
Stuchbury, G.; Munch, G. Inflamația asociată cu Alzheimer, potențialele ținte de droguri și terapiile viitoare. J. Transm. Neural. 2005 , 112 , 429–453. [ Google Scholar ] [ CrossRef ]
Park, SY; Jin, ML; Kim, YH; Kim, Y.; Lee, SJ Efecte antiinflamatorii ale turmeronului aromatic prin blocarea căilor de semnalizare MAPK NF-κB, JNK și p38 în microglia stimulată de beta-amiloid. Int. Imunofarmacol. 2012 , 14 , 13–20. [ Google Scholar ] [ CrossRef ]
Samuel, W.; Masliah, E.; Hill, LR; Unturi, N.; Terry, R. Hipocampal conectivitate și demența Alzheimer: Efectele pierderii sinapselor și frecvența încurcăturii într-un model cu două componente. Neurologie 1994 , 44 , 2081–2088. [ Google Scholar ] [ CrossRef ]
Terry, AV, Jr.; Buccafusco, JJ Ipoteza colinergică a vârstei și a deficitelor cognitive legate de boala Alzheimer: provocări recente și implicațiile lor pentru dezvoltarea de noi medicamente. J. Pharmacol. Exp. Acolo. 2003 , 306 , 821–827. [ Google Scholar ] [ CrossRef ]
Rinne, JO; Laine, M.; Hiltunen, J.; Erkinjuntti, T. Luarea deciziilor semantice în AD probabile timpurii: Un studiu de activare PET. Brain Res. Cogn. Brain Res. 2003 , 18 , 89–96. [ Google Scholar ] [ CrossRef ]
Spencer, JP; Middleton, LJ; Davies, CH Investigarea eficacității inhibitorului de acetilcolinesterază, a donepezilului și a noilor agenți procognitivi pentru a induce oscilații γ în feliile de hipocamp de șobolan. Neurofarmacologie 2010 , 59 , 437–443. [ Google Scholar ] [ CrossRef ]
Guermonprez, L.; Ducrocq, C.; Gaudry-Talarmain, YM Inhibarea sintezei acetilcolinei și nitrarea tirozinei induse de peroxinitrit sunt prevenite în mod diferențial de antioxidanți. Mol. Pharmacol. 2001 , 60 , 838–846. [ Google Scholar ] [ PubMed ]
Feng, Z.; Cheng, Y.; Zhang, JT Efectele pe termen lung ale melatoninei sau 17 β-estradiol asupra îmbunătățirii performanței memoriei spațiale la șobolani adulți ovariectomizați, cu deficiențe cognitive. J. Pineal Res. 2004 , 37 , 198–206. [ Google Scholar ] [ CrossRef ]
Tang, F.; Nag, S.; Shiu, SY; Pang, SF Efectele melatoninei și extractului de Ginkgo biloba asupra pierderii memoriei și activităților colin acetiltransferazei în creierul șobolanilor infuzați intracerebroventricular cu β-amiloid 1-40. Life Sci. 2002 , 71 , 2625–2631. [ Google Scholar ] [ CrossRef ]
Agrawal, R.; Tyagi, E.; Shukla, R.; Nath, C. Un studiu al receptorilor de insulină din creier, activitatea AChE și stresul oxidativ în modelul de șobolan al demenței induse de ICV STZ. Neurofarmacologie 2009 , 56 , 779–787. [ Google Scholar ] [ CrossRef ]
Fernandez-Bachiller, MI; Perez, C.; Campillo, NE; Paez, JA; Gonzalez-Munoz, GC; Usan, P.; Garcia-Palomero, E.; Lopez, MG; Villarroya, M.; Garcia, AG; et al. Hibrizii tacrină-melatonină ca agenți multifuncționali pentru boala Alzheimer, cu proprietăți colinergice, antioxidante și neuroprotectoare. ChemMedChem 2009 , 4 , 828–841. [ Google Scholar ] [ CrossRef ]
Spuch, C.; Antequera, D.; Isabel Fernandez-Bachiller, M.; Isabel Rodriguez-Franco, M.; Carro, E. Un nou hibrid tacrina-melatonina reduce sarcina de amiloid si deficitele comportamentale intr-un model de soarece al bolii Alzheimer. Neurotox. Res. 2010 , 17 , 421–431. [ Google Scholar ] [ CrossRef ]
O’Neill, C.; Kiely, AP; Coakley, MF; Manning, S.; Long-Smith, CM Insulină și semnalizare IGF-1: longevitate, homeostazia proteinelor și boala Alzheimer. Biochim. Soc. Trans. 2012 , 40 , 721–727. [ Google Scholar ] [ CrossRef ]
Hildreth, KL; van Pelt, RE; Schwartz, RS Obezitatea, rezistența la insulină și boala Alzheimer. Obezitate 2012 , 20 , 1549–1557. [ Google Scholar ] [ CrossRef ]
De la Monte, SM Rezistența creierului la insulină și deficiența ca ținte terapeutice în boala Alzheimer. Curr. Alzheimer Res. 2012 , 9 , 35–66. [ Google Scholar ] [ CrossRef ]
Thambisetty, M.; Jeffrey, ME; Yang, A.; Dolan, H.; Marano, C.; Zonderman, AB; Troncoso, JC; Zhou, Y.; Wong, DF; Ferrucci, L.; et al. Intoleranța la glucoză, rezistența la insulină și caracteristicile patologice ale bolii Alzheimer în studiul longitudinal Baltimore asupra îmbătrânirii. JAMA Neurol. 2013 , 70 , 1167–1172. [ Google Scholar ] [ CrossRef ]
Cardinali, DP; Cano, P.; Jimenez-Ortega, V.; Esquifino, AI Melatonina și sindromul metabolic: implicații fiziopatologice și terapeutice. Neuroendocrinologie 2011 , 93 , 133–142. [ Google Scholar ] [ CrossRef ]
Cardinali, DP; Bernasconi, PA; Reynoso, R.; Toso, CF; Scacchi, P. Melatonina poate reduce sindromul metabolic: Studii asupra sindromului metabolic inițial și complet stabilit indus de fructoză la șobolani. Int. J. Mol. Sci. 2013 , 14 , 2502–2514. [ Google Scholar ] [ CrossRef ]
Magri, F.; Locatelli, M.; Balza, G.; Molla, G.; Cuzzoni, G.; Fioravanti, M.; Solerte, SB; Ferrari, E. Modificări ale ritmurilor circadiene endocrine ca markeri ai îmbătrânirii creierului fiziologic și patologic. Chronobiol. Int. 1997 , 14 , 385–396. [ Google Scholar ] [ CrossRef ]
Zhou, JN; Liu, RY; Kamphorst, W.; Hofman, MA; Swaab, DF Modificările neuropatologice precoce ale bolii Alzheimer la persoanele în vârstă sunt însoțite de scăderea nivelului de melatonină în lichidul cefalorahidian. J. Pineal Res. 2003 , 35 , 125–130. [ Google Scholar ] [ CrossRef ]
Skene, DJ; Vivien-Roels, B.; Sparks, DL; Hunsaker, JC; Pevet, P.; Ravid, D.; Swaab, DF Variația zilnică a concentrației de melatonină și 5-metoxitriptofol în glanda pineală umană: Efectul vârstei și al bolii Alzheimer. Brain Res. 1990 , 528 , 170–174. [ Google Scholar ] [ CrossRef ]
Ohashi, Y.; Okamoto, N.; Uchida, K.; Iyo, M.; Mori, N.; Morita, Y. Ritmul zilnic al nivelurilor serice de melatonina si efectul expunerii la lumina la pacientii cu dementa de tip Alzheimer. Biol. Psihiatrie 1999 , 45 , 1646–1652. [ Google Scholar ] [ CrossRef ]
Liu, RY; Zhou, JN; van Heerikhuize, J.; Hofman, MA; Swaab, DF Scăderea nivelului de melatonina în lichidul cefalorahidian postmortem în raport cu îmbătrânirea, boala Alzheimer și genotipul apolipoproteinei E-epsilon4/4. J. Clin. Endocrinol. Metab. 1999 , 84 , 323–327. [ Google Scholar ] [ PubMed ]
Mishima, K.; Tozawa, T.; Satoh, K.; Matsumoto, Y.; Hishikawa, Y.; Okawa, M. Tulburări ale ritmului de secreție a melatoninei la pacienții cu demență senilă de tip Alzheimer cu tulburări de somn-vegie. Biol. Psihiatrie 1999 , 45 , 417–421. [ Google Scholar ] [ CrossRef ]
Skene, DJ; Swaab, DF Ritmicitatea melatoninei: Efectul vârstei și al bolii Alzheimer. Exp. Gerontol. 2003 , 38 , 199–206. [ Google Scholar ] [ CrossRef ]
Wu, YH; Feenstra, MG; Zhou, JN; Liu, RY; Torano, JS; van Kan, HJ; Fischer, DF; Ravid, R.; Swaab, DF Modificari moleculare care stau la baza nivelurilor reduse de melatonina pineala in boala Alzheimer: modificari in stadiile preclinice si clinice. J. Clin. Endocrinol. Metab. 2003 , 88 , 5898–5906. [ Google Scholar ] [ CrossRef ]
Savaskan, E.; Ayoub, MA; Ravid, R.; Angeloni, D.; Fraschini, F.; Meier, F.; Eckert, A.; Muller-Spahn, F.; Jockers, R. Expresia redusă a receptorului melatoninei MT 2 din hipocamp în boala Alzheimer. J. Pineal Res. 2005 , 38 , 10–16. [ Google Scholar ] [ CrossRef ]
Savaskan, E.; Olivieri, G.; Meier, F.; Brydon, L.; Jockers, R.; Ravid, R.; Wirz-Justice, A.; Muller-Spahn, F. Imunoreactivitate crescută a receptorului de melatonină 1a în hipocampul pacienților cu boala Alzheimer. J. Pineal Res. 2002 , 32 , 59–62. [ Google Scholar ] [ CrossRef ]
Weldemichael, DA; Grossberg, GT Tulburări de ritm circadian la pacienții cu boala Alzheimer: o revizuire. Int. J. Alzheimer Dis. 2010 , 2010 . [ Google Scholar ] [ CrossRef ]
Klaffke, S.; Staedt, J. Sundowning și tulburări de ritm circadian în demență. Acta Neurol. Belg. 2006 , 106 , 168–175. [ Google Scholar ] [ PubMed ]
Cardinali, DP; Furio, AM; Brusco, LI Aspecte clinice ale intervenției melatoninei în progresia bolii Alzheimer. Curr. Neurofarmacol. 2010 , 8 , 218–227. [ Google Scholar ] [ CrossRef ]
De Jonghe, A.; Korevaar, JC; van Munster, BC; de Rooij, SE Eficacitatea tratamentului cu melatonină asupra tulburărilor de ritm circadian în demență. Există implicații pentru delir? O revizuire sistematică. Int. J. Geriatr. Psihiatrie 2010 , 25 , 1201–1208. [ Google Scholar ] [ CrossRef ]
Pappolla, MA; Chyan, YJ; Poeggeler, B.; Frangione, B.; Wilson, G.; Ghiso, J.; Reiter, RJ O evaluare a proprietăților antioxidante și antiamiloidogene ale melatoninei: Implicații pentru boala Alzheimer. J. Transm. Neural. 2000 , 107 , 203–231. [ Google Scholar ] [ CrossRef ]
Wu, YH; Swaab, DF Glanda pineală umană și melatonina în îmbătrânire și boala Alzheimer. J. Pineal Res. 2005 , 38 , 145–152. [ Google Scholar ] [ CrossRef ]
Wu, YH; Zhou, JN; van Heerikhuize, J.; Jockers, R.; Swaab, DF Scăderea expresiei receptorului de melatonină MT 1 în nucleul suprachiasmatic în îmbătrânire și boala Alzheimer. Neurobiol. Îmbătrânire 2007 , 28 , 1239–1247. [ Google Scholar ] [ CrossRef ]
Fainstein, I.; Bonetto, A.; Brusco, LI; Cardinali, DP Efectele melatoninei la pacienții vârstnici cu tulburări de somn. Un studiu pilot. Curr. Acolo. Res. 1997 , 58 , 990–1000. [ Google Scholar ] [ CrossRef ]
Brusco, LI; Marquez, M.; Cardinali, DP Tratamentul cu melatonina stabilizeaza simptomele cronobiologice si cognitive in boala Alzheimer. Neuro Endocrinol. Lett. 1998 , 19 , 111–115. [ Google Scholar ]
Brusco, LI; Marquez, M.; Cardinali, DP Gemeni monozigoți cu boala Alzheimer tratați cu melatonină: Raport de caz. J. Pineal Res. 1998 , 25 , 260–263. [ Google Scholar ] [ CrossRef ]
Cohen-Mansfield, J.; Garfinkel, D.; Lipson, S. Melatonina pentru tratamentul apusului soarelui la persoanele în vârstă cu demență – Un studiu preliminar. Arc. Gerontol. Geriatr. 2000 , 31 , 65–76. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Mishima, K.; Okawa, M.; Hozumi, S.; Hishikawa, Y. Administrarea suplimentară a luminii strălucitoare artificiale și a melatoninei ca tratament puternic pentru activitatea de repaus circadiană dezorganizată și sistemele autonome și neuroendocrine disfuncționale la persoanele vârstnice dementate instituționalizate. Chronobiol. Int. 2000 , 17 , 419–432. [ Google Scholar ] [ CrossRef ]
Serfaty, M.; Kennell-Webb, S.; Warner, J.; Blizard, R.; Raven, P. Studiu dublu orb, randomizat, controlat cu placebo, de melatonină în doză mică pentru tulburările de somn în demență. Int. J. Geriatr. Psihiatrie 2002 , 17 , 1120–1127. [ Google Scholar ] [ CrossRef ]
Cardinali, DP; Brusco, LI; Liberczuk, C.; Furio, AM Utilizarea melatoninei în boala Alzheimer. Neuro Endocrinol. Lett. 2002 , 23 , 20–23. [ Google Scholar ] [ PubMed ]
Singer, C.; Tractenberg, RE; Kaye, J.; Schafer, K.; Gamst, A.; Grundman, M.; Thomas, R.; Thal, LJ Un studiu multicentric, controlat cu placebo, al melatoninei pentru tulburările de somn în boala Alzheimer. Sleep 2003 , 26 , 893–901. [ Google Scholar ] [ PubMed ]
Asayama, K.; Yamadera, H.; Ito, T.; Suzuki, H.; Kudo, Y.; Endo, S. Studiu dublu-orb al efectelor melatoninei asupra ritmului somn-veghe, funcțiilor cognitive și non-cognitive în demența de tip Alzheimer. J. Nippon Med. Sch. 2003 , 70 , 334–341. [ Google Scholar ] [ CrossRef ]
Mahlberg, R.; Kunz, D.; Sutej, I.; Kuhl, KP; Hellweg, R. Tratamentul cu melatonina a tulburarilor de ritm zi-noapte si apusul soarelui in boala Alzheimer: un studiu pilot deschis folosind actigrafie. J. Clin. Psihofarmacol. 2004 , 24 , 456–459. [ Google Scholar ] [ PubMed ]
Mahlberg, R.; Walther, S. Actigrafie la pacientii agitati cu dementa. Monitorizarea rezultatelor tratamentului. Z. Gerontol. Geriatr. 2007 , 40 , 178–184. [ Google Scholar ] [ CrossRef ]
Anderson, KN; Jamieson, S.; Graham, AJ; Shneerson, JM Tulburare de comportament în somn REM tratată cu melatonină la un pacient cu boala Alzheimer. Clin. Neurol. Neurochirurgie. 2008 , 110 , 492–495. [ Google Scholar ] [ CrossRef ]
Dowling, GA; Burr, RL; van Someren, EJ; Hubbard, EM; Luxenberg, JS; Mastick, J.; Cooper, BA Melatonina și tratamentul cu lumină puternică pentru întreruperea activității de odihnă la pacienții instituționalizați cu boala Alzheimer. J. Am. Geriatr. Soc. 2008 , 56 , 239–246. [ Google Scholar ] [ CrossRef ]
Gehrman, PR; Connor, DJ; Martin, JL; Shochat, T.; Corey-Bloom, J.; Ancoli-Israel, S. Melatonin nu reușește să îmbunătățească somnul sau agitația într-un studiu dublu-orb randomizat controlat cu placebo de pacienți instituționalizați cu boala Alzheimer. A.m. J. Geriatr. Psihiatrie 2009 , 17 , 166–169. [ Google Scholar ] [ CrossRef ]
Farias, ST; Mungas, D.; Reed, BR; Harvey, D.; DeCarli, C. Progresia deficienței cognitive ușoare la demență în cohortele clinice vs comunitare. Arc. Neurol. 2009 , 66 , 1151–1157. [ Google Scholar ] [ PubMed ]
Davies, L.; Wolska, B.; Hilbich, C.; Multhaup, G.; Martins, R.; Simms, G.; Beyreuther, K.; Masters, depunerea de proteină amiloid CL A 4 și diagnosticul bolii Alzheimer: Prevalența în creierul îmbătrânit determinată prin imunocitochimie în comparație cu tehnicile neuropatologice convenționale. Neurologie 1988 , 38 , 1688–1693. [ Google Scholar ] [ CrossRef ]
Preț, JL; Morris, JC Încurcături și plăci în îmbătrânirea nedementabilă și boala Alzheimer „preclinic”. Ann. Neurol. 1999 , 45 , 358–368. [ Google Scholar ] [ CrossRef ]
Braak, H.; Braak, E. Stadializarea modificărilor neurofibrilare legate de boala Alzheimer. Neurobiol. Aging 1995 , 16 , 271–278. [ Google Scholar ] [ CrossRef ]
Braak, H.; Braak, E. Evoluția modificărilor neuronale în cursul bolii Alzheimer. J. Transm. Neural. Suppl. 1998 , 53 , 127–140. [ Google Scholar ] [ CrossRef ]
Furio, AM; Brusco, LI; Cardinali, DP Valoarea terapeutică posibilă a melatoninei în deficiența cognitivă ușoară. Un studiu retrospectiv. J. Pineal Res. 2007 , 43 , 404–409. [ Google Scholar ] [ CrossRef ]
Cardinali, DP; Vigo, DE; Olivar, N.; Vidal, MF; Furio, AM; Brusco, LI Aplicarea terapeutică a melatoninei în deficiența cognitivă ușoară. A.m. J. Neurodegener Dis. 2012 , 1 , 280–291. [ Google Scholar ] [ PubMed ]
Wu, YH; Swaab, DF Tulburări și strategii pentru reactivarea sistemului de ritm circadian în îmbătrânire și boala Alzheimer. Sleep Med. 2007 , 8 , 623–636. [ Google Scholar ] [ CrossRef ]
Pandi-Perumal, SR; BaHammam, AS; Brown, GM; Spence, DW; Bharti, VK; Kaur, C.; Hardeland, R.; Cardinali, DP Apărarea antioxidativă a melatoninei: implicații terapeutice pentru procesele de îmbătrânire și neurodegenerative. Neurotox. Res. 2013 , 23 , 267–300. [ Google Scholar ] [ CrossRef ]
Rosales-Corral, SA; Acuña-Castroviejo, D.; Coto-Montes, A.; Boga, JA; Manchester, LC; Fuentes-Broto, L.; Korkmaz, A.; Ma, S.; Tan, DX; Reiter, RJ Boala Alzheimer: mecanisme patologice și rolul benefic al melatoninei. J. Pineal Res. 2012 , 52 , 167–202. [ Google Scholar ] [ CrossRef ]
Tan, DX; Manchester, LC; Sanchez-Barcelo, E.; Mediavilla, MD; Reiter, RJ Semnificația nivelurilor ridicate de melatonină endogene în lichidul cefalorahidian al mamiferelor și în sistemul nervos central. Curr. Neurofarmacol. 2010 , 8 , 162–167. [ Google Scholar ] [ CrossRef ]
Legros, C.; Chesneau, D.; Boutin, JA; Barc, C.; Malpaux, B. Melatonina din lichidul cefalorahidian, dar nu din sânge, ajunge la țesuturile cerebrale de oaie în condiții fiziologice. J. Neuroendocrinol. 2014 . [ Google Scholar ] [ CrossRef ]
Monti, JM; Alvarino, F.; Cardinali, DP; Savio, I.; Pintos, A. Studiu polisomnografic al efectului melatoninei asupra somnului la pacienții vârstnici cu insomnie primară cronică. Arc. Gerontol. Geriatr. 1999 , 28 , 85–98. [ Google Scholar ] [ CrossRef ]
Cardinali, DP; Srinivasan, V.; Brzezinski, A.; Brown, GM Melatonin și analogii săi în insomnie și depresie. J. Pineal Res. 2012 , 52 , 365–375. [ Google Scholar ] [ CrossRef ]
Rajaratnam, SM; Polymeropoulos, MH; Fisher, DM; Roth, T.; Scott, C.; Birznieks, G.; Klerman, EB Tasimelteon agonist al melatoninei (VEC-162) pentru insomnia tranzitorie după schimbarea timpului de somn: două studii multicentre controlate randomizate. Lancet 2009 , 373 , 482–491. [ Google Scholar ] [ CrossRef ]
Mulchahey, JJ; Goldwater, DR; Zemlan, FP Un singur studiu orb, controlat cu placebo, pe grupuri de creștere a dozei privind siguranța, tolerabilitatea, farmacocinetica și farmacodinamia analogului melatoninei β-metil-6-cloromelatoninei. Life Sci. 2004 , 75 , 1843–1856. [ Google Scholar ] [ CrossRef ]
McKenna, JT; Christie, MA; Jeffrey, BA; McCoy, JG; Lee, E.; Connolly, NP; Ward, CP; Strecker, RE Tratamentul cronic cu ramelteon într-un model de șoarece al bolii Alzheimer. Arc. Ital. Biol. 2012 , 150 , 5–14. [ Google Scholar ] [ PubMed ]
Ursing, C.; Hartter, S.; von Bahr, C.; Tybring, G.; Bertilsson, L.; Rojdmark, S. Metabolismul hepatic al melatoninei afectează nivelul melatoninei serice endogen la om? J. Endocrinol. Investig. 2002 , 25 , 459–462. [ Google Scholar ] [ CrossRef ]
Hartter, S.; Grozinger, M.; Weigmann, H.; Roschke, J.; Hiemke, C. Biodisponibilitate crescută a melatoninei orale după administrarea concomitentă a fluvoxaminei. Clin. Pharmacol. Acolo. 2000 , 67 , 1–6. [ Google Scholar ] [ CrossRef ]
Hartter, S.; Nordmark, A.; Trandafir, DM; Bertilsson, L.; Tybring, G.; Laine, K. Efectele consumului de cofeină asupra farmacocineticii melatoninei, un medicament probă pentru activitatea CYP1A2. Br. J. Clin. Pharmacol. 2003 , 56 , 679–682. [ Google Scholar ] [ CrossRef ]
Souetre, E.; Salvati, E.; Belugou, JL; de Galeani, B.; Krebs, B.; Ortonne, JP; Darcourt, G. 5-Methoxypsoralen crește nivelurile plasmatice de melatonina la om. J. Investig. Dermatol. 1987 , 89 , 152–155. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Garde, E.; Micic, S.; Knudsen, K.; Angelo, HR; Wulf, HC 8-Methoxypsoralen crește nivelurile plasmatice de melatonina în timpul zilei la oameni prin inhibarea metabolismului. Fotochimie. Fotobiol. 1994 , 60 , 475–480. [ Google Scholar ] [ CrossRef ]
Mauviard, F.; Raynaud, F.; Geoffriau, M.; Claustrat, B.; Pevet, P. 5-Methoxypsoralen inhibă 6-hidroxilarea melatoninei la șobolan. Biol. Semnale 1995 , 4 , 32–41. [ Google Scholar ] [ CrossRef ]
Weishaupt, JH; Bartels, C.; Polking, E.; Dietrich, J.; Rohde, G.; Poeggeler, B.; Mertens, N.; Sperling, S.; Bohn, M.; Huther, G.; et al. Reducerea daunelor oxidative în SLA prin tratamentul cu doze mari de melatonină enterală. J. Pineal Res. 2006 , 41 , 313–323. [ Google Scholar ] [ CrossRef ]
Chahbouni, M.; Escames, G.; Venegas, C.; Sevilla, B.; Garcia, JA; Lopez, LC; Munoz-Hoyos, A.; Molina-Carballo, A.; Acuña-Castroviejo, D. Tratamentul cu melatonina normalizează citokinele proinflamatorii plasmatice și stresul nitrozativ/oxidativ la pacienții care suferă de distrofie musculară Duchenne. J. Pineal Res. 2010 , 48 , 282–289. [ Google Scholar ] [ CrossRef ]
Voordouw, BC; Euser, R.; Verdonk, RE; Alberda, BT; de Jong, FH; Drogendijk, AC; Fauser, BC; Cohen, M. Melatonina și combinațiile melatonină-progestin modifică funcția hipofizo-ovariană la femei și pot inhiba ovulația. J. Clin. Endocrinol. Metab. 1992 , 74 , 108–117. [ Google Scholar ] [ PubMed ]
Nickkholgh, A.; Schneider, H.; Sobirey, M.; Venetz, WP; Hinz, U.; Pelzle, H.; Gotthardt, DN; Cekauskas, A.; Manikas, M.; Mikalauskas, S.; et al. Utilizarea melatoninei în doze mari în rezecția hepatică este sigură: prima experiență clinică. J. Pineal Res. 2011 , 50 , 381–388. [ Google Scholar ] [ CrossRef ]
Cardinali, DP; Pandi-Perumal, SR; Niles, LP Melatonina și receptorii săi: Funcția biologică în reglementarea circadiană a somnului-veghe. În Neurochimia somnului și a stării de veghe ; Monti, JM, Pandi-Perumal, SR, Sinton, CM, Eds.; Cambridge University Press: Cambridge, Marea Britanie, 2008; p. 283–314. [ Google Scholar ]
Dagan, Y.; Zisapel, N.; Nof, D.; Laudon, M.; Atsmon, J. Inversarea rapidă a toleranței la hipnoticele benzodiazepine prin tratament cu melatonină orală: un raport de caz. EURO. Neuropsihofarmacol. 1997 , 7 , 157–160. [ Google Scholar ] [ CrossRef ]
Garfinkel, D.; Zisapel, N.; Wainstein, J.; Laudon, M. Facilitarea întreruperii benzodiazepinei de către melatonină: O nouă abordare clinică. Arc. Intern. Med. 1999 , 159 , 2456–2460. [ Google Scholar ] [ CrossRef ]
Siegrist, C.; Benedetti, C.; Orlando, A.; Beltran, JM; Tuchscherr, L.; Noseda, CM; Brusco, LI; Cardinali, DP Lipsa modificărilor serice ale prolactinei, FSH, TSH și estradiolului după tratamentul cu melatonină în doze care îmbunătățesc somnul și reduc consumul de benzodiazepine la pacienții cu tulburări de somn, de vârstă mijlocie și vârstnici. J. Pineal Res. 2001 , 30 , 34–42. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Kunz, D.; Bineau, S.; Maman, K.; Milea, D.; Toumi, M. Întreruperea benzodiazepinei cu melatonină cu eliberare prelungită: Sugestii dintr-o bază de date de prescripție longitudinală germană. Opinia expertului. Pharmacother. 2012 , 13 , 9–16. [ Google Scholar ] [ CrossRef ]
Clay, E.; Falissard, B.; Moore, N.; Toumi, M. Contribuția campaniilor de melatonină și anti-benzodiazepină cu eliberare prelungită la reducerea consumului de benzodiazepine și medicamente Z în nouă țări europene. EURO. J. Clin. Pharmacol. 2013 , 69 , 1–10. [ Google Scholar ] [ PubMed ]
Leger, D.; Laudon, M.; Zisapel, N. Excreția nocturnă a 6-sulfatoximelatoninei în insomnie și relația sa cu răspunsul la terapia de substituție cu melatonină. A.m. J. Med. 2004 , 116 , 91–95. [ Google Scholar ] [ CrossRef ]
Zhdanova, IV; Wurtman, RJ; Regan, MM; Taylor, JA; Shi, JP; Leclair, OU Tratament cu melatonina pentru insomnia legata de varsta. J. Clin. Endocrinol. Metab. 2001 , 86 , 4727–4730. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Wilson, SJ; Nutt, DJ; Alford, C.; Argyropoulos, SV; Baldwin, DS; Bateson, AN; Britton, TC; Crowe, C.; Dijk, DJ; Espie, CA; et al. Declarație de consens al Asociației Britanice pentru Psihofarmacologie privind tratamentul bazat pe dovezi al insomniei, parasomniilor și tulburărilor de ritm circadian. J. Psihofarmacol. 2010 , 24 , 1577–1601. [ Google Scholar ] [ CrossRef ]
Cardinali, DP; Furio, AM; Brusco, LI Utilizarea cronobioticelor în resincronizarea ciclului somn/veghe. Aplicație terapeutică în fazele incipiente ale bolii Alzheimer. Brevetul recent. Endocr. Metab. Drug Imun Discov. 2011 , 5 , 80–90. [ Google Scholar ] [ CrossRef ]
Jean-Louis, G.; von Gizycki, H.; Zizi, F. Efectele melatoninei asupra somnului, dispoziției și cogniției la vârstnici cu deficiențe cognitive ușoare. J. Pineal Res. 1998 , 25 , 177–183. [ Google Scholar ] [ CrossRef ]
Peck, JS; LeGoff, DB; Ahmed, I.; Goebert, D. Efectele cognitive ale administrării melatoninei exogene la persoanele în vârstă: un studiu pilot. A.m. J. Geriatr. Psihiatrie 2004 , 12 , 432–436. [ Google Scholar ] [ PubMed ]
Wade, AG; Ford, I.; Crawford, G.; McMahon, AD; Nir, T.; Laudon, M.; Zisapel, N. Eficacitatea melatoninei cu eliberare prelungită la pacienții cu insomnie cu vârsta cuprinsă între 55 și 80 de ani: calitatea somnului și rezultatele de vigilență în ziua următoare. Curr. Med. Res. Opinează. 2007 , 23 , 2597–2605. [ Google Scholar ] [ CrossRef ]
Riemersma-van der Lek, RF; Swaab, DF; Twisk, J.; Hol, EM; Hoogendijk, WJ; van Someren, EJ Efectul luminii strălucitoare și al melatoninei asupra funcției cognitive și noncognitive la rezidenții vârstnici ai unităților de îngrijire de grup: un studiu controlat randomizat. JAMA 2008 , 299 , 2642–2655. [ Google Scholar ] [ CrossRef ]
Garzon, C.; Guerrero, JM; Aramburu, O.; Guzman, T. Efectul administrării melatoninei asupra somnului, tulburărilor de comportament și întreruperii medicamentelor hipnotice la vârstnici: un studiu randomizat, dublu-orb, controlat cu placebo. Aging Clin. Exp. Res. 2009 , 21 , 38–42. [ Google Scholar ] [ CrossRef ]
Cazzola, R.; Rondanelli, M.; Faliva, M.; Cestaro, B. Efectele suplimentării cu DHA-fosfolipide, melatonină și triptofan asupra proprietăților fizico-chimice ale membranei eritrocitelor la pacienții vârstnici care suferă de deficiență cognitivă ușoară. Exp. Gerontol. 2012 , 47 , 974–978. [ Google Scholar ] [ CrossRef ]
Rondanelli, M.; Opizzi, A.; Faliva, M.; Mozzoni, M.; Antoniello, N.; Cazzola, R.; Savare, R.; Cerutti, R.; Grossi, E.; Cestaro, B. Efectele unei integrări dietetice cu o emulsie uleioasă de DHA-fosfolipide care conțin melatonină și triptofan la pacienții vârstnici care suferă de deficiență cognitivă ușoară. Nutr. Neurosci. 2012 , 15 , 46–54. [ Google Scholar ] [ CrossRef ] [ PubMed ]
















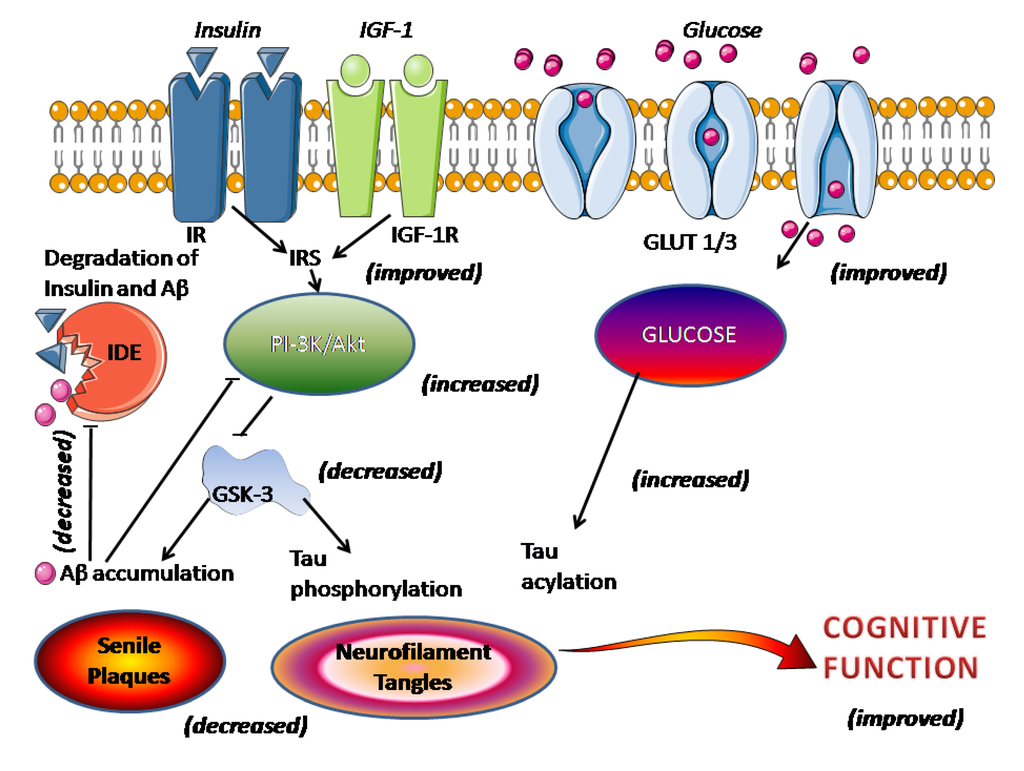
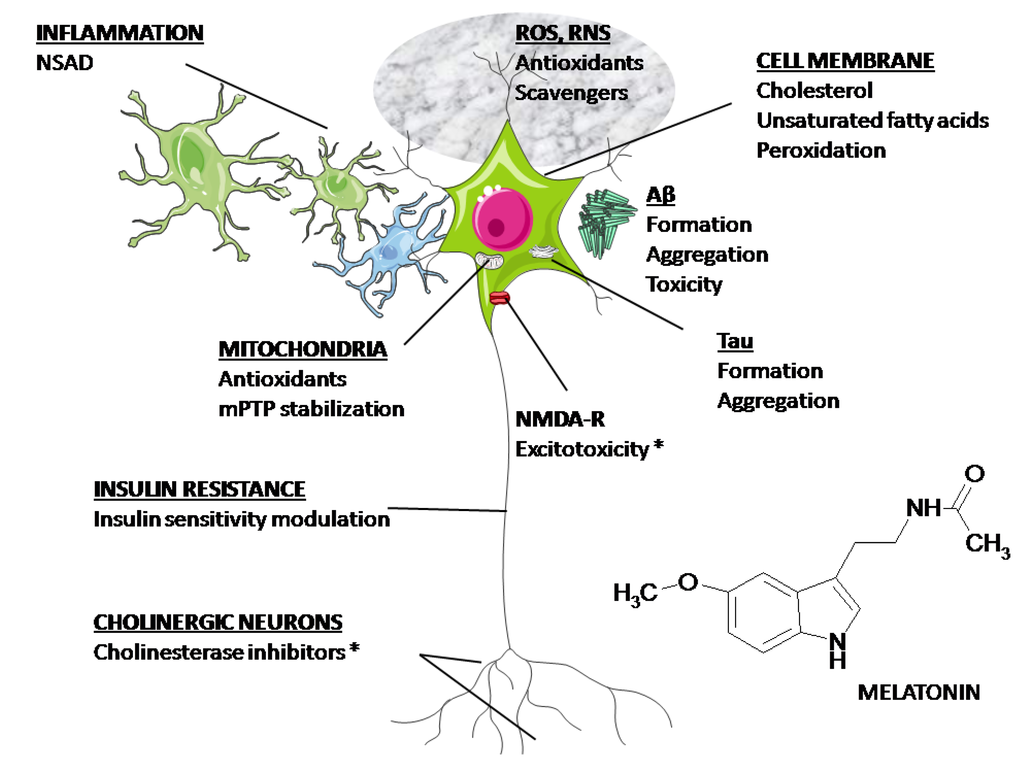


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
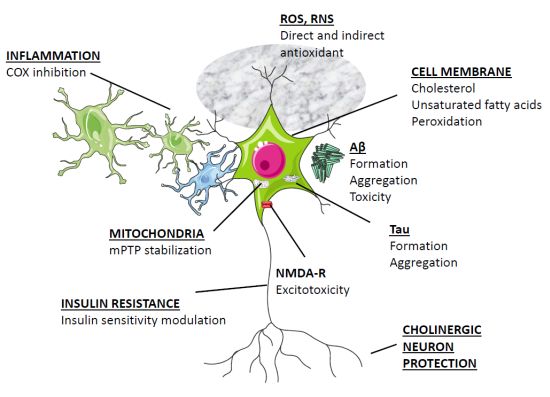{kind=link}
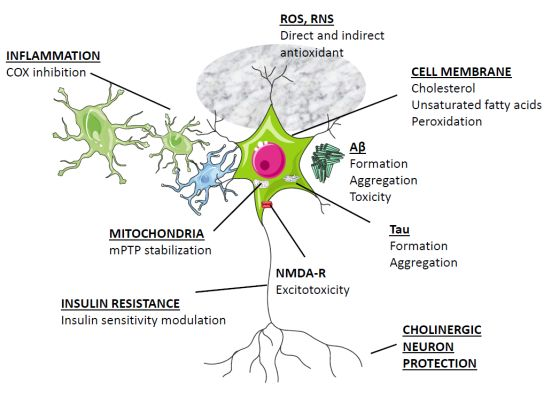{kind=link}
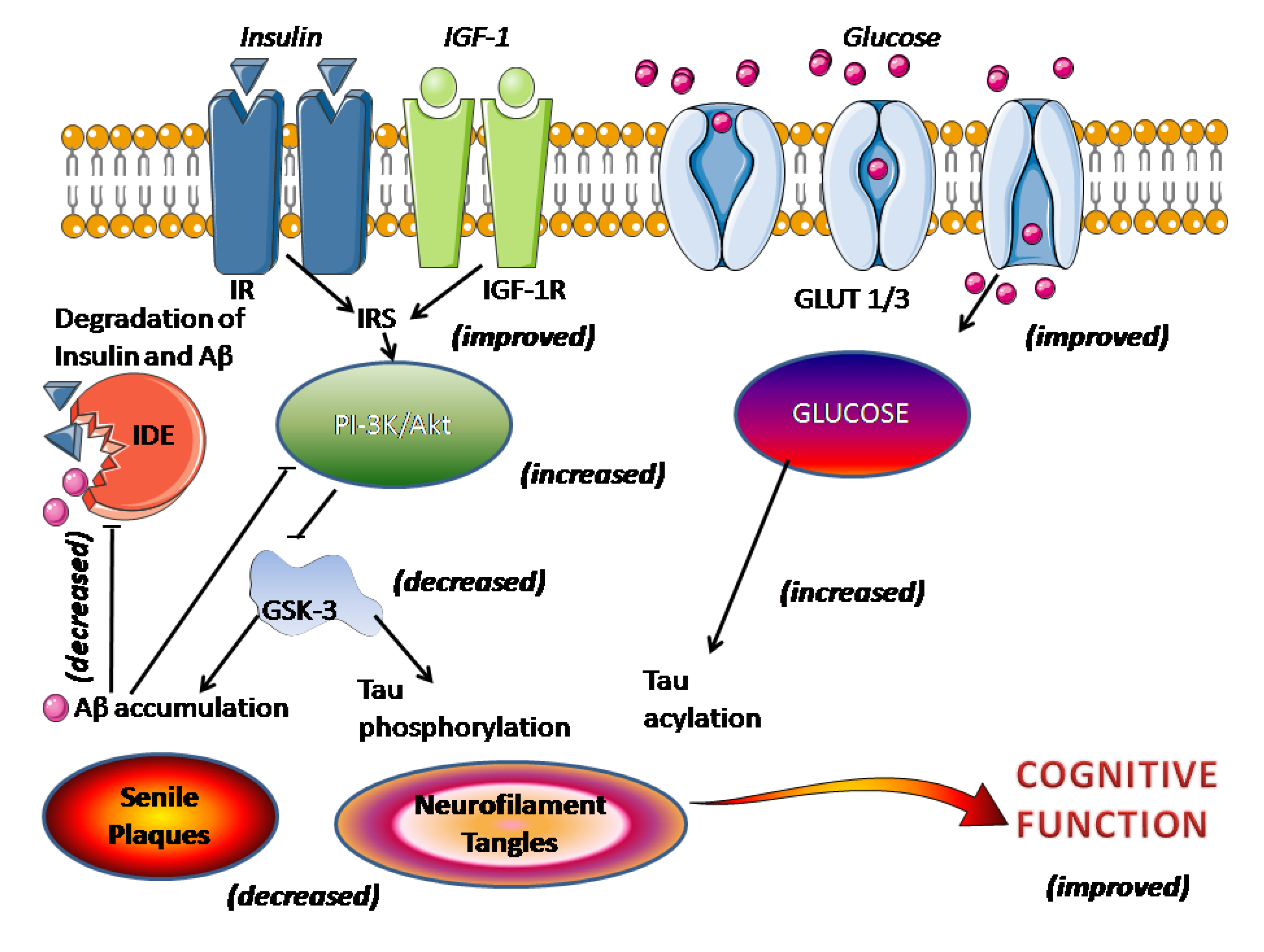{kind=link}
{kind=link}